
Abstract
Western blotting analysis of mouse nasal tissue using a specific anti-mouse secreted carbonic anhydrase (CA VI) antibody has shown that CA VI is present in this tissue. A single immunoreactive band of 42 kD was observed, as has been found previously for salivary tissues. RT-PCR analysis has shown that nasal mucosa expressed CA VI mRNA. By immunohistochemistry (IHC), CA VI was observed in acinar cells, in duct contents of the anterior gland of the nasal septum, and in the lateral nasal gland. The Bowman's gland, the posterior gland of the nasal septum, and the maxillary sinus gland were negative. Immunoreactivity was also observed in the mucus covering the respiratory and olfactory mucosa and in the lumen of the nasolacrimal duct. In contrast, an anti-rat CA II antibody (that crossreacts with the mouse enzyme) stained only known CA II-positive cells and an occasional olfactory receptor neuron. These results indicate that CA VI is produced by the nasal gland and is secreted over the nasal mucosa. By reversible hydration of CO2, CA VI is presumed to play a role in mucosal functions such as CO2 sensation and acid–base balance. It may also play a role in olfactory function as a growth factor in maturation of the olfactory epithelial cells.
C
A protein named gustin that is present in saliva and thought to be involved in the mechanism of taste has been described and has been shown to be identical to CA VI (Thatcher et al. 1998). Henkin et al. (1971,1975) described a syndrome characterized by hypogeusia (loss of taste) with or without dysgeusia (distortion of taste) and hyposmia (loss of smell) with or without dysosmia (distortion of smell) after an influenza-like illness. The patients’ saliva and nasal mucus characteristically showed decreased concentrations of CA (Doherty et al. 1997; Henkin et al. 1999a). After treatment with zinc, the enzyme concentration returned to normal and the taste and smell acuity were improved (Henkin et al. 1999b). These facts suggest that CA VI is implicated in oral and nasal sensory functions. In fact, not only systemic but also topical application of a CA inhibitor, such as acetazolamide, alters taste sensation and eliminates the pungency or prickly sensation of carbonated drinks (Hansson, as quoted in Graber and Kelleher 1988).
CA VI in tears is discharged through the nasolacrimal ducts into the nasal vestibule and may contribute to the nasal CA VI. Hansson's histochemistry has demonstrated CA activity in nasal glands of the guinea pig and bullfrog (Okamura et al. 1996; Coates et al. 1998). A molecular analysis of the mucosal scrapings has suggested that CA VI is produced in the human nose (Tarun et al. 2003). In the present study, using IHC in combination with biochemical and molecular techniques, we have localized CA VI in the mouse nasal gland. Therefore, CA VI is secreted by four exocrine glands: salivary, lacrimal, mammary, and nasal glands.
Materials and Methods
The study was reviewed and approved by the Intramural Animal Use and Care Committee of Osaka University Graduate School of Dentistry. All procedures were carried out at room temperature unless otherwise noted. Animals used were 6-week-old male mice of the ICR strain. After exsanguination under deep anesthesia with chloral hydrate (20 mg/100 g bw), whole rostrum including the nasal cavity was obtained. Soft tissues surrounding the rostrum, e.g., skin, oral mucosa, and salivary glands, were completely removed to expose the viscerocranium. For extraction of RNA, the rostrum was cut mid-sagittally. The nasal mucosa was carefully scraped from the lateral and septal walls of the nasal cavity with a clean sharp spatula. From some animals, submandibular gland, liver, and blood were obtained. The tissues were immediately frozen in liquid nitrogen and stored at –80C until used, and the blood was directly processed to prepare erythrocyte lysate according to the method described previously (Ogawa et al. 1992).
For IHC, anesthetized mice were perfused through the left ventricle with 4% paraformaldehyde and 0.05% glutaraldehyde in 0.1 M phosphate buffer (pH 7.2). The rostrum was removed, cleared of the surrounding soft tissues, and immersed in the same fixative overnight at 4C. The rostrum was then decalcified in 4.13% EDTA (pH 7.2) containing 0.2 M sucrose at 4C for a week, washed in 10 mM PBS, pH 7.2, containing 0.25 M sucrose, dehydrated, and embedded in paraffin. Coronal sections (6 μm thick) were cut and mounted on silane-coated glass slides.
Purification of Mouse CA and Production of Antibody to Mouse CA VI
Submandibular glands were homogenized on ice in 50 mM phosphate buffer (pH 7.4) containing 5 mM benzamidine and 1 mg/ml aprotinin. The homogenate was centrifuged (20,000 × g for 15 min at 4C). The supernatant was filtered through a 0.45-μm filter and loaded onto a sulfonamide-Sepharose affinity column. CA was isolated as described previously (Fernley et al. 1988). CA VI was purified from this salivary CA by anion exchange chromatography (Q Sepha-rose; Amersham Pharmacia Biotech, Uppsala, Sweden). The enzyme was eluted from the column with a linear salt gradient (0–0.5 M NaCl) in 50 mM Tris buffer (pH 8.0).
Rostrums were homogenized and the supernatant was prepared as described above. Nasal CA was isolated from the supernatant by the sulfonamide affinity chromatography, as was erythrocyte CA from erythrocyte lysate.
To raise a polyclonal antibody, 0.1 mg of purified CA VI was emulsified with an equal volume of Freund's complete adjuvant and injected SC into a male New Zealand rabbit. The rabbit was boosted with the same volume of the antigen in Freund's incomplete adjuvant according to the protocol by Fernley et al. (1988). Two weeks after the second booster, exsanguination was performed under anesthesia with pentobarbital sodium (50 mg/kg bw) and the antiserum was obtained. Immunoglobulin was prepared from the antiserum by protein G affinity chromatography (HiTrap Protein G HP; Amersham Pharmacia Biotech).
Western Analysis
Western analysis was performed as described previously (Ogawa et al. 2002). CA samples were run on a 10.5% SDS-polyacrylamide gel and electroblotted onto Immobilon (Millipore; Bedford, MA). The blot was blocked with 10% skimmed milk powder in 20 mM Tris-buffered saline (pH 7.5; TBS) and reacted with anti-CA VI antibody (1:5000 in 3% skimmed milk powder in TBS) or anti-rat erythrocyte CA antibody (1:10000; Ogawa et al. 1992). The blot was then reacted with peroxidase-conjugated goat anti-rabbit IgG (1:2500; Bio-Rad Laboratories; Richmond, CA) and the complex was visualized by treatment with 3,3'-diaminobenzidine tetrahydrochloride-H2O2 solution (DAB-H2O2).
RT-PCR Analysis
Total RNA was isolated separately from nasal mucosa, salivary gland, and liver, and RT and PCR were performed as described previously (Ogawa et al. 2002). For PCR, 35 cycles of denaturation (94C, 30 sec), annealing (58C, 30 sec), and extension (72C, 40 sec) were performed in a DNA thermal cycler (Perkin-Elmer; Norwalk, CT). The primer sequences for mouse CA VI were 5'-CTGTGTTAGCCGTCTTGTTTA-3’ and 5'-TTGAATGGTGTTGTTGTTGTG-3', which generate a 302-bp fragment (Sok et al. 1999). The primers for mouse glyceraldehyde-3-phosphate dehydrogenase (GAPDH), a constitutively expressed housekeeping gene, were 5'-AAGCAACATAGACGTTGTCGC-3’ and 5'-AATCAACACCTTCTTCGCACC-3', which generate a 286-bp fragment (Tso et al. 1985).
Immunohistochemistry
Deparaffinized sections were stained by an avidin-biotin-peroxidase technique (Ogawa et al. 1999). After treatment with 0.3% H2O2 and then with normal swine serum (1:100 in PBS containing 1% bovine serum albumin; PBS-BSA), the sections were reacted with anti-CA VI antibody (1:5000 in PBS-BSA) or anti-rat erythrocyte CA antibody (1:10000). Then they were sequentially reacted with biotinylated swine anti-rabbit IgG (1:500 in PBS-BSA containing 1% normal mouse serum; DAKO, Glostrup, Denmark) and streptavidin-biotinylated peroxidase reagent (1:100 in PBS; DAKO). After incubation with DAB-H2O2, the sections were briefly counterstained with Mayer's hematoxylin, dehydrated, and coverslipped with Permount.
Results
Western Analysis
CA VI was purified from the mouse salivary gland by sulfonamide affinity and ion-exchange chromatography. When treated with SDS with 2-mercaptoethanol and run on a 10.5% polyacrylamide gel in the presence of 0.1% SDS, the enzyme was seen as a single band with an apparent molecular weight of ≃42 kD, which was similar to that described previously (Fernley et al. 1989). The enzyme was used for the production of a rabbit antibody to mouse CA VI. The antibody recognized CA VI but not 30-kD erythrocyte CA (CA I and CA II) (Figure 1A). When nose CA was loaded and transferred to an Immobilon membrane, the antibody stained a 42-kD protein, indicating the tissue contained CA VI (Figure 1A).
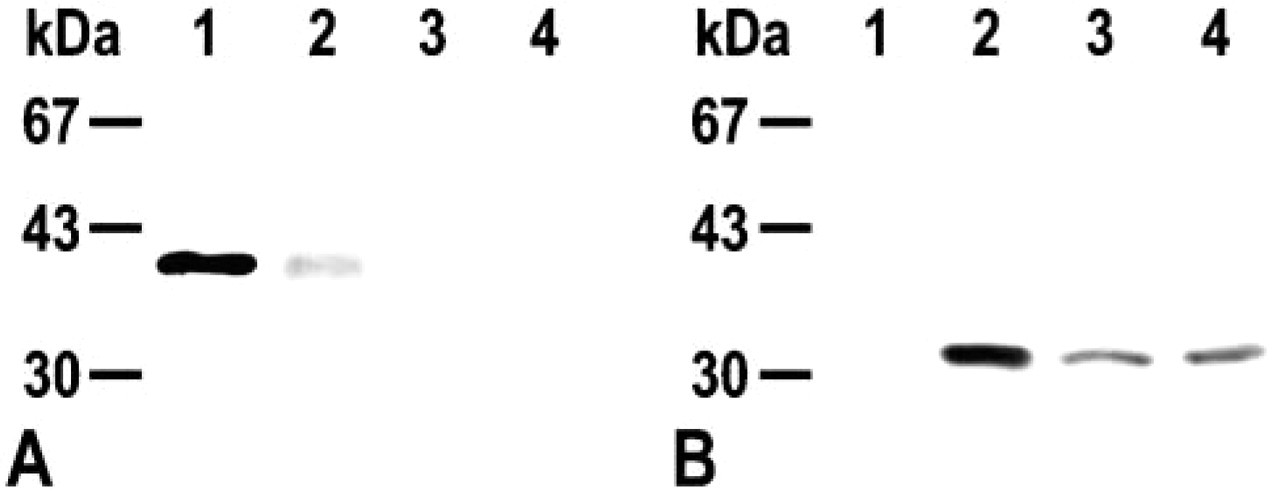
Western blotting analysis of mouse CA by anti-mouse CA VI (
RT-PCR Analysis
The nasal CA VI might arise from the nasolacrimal ducts that originate from the lacrimal glands. To determine whether the CA VI was synthesized in nasal tissue, RNA from the nasal mucosa was analyzed by RT-PCR. The PCR products of both salivary gland and nasal mucosa yielded a single band of ≃300 bp (Figure 2). They were excised from the gel, purified, and sequenced (Ogawa et al. 2002). Both nucleotide sequences were identical to the known sequence of mouse CA VI (Sok et al. 1999). Therefore, Western and RT-PCR analyses have shown that the nasal mucosa produces CA VI. We then performed IHC to determine the site at which the enzyme is produced.
Immunohistochemistry
There are several secretory structures in the mouse nose (Cuschieri and Bannister 1974). Before describing IHC of CA VI, the terminology and position of these glands are outlined briefly. Bowman's olfactory glands are situated in the lamina propria of the olfactory mucosa. Anterior and posterior glands of the nasal septum (AGNS and PGNS, respectively) underlie the respiratory mucosa of the nasal septum. AGNS are located in the anterior half and PGNS in the posterior half. PGNS, however, extend anteriorly for some distance ventral to AGNS and lie partly within the bony capsule of the vomeronasal organ, the main sensory organ for pheromones. Lateral nasal glands (LNG) form a prominent structure on the lateral wall. They are situated ventral to the ostium of the maxillary sinus and extend deeply into the connective tissue. Maxillary sinus glands (MSG) are situated in the lamina propria of the sinus mucosa. CA VI immunoreactivity was seen in the acinar cells and some ducts of LNG and AGNS (Figures 3B–3D). Bowman's glands, PGNS, and MSG were devoid of the immunoreactivity (Figures 3B and 3E). In addition to the nasal glands, CA VI immunoreactivity was seen in the nasolacrimal duct and in the mucus covering the respiratory and olfactory epithelia (Figures 3B, 3D, and 3E).
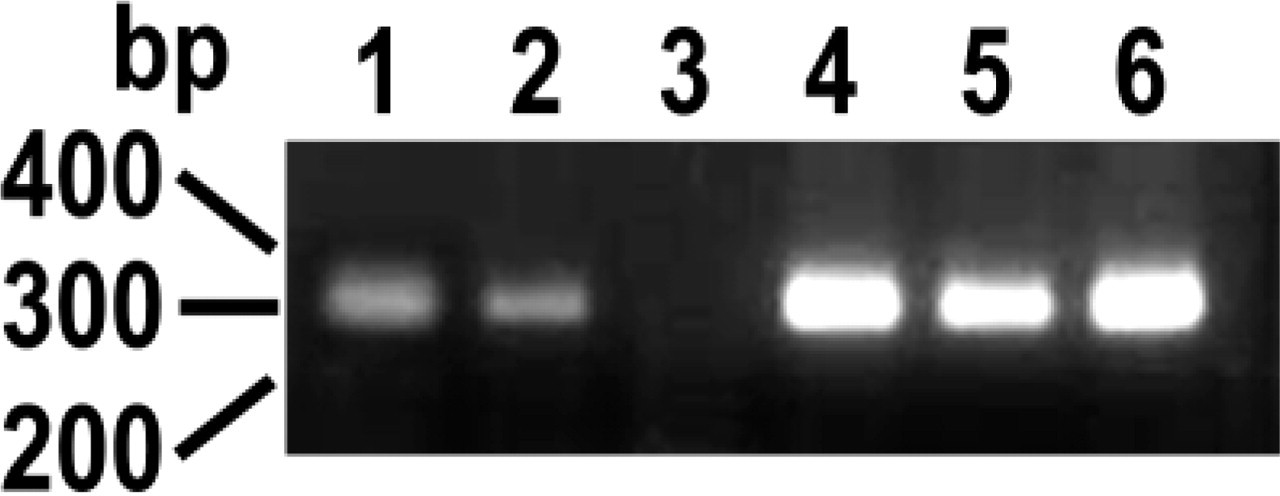
RT-PCR analysis of mouse CA VI (Lanes 1–3) and GAPDH (Lanes 4–6). Lanes 1 and 4, submandibular gland; Lanes 2 and 5, nasal mucosa; Lanes 3 and 6, liver. Aliquots (5 μl) of PCR products were run on a 2% agarose gel. CA VI mRNA is expressed by salivary gland and nasal mucosa but not by liver, whereas all the tissues express GAPDH mRNA.
An antibody to rat erythrocyte CA (Ogawa et al. 1992,1995) recognized mouse erythrocyte CA but not salivary and nasal CA VI (Figure 1B). The antibody stained the salivary gland ducts, the chondrocytes in the nasal septum, and the osteoclasts and their precursors on the bone surface, indicating that it recognized mouse CA II (Figure 3F). In the nasal mucosa, occasional cells in the olfactory epithelium were strongly stained by the antibody (Figure 3G). The characteristic bipolar appearance indicated that these cells were olfactory receptor neurons (Anholt 1993) (Figure 3H).
Discussion
The present study has demonstrated that CA VI is produced and secreted by the nasal gland of the mouse. The enzyme was produced and secreted by AGNS and LNG. CA activity has been demonstrated in Bowman's glands of the guinea pig but not those of the rat (Brown et al. 1984; Okamura et al. 1996; Coates 2001). Neither CA VI nor erythrocyte CA (CA II and probably CA I) were observed in the mouse glands. Previous carbohydrate histochemistry has suggested that the secretion of each gland structure is qualitatively different (Bojsen-Møller 1964; Cuschieri and Bannister 1974; Klaassen et al. 1981). Indeed, the secretions of PGNS and MSG include vomeromodulin and those of AGNS and LNG odorant-binding protein (OBP) (Rama Krishna et al. 1994).
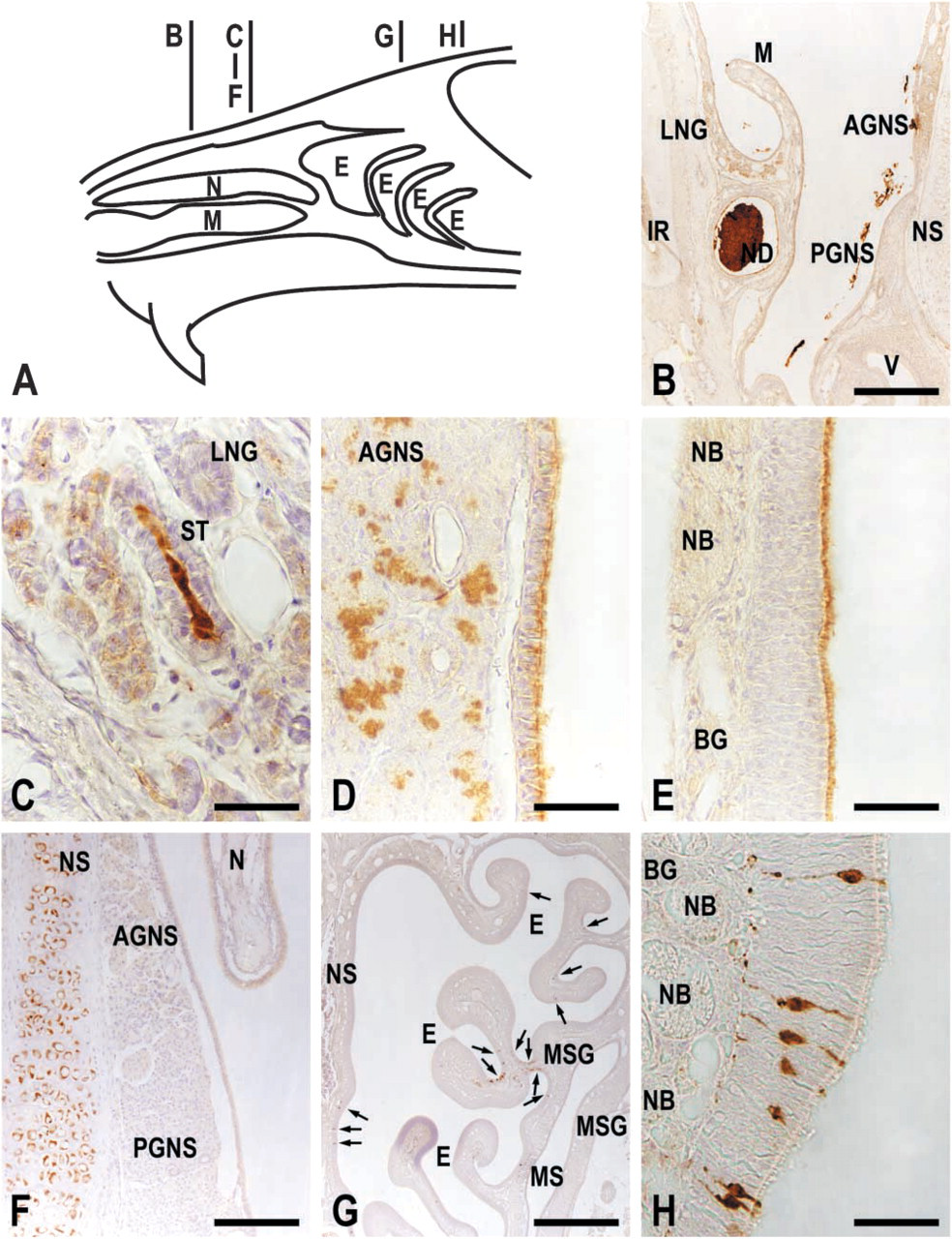
Immunohistochemistry of CA VI (
CA VI is carried through the ducts of AGNS and LNG, which open into the nasal vestibule (Warshawsky 1963; Bojsen-Møller 1964). CA VI immunoreactivity in the respiratory and olfactory mucus indicates that the enzyme has been dispersed throughout the nasal mucus, probably by atomization into the inhaled air current at a point of maximal air velocity and minimal air pressure (Bojsen-Møller 1964) and by mucociliary transport (Jones 2001). This mechanism has been proposed for the other mucous proteins such as OBP. OBP binds a variety of odorants with little specificity for chemical structures, and is believed to function in olfactory sensation (Pevsner and Snyder 1990; Pelosi 1994). Like CA VI, it is secreted by AGNS and LNG and is distributed throughout the nasal cavity (Pevsner and Snyder 1990; Pelosi 1994; Rama Krishna et al. 1994).
CA VI is presumed to provide a greater buffering capacity to the nasal mucus and thus to maintain the integrity of the underlying tissues. Inhaled acids and acids generated by CO2 (see below) would be rapidly neutralized and, at the optimal pH, environmental toxins and microbes could be efficiently eliminated by proteins in the mucus such as cytochrome P-450, lysozyme, and lactoferrin (Anholt 1993; Shusterman and Avila 2003). The CA VI activity should bring about the balanced ion concentration that is necessary for depolarization of the olfactory receptor neurons during olfactory transduction (Anholt 1993; Paysan and Breer 2001). Patients with gustatory and olfactory dysfunction after an influenza-like illness showed decreased CA in the saliva and the nasal mucus as well as apoptotic degeneration of the taste bud cells (Henkin et al. 1971,1975; Doherty et al. 1997; Henkin et al. 1999a). When the CA level was restored to normal by zinc treatment, the taste and smell acuity improved and the taste bud pathology disappeared (Henkin et al. 1999b). CA VI has been detected in taste buds, taste pores, and in saliva secreted from von Ebner's gland in direct contact with taste buds in circumvallate papillae (Leinonen et al. 2001). These results in saliva could be consistent with a role for CA VI in taste function through its action as a growth factor acting through stem cell stimulation, resulting in maturation of taste cell anatomy (Henkin et al. 1999b). The present results suggest a similar action for nasal mucus in smell function through direct effects on olfactory epithelial cells.
CA is implicated in CO2 sensing by the oral mucosa (Komai and Bryant 1993). There appear to be two types of CO2 receptors in nasal cavity: trigeminal nerve endings stimulated by high concentrations of CO2 (noxious CO2) and olfactory neurons by low concentrations of CO2 (respiratory CO2). Both receptor responses are attenuated by acetazolamide (Coates 2001), indicating that CA is also implicated in CO2 sensing by nasal mucosa. Because there is a 30-μm-thick mucus barrier between the nasal atmosphere and the nasal mucosa (Anholt 1993), it is conceivable that CA VI, through hydration of CO2, facilitates CO2 diffusion into the mucus and thus detection by the mucosa of CO2 changes in the atmosphere. In the rat and guinea pig, a small subset of the olfactory neurons possessed CA activity (Brown et al. 1984; Okamura et al. 1996; Coates 2001). The present study has demonstrated that this is also the case for the mouse, and the enzyme activity is attributed to CA II, the isozyme widely distributed throughout the body, including CO2 chemoreceptors (Lahiri 1991, Neubauer 1991, Yamamoto et al. 2003). The high activity isozyme would enable the olfactory neurons to detect low levels of CO2. CO2 in the mucus readily diffuses across the cell membrane and is catalyzed by the intracellular CA II. This increases the intracellular H+, which elicits the signal transduction process (Lahiri 1991; Neubauer 1991). To our knowledge, CA has not been localized in the trigeminal nerve endings. Therefore, they may require a different mechanism. CO2 in the mucus should be hydrated by CA VI to raise the local concentration of H+ (Shusterman and Avila 2003). For large amounts of CO2, the concentration would be sufficient to activate the nociceptors by gating of the acid-sensitive ion channels and/or H+ influx through the H+ or Na+ channel (Simons et al. 1999).