
Abstract
The human HtrA family of proteases consists of four members: HtrA1, HtrA2, HtrA3, and HtrA4. In humans the four HtrA homologues appear to be involved in several important functions such as cell growth, apoptosis, and inflammatory reactions, and they control cell fate via regulated protein metabolism. In previous studies it was shown that the expression of HtrA1 was ubiquitous in normal adult human tissues. Here we examined the expression of HtrA1 protein and its corresponding mRNA during mouse embryogenesis using Northern blotting hybridization, RT-PCR, and immunohistochemical staining analyses. Our results indicate that HtrA1 is expressed in a variety of tissues in mouse embryos. Furthermore, this expression is regulated in a spatial and temporal manner. Relatively low levels of HtrA1 mRNA are detected in embryos at the beginning of organogenesis (E8), and the levels of expression increase during late organogenesis (E14-E19). Our results show that HtrA1 was expressed during embryonic development in specific areas where signaling by TGFβ family proteins plays important regulatory roles. The expression of HtrA1, documented both at mRNA and protein levels by RT-PCR and immunohistochemistry in the developing nervous system, is consistent with a possible role of this protein both in dividing and postmitotic neurons, possibly via its documented inhibitory effects on TGFβ proteins. An exhaustive knowledge of the different cell- and tissue-specific patterns of expression of HtrA1 in normal mouse embryos is essential for a critical evaluation of the exact role played by this protein during development.
T
HtrA2, the most well-characterized member of this family, is able to induce apoptosis both in a caspase-independent manner by its protease activity and by eliminating the caspase-inhibitory activity of inhibitor of apoptosis proteins (IAPs) (Suzuki et al. 2001; Martins et al. 2002; van Loo et al. 2002; Verhagen et al. 2002).
HtrA3, highly homologous to HtrA1, appears to be involved in the formation and function of the placenta during pregnancy (Nie et al. 2003a, b).
HtrA1 is a secreted protein (Gray et al. 2000) involved in the degradation of extracellular matrix (ECM) proteins important in arthritis and in tumor progression and invasion (Clausen et al. 2002). Several studies have indicated that HtrA1, by downregulation, plays important roles in malignant progression of several tumors as well as ovarian cancers (Shridhar et al. 2002) and melanomas (Baldi et al. 2002,2003), and it has been proposed as a novel tumor suppressor gene in specific cancers (Baldi et al. 2002; Chien et al. 2004). In addition, Hu et al. (1998) have demonstrated that upmodulation of HtrA1 appears to be involved in osteoarthritis. Recently, HtrA1 tissue distribution has been demonstrated in humans (De Luca et al. 2003). This protein has a widespread pattern of expression, consistent with previous data on the RNA expression profile for this protein in human tissues (Zumbrunn and Trueb 1996; Nie et al. 2003a, b). Several points of evidence confirm that the transcription of this gene is highly regulated during development (Nagata et al. 2003) and in adult tissues (De Luca et al. 2003), suggesting that HtrA1 may exert its functions not only in neoplastic cells but also under physiological conditions (De Luca et al. 2004). Intriguingly, it has recently been demonstrated that HtrA1 is upregulated during the progress of human pregnancy, suggesting an important role of this protein in placental formation and function (De Luca et al. 2004).
Structurally, HtrA1 has a signal sequence for secretion and two distinct homology domains. The first one, at the N-terminal region, is homologous to mac25, a gene product related to insulin-like growth factor-binding protein (IGFBP) (Swisshelm et al. 1995), and to follistatin, an activin-binding protein (Kato et al. 1996). This specific domain probably makes this protein able to regulate many biological processes, perhaps modulating several growth factor system as well as IGF and the activin/inhibin system (Petraglia et al. 1998; Hu et al. 1998). The second domain shows homology greater than 40% to bacterial serine protease and allows its proteolytic activity (Hu et al. 1998). Among two distinct domains, a Kazal type of protease inhibitor motif is contained. The presence of this inhibitory motif suggests that human HtrA1 may function as a self-regulating enzyme or in regulation of other serine proteases (Hu et al. 1998).
To clarify in depth the exact roles of HtrA1, here we have investigated HtrA1 localization during mouse development with particular regard to the nervous system.
Materials and Methods
Animals and Tissue Preparation
Swiss outbred mice were housed and handled according to the international animal ethics guidelines on the care and use of laboratory animals. Adult female mice (6–8 weeks old) were mated with fertile males of the same strain to obtain pregnant mice. The finding of a vaginal plug was considered as day 0 of pregnancy. Tissues from embryos at different gestational days (8–20 dpc) and from mice at postnatal day 1 were frozen in liquid nitrogen for extracting RNA and also fixed in formalin for immunohistochemistry (IHC).
Northern Blotting
Total RNA from whole frozen embryos was extracted utilizing the Atlas Pure Total RNA Extraction System (Clontech; Palo Alto, CA). RNAs were quantified spectrophotometrically and their integrity confirmed by fractionation of 1 μg of RNA on a 1% agarose gel with ethidium bromide staining. Ten μg of RNA from each sample was subjected to electrophoresis through a 1% denaturing agarose gel containing formaldehyde. RNAs were transferred overnight onto a Hybond-N+ (Amersham; Poole, UK) nylon membrane with 20 X SSC, and RNA was UV-crosslinked onto the membrane. The membrane was then hybridized at 68C using the ULTRAhyb hybridization buffer (Ambion; Palo Alto, CA) with α[32P]-dCTP random primer-labeled cDNA probes (Random Primed DNA Labeling kit; Boeheringer Mannheim, Mannheim, Germany) using 1 × 106 cpm/ml. The probe used to detect the RNA expression of HtrA1 was a cDNA corresponding to the full-length coding region of the gene codifying for HtrA1. Filters were also probed with β-actin to normalize the signals detected. After overnight hybridization, blots were washed in 2 X SSC/0.1% SDS twice at room temperature for 10 min, then in 0.1 X SSC/0.1% SDS three times at 68C for 20 min, and exposed to a Kodak X-ray film at −80C with the aid of an intensifying screen. Two different RNA preparations from each sample were used and the different patterns of expression observed were confirmed in duplicate.
Semiquantitative RT-PCR
This was performed using the Mouse Brain Developmental Rapid-Scan Gene Expression Panel from OriGene Technologies (Rockville, MD). This product contains first-strand cDNA prepared from 48 mouse brain parts of different developmental stages. The cDNAs were normalized against β-actin, serially diluted over a 2-log range, and arrayed in duplicate on a 96-well PCR plate and then dried. Individual cDNA pools were confirmed to be free of genomic DNA contamination and to contain complete reverse transcripts. Primers used to amplify a fragment of mouse HtrA1 cDNA of 726 bp in the coding sequence of the gene (from nucleotide 473 to nucleotide 1199) were sense GCAGGAAGATCCCAACAGTT and antisense GTCCTTCAGCTCTTTGGCTT. PCR was performed with 30 cycles at the following temperatures: denaturing 95C, annealing 55C, and extension 72C. The identity of the PCR fragment was confirmed by sequencing. The different patterns of expression observed were confirmed in duplicate.
In Vitro Transcription/Translation and Immunoprecipitation Assay
One μg of supercoiled plasmid encoding for human cyclin HtrA1, HtrA2, and HtrA3 was used to program a TnT rabbit reticulocyte lysate (Promega; Madison, WI) under control of the T7 polymerase in the presence of [35S]-methionine. Immunoprecipitation was carried out as described previously (Baldi et al. 1996)
Immunohistochemistry
Whole mouse embryos and tissues at different developmental stages were immediately fixed in formalin overnight at 4C. Representative sections of each specimen were stained with hematoxylin-eosin. IHC was carried out essentially as described previously (De Luca et al. 2003,2004). Briefly, sections from each specimen embedded in paraffin were cut at 5–7 μm, mounted on glass, and dried overnight at 37C. All sections then were deparaffinized in xylene, rehydrated through a graded alcohol series, and washed in PBS. PBS was used for all subsequent washes and for antiserum dilution. Tissue sections were quenched sequentially in 3% hydrogen peroxide in aqueous solution and blocked with PBS-6% nonfat dry milk (BioRad; Hercules, CA) for 1 hr at RT. Slides then were incubated at 4C overnight with affinity-purified rabbit polyclonal antiserum raised against a purified bacterially expressed glutathione-S-transferase (GST)-HtrA1 (aa 363–480) human fusion protein (Baldi et al. 2002) at final 1:10 dilution (original concentration of the immune serum 1 μg/μl) in PBS-3% nonfat dry milk (BioRad). After three washes in PBS to remove excess antiserum, the slides were incubated with diluted goat anti-rabbit biotinylated antibody (Vector Laboratories; Burlingame, CA) at 1:200 dilution in PBS-3% nonfat dry milk (Biorad) for 1 hr. All the slides were then processed by the ABC method (Vector Laboratories) for 30 min at RT. Novared (Vector Laboratories) was used as the final chromogen and hematoxylin was used as the nuclear counterstain. Negative controls for each tissue section were prepared by substituting the primary antiserum with the isotype-matched non-immune IgG. All samples were processed under the same conditions. The expression level of HtrA1-stained cells per field (X250) at light microscopy was calculated and compared in different specimens by two separate observers (A.B. and F.B.) in a double-blind fashion and was described as: score 0 (less than 1% positive cells); score 1 (1–10% positive cells); score 2 (10–20% positive cells); score 3 (more than 20% positive cells). An average of 22 fields was observed for each specimen. All values were expressed as mean ± SEM and differences were compared using Student's t-test.
Results
HtrA1 Expression in Mouse Embryos
HtrA1 mRNA was barely detectable at 8 dpc by Northern blot analysis. Its expression was clearly detectable at 12 dpc and levels increased at 16 dpc, then remaining at the same level until birth (Figure 1).
The localization of HtrA1 protein in mouse embryos at different stages of development was performed by IHC using a rabbit polyclonal antiserum produced by immunizing rabbits with a chimeric GST-HtrA1 (aa 363–480) protein. The specificity of this antiserum and its suitability for IHC were previously validated by several experiments (Baldi et al. 2002; De Luca et al. 2003,2004). Moreover, to prove that the antiserum was specific only for HtrA1, the in vitro translated products of HtrA1, HtrA2, and HtrA3 were assayed with the antiserum anti-HtrA1. As shown in Figure 2, no crossreaction was observed.
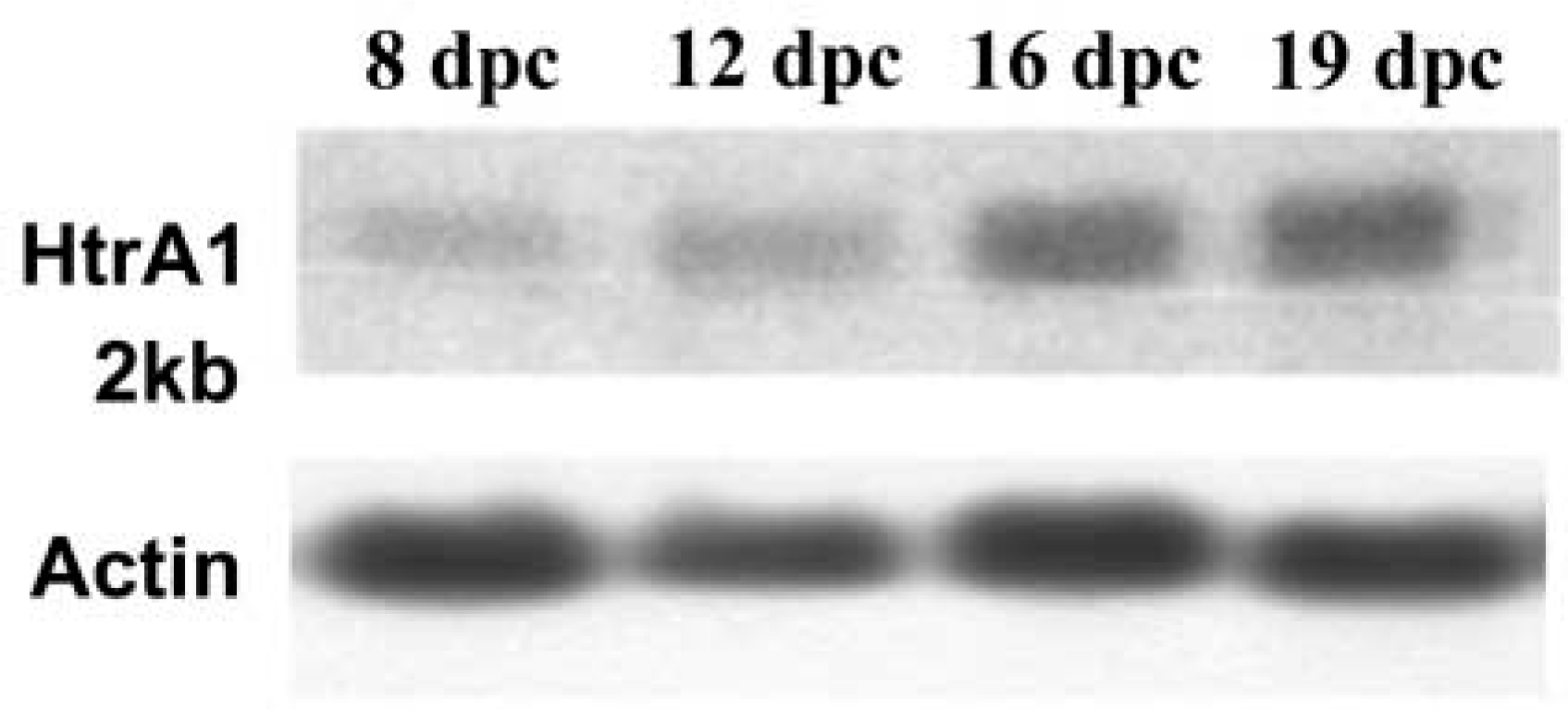
HtrA1 mRNA expression in developing embryos. Total RNA (10 μg) from whole embryos at different developmental stages was probed with 32P-labeled full-length human HtrA1 cDNA. Expression of β-actin is shown as the loading control.
In the 8 dpc embryo, HtrA1 immunopositivity was evident in neuroepithelial tissues, epithelium of gut diverticulum, mesodermal tissue, developing heart, and in dorsal aorta and vitelline artery walls. Intense immunostaining was also found in the base of allantois (Figures 3A and 3B). No immunostaining was detectable in negative control incubated with the isotype-matched non-immune IgG (Figure 3E). In the 12 dpc embryo (Figure 3D), a moderate immunopositivity for HtrA1 was localized in specific regions of brain, e.g., midbrain and neopallial cortex (future cerebral cortex) as well as in epithelium surrounding ventricles, in the pons/midbrain junction (Figure 3D), and in spinal cord. In addition, intense positivity was shown in auditory epithelium, e.g., in the cochlea and saccule, and in olfactory epithelium (Figure 3D). Intense immunopositivity for HtrA1 was evident in dorsal and lumbar root ganglia (Figure 3D). Moreover, an intermediate expression level was found in cartilage primordia of different bones and in the heart (Figure 3G). In the 19 dpc embryo (Figure 3E), we observed intense positivity in mature and outer layers of stratified epithelium of skin (Figure 3I), in epithelium of esophagus, intestine, urinary duct, and in striated and cardiac muscle. In addition, HtrA1 was moderately expressed in liver and brain. In postnatal day 1 mouse (Figure 3F), HtrA1 was highly expressed in liver (Figure 3H), in stratified epithelium of skin (Figure 3J), in epithelium of esophagus, and intestine, in cardiac and striated muscle, and in spinal cord, whereas moderate positivity was found in brain and in epithelia of lung and kidney.
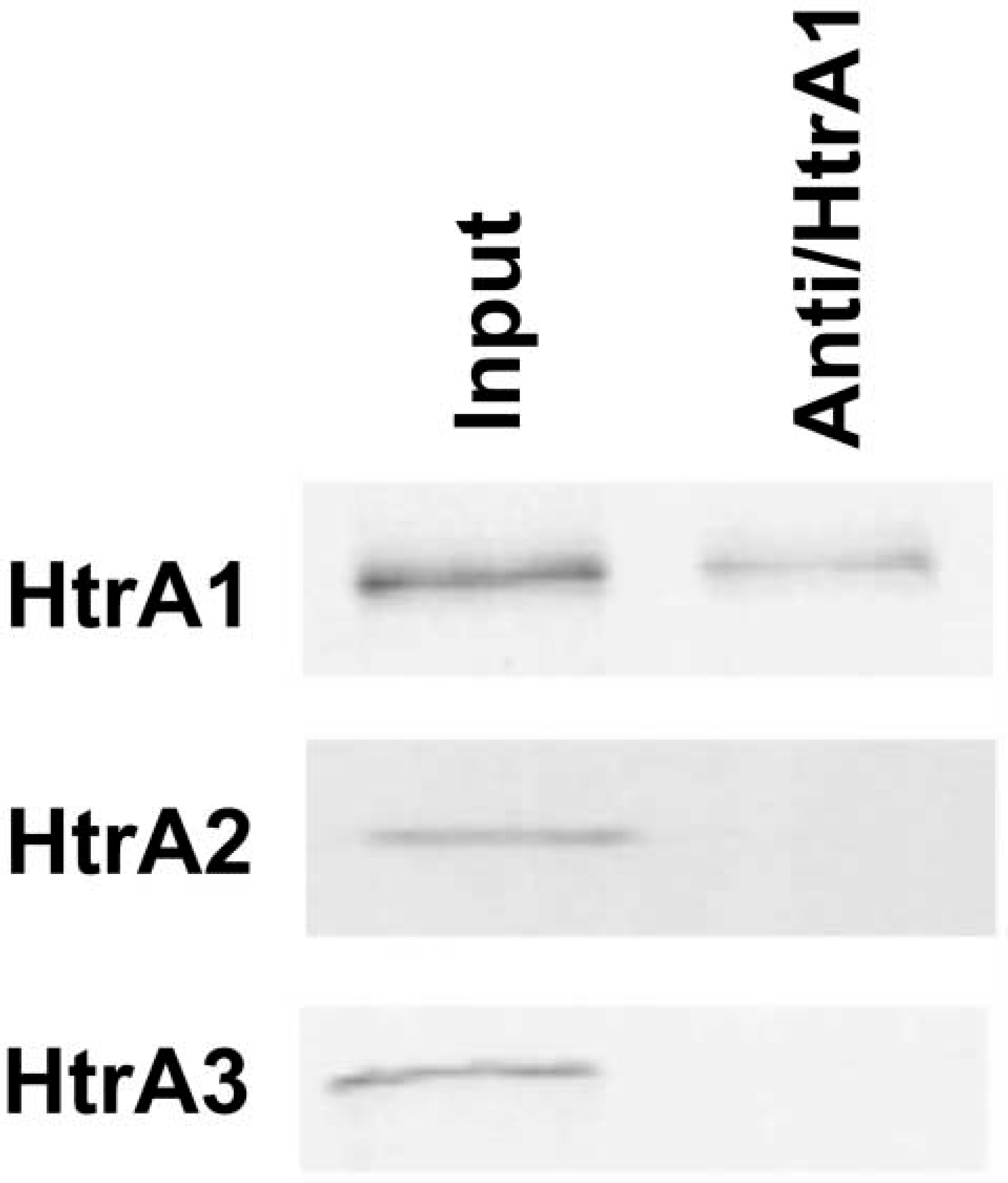
The antibody raised against HtrA1 does not crossreact with HtrA2 and HtrA3. Western blot analysis: the antiserum was able to recognize only the denatured product of HtrA1. No signal was detected with HtrA2 and HtrA3.
HtrA1 Expression in Developing Nervous System
To clarify possible roles of HtrA1 in developing nervous system, we performed a PCR analysis on 48 mouse brain parts at different developmental stages using oligonucleotide primer pairs specific for a 3′ segment of HtrA1 cDNA. The results of this analysis, using expression levels of β-actin for comparison, indicated that at 13 dpc HtrA1 was expressed at a low level in the midbrain and at a moderate level in the hindbrain. No expression was detectable in the telencephalon, in contrast to a high level of expression in the spinal cord (Figure 4A). At 15 d, expression level was low in diencephalons and midbrain whereas moderate level of expression was observed in the pons, medulla and spinal cord (Figure 4B). At 18 dpc, HtrA1 was expressed at highest levels in both frontal and posterior cortex, whereas a moderate level of expression was observed in the olfactory bulb and in the striatum, hypothalamus, medulla, and spinal cord. A low expression level was found in the thalamus and in the hippocampus, together with an undetectable level of expression measured in entorhinal cortex, midbrain, and pons (Figure 4C). The data are also presented in a graph format in Figure 4D.
To confirm the expression level of HtrA1 cDNA, we performed an IHC analysis on brain sections at different developmental stages. Figure 5A shows a representative section of brain at 14.5 dpc in which we observed a moderate/intense immunopositivity in specific regions such as striatum (Figure 5B), olfactory epithelium, vomeronasal organ, and skin (Figure 5C), together with a low immunopositivity in cartilage primordium of nasal septum and tongue muscles (Figure 5C). No immunostaining was detectable in negative control incubated with the isotype-matched non-immune IgG (Figure 5D). Intense immunopositivity for HtrA1 was also found in epithelia surrounding cerebral ventricles in brain of 12 dpc mouse embryos, together with moderate immunostaining in the pons/midrain junction (Figure 5E). In the brain of 19 dpc mouse embryos, HtrA1 was highly expressed mainly in the olfactory epithelium (Figure 5F).
Finally, Figure 6 summarizes data about patterns of expression of HtrA1 during mouse development to corroborate data obtained with Northern blotting, RT-PCR, and IHC.
Discussion
In contrast to HtrA2, the precise function of HtrA1 is still largely unknown. It is reported to be a secreted protein (Gray et al. 2000) and it has been suggested to be involved in the degradation of ECM proteins important for arthritis and for tumor progression and invasion (Hu et al. 1998; Clausen et al. 2002). The HtrA1 gene is expressed in many human tissues (Nie et al. 2003a, b). The recent development of a polyclonal antibody against hHtrA1 has contributed in defining the presence of this protein in some cell lines and in normal human tissues (Baldi et al. 2002; De Luca et al. 2003). Moreover, it has been recently demonstrated that the expression of HtrA1 in human placenta is dramatically upregulated from the first to the third trimester (De Luca et al. 2004). Finally, by electron microscopy it has been shown that HtrA1 is localized either in the cytoplasmic space of placental cells or in extracytoplasmic space of the stroma of placental villi, particularly in the spaces between collagen fibers and on collagen fibers themselves (De Luca et al. 2004). On the basis of its subcellular distribution it can be postulated that HtrA1 acts on different targets, such as intracellular growth factors or ECM proteins, to favor the correct formation/function of the placenta.
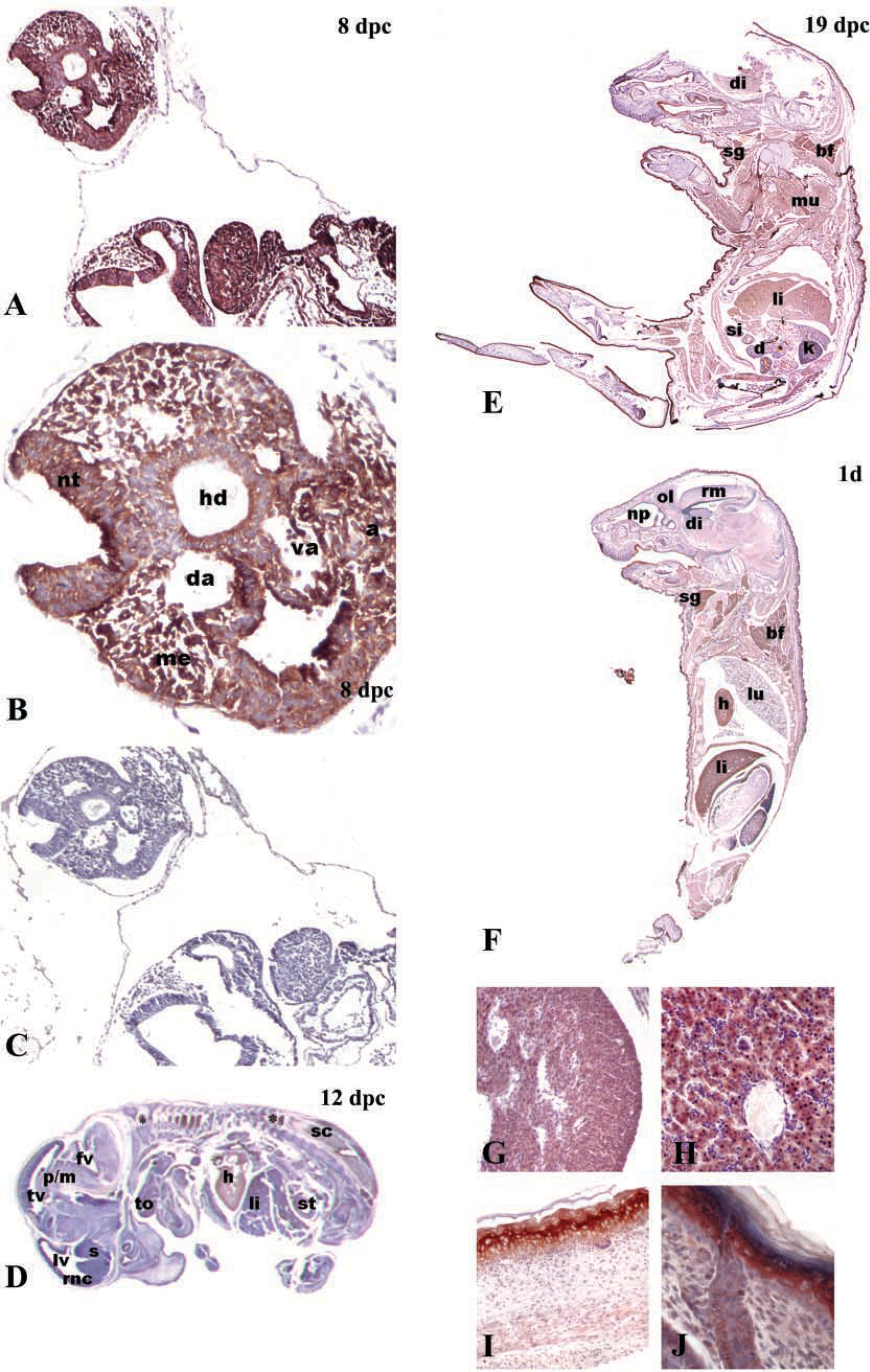
Pattern of expression of HtrA1 in mouse embryos. (
It is well known that most of the genes involved in placental development are either identical to ones used in other organ systems or are co-opted to take on new functions (Cross et al. 2003). In the present study we have examined the expression of HtrA1 protein and its corresponding mRNA during mouse embryogenesis using Northern blotting hybridization, RT-PCR, and IHC staining analyses. Our results indicate that HtrA1 is expressed in a variety of tissues in mouse embryos. Furthermore, this expression is regulated in a spatial and temporal manner. Northern blot analysis on whole embryos showed that relatively low levels of HtrA1 mRNA were detected in the embryos at the beginning of organogenesis (E8), whereas the levels of expression increased during late organogenesis (E14-E19). These data suggest that HtrA1 does not play a role in very early embryogenesis, being more involved at the beginning of organogenesis. In particular, the expression level of HtrA1 increased with gestational age in some organs as well as liver, heart, and epithelia, in contrast to a decreasing expression level in the nervous system. IHC analyses confirmed this expression pattern and were quite consistent with earlier observations (Oka et al. 2004). However, it must be emphasized that most of the previous reports used in situ hybridization to locate HtrA1 mRNA. Considering the fact that HtrA1 is believed to be essentially a secretory protein, it is not possible to simply compare data obtained by these two different methods. For example, it has been reported that mouse embryos of 12.5 and 14.5 dpc show strong expression of HtrA1 RNA in the vertebral primordia (Oka et al. 2004), whereas we detected only weak IHC staining.
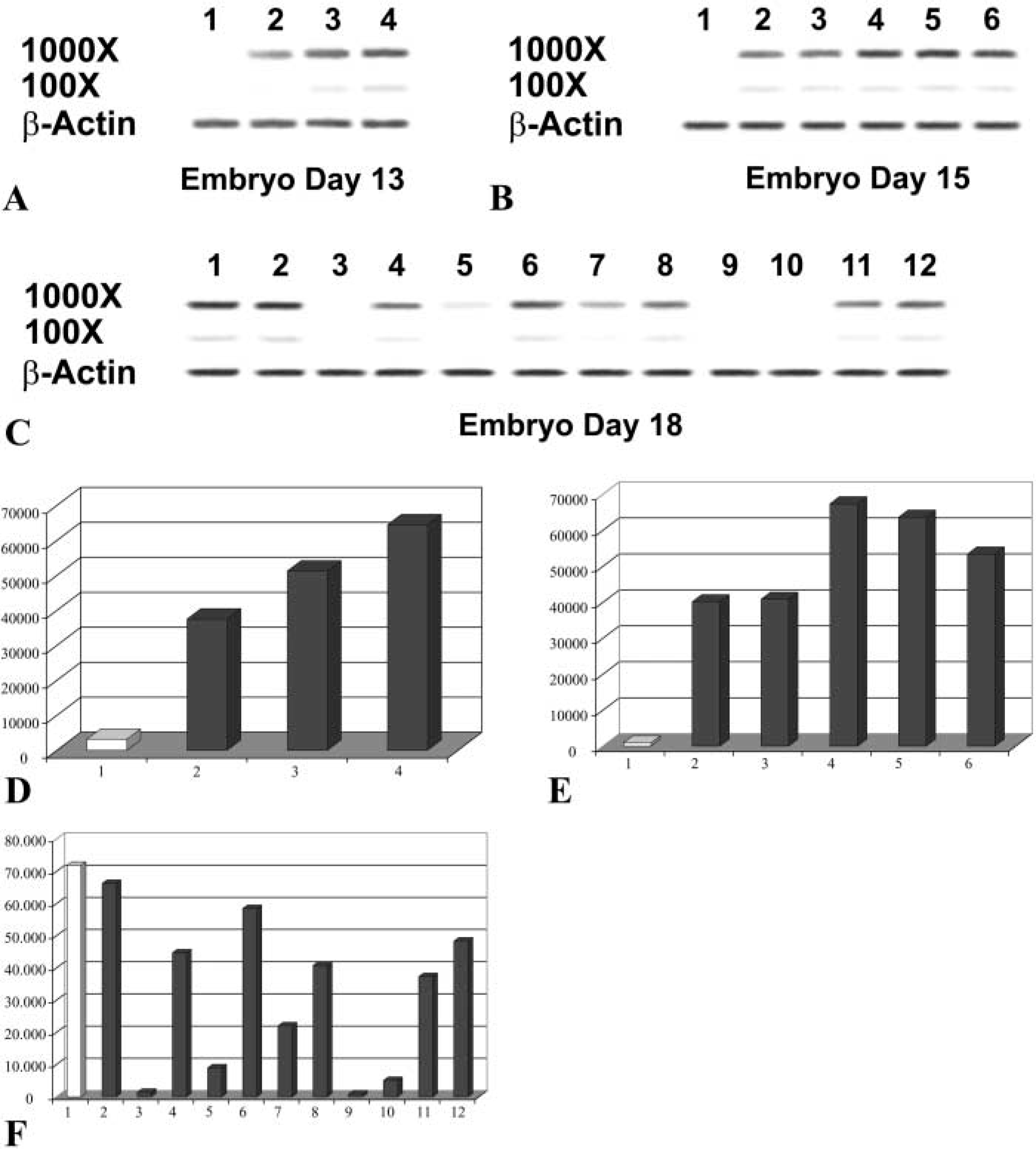
Pattern of expression of HtrA1 cDNA in mouse developing nervous system. (
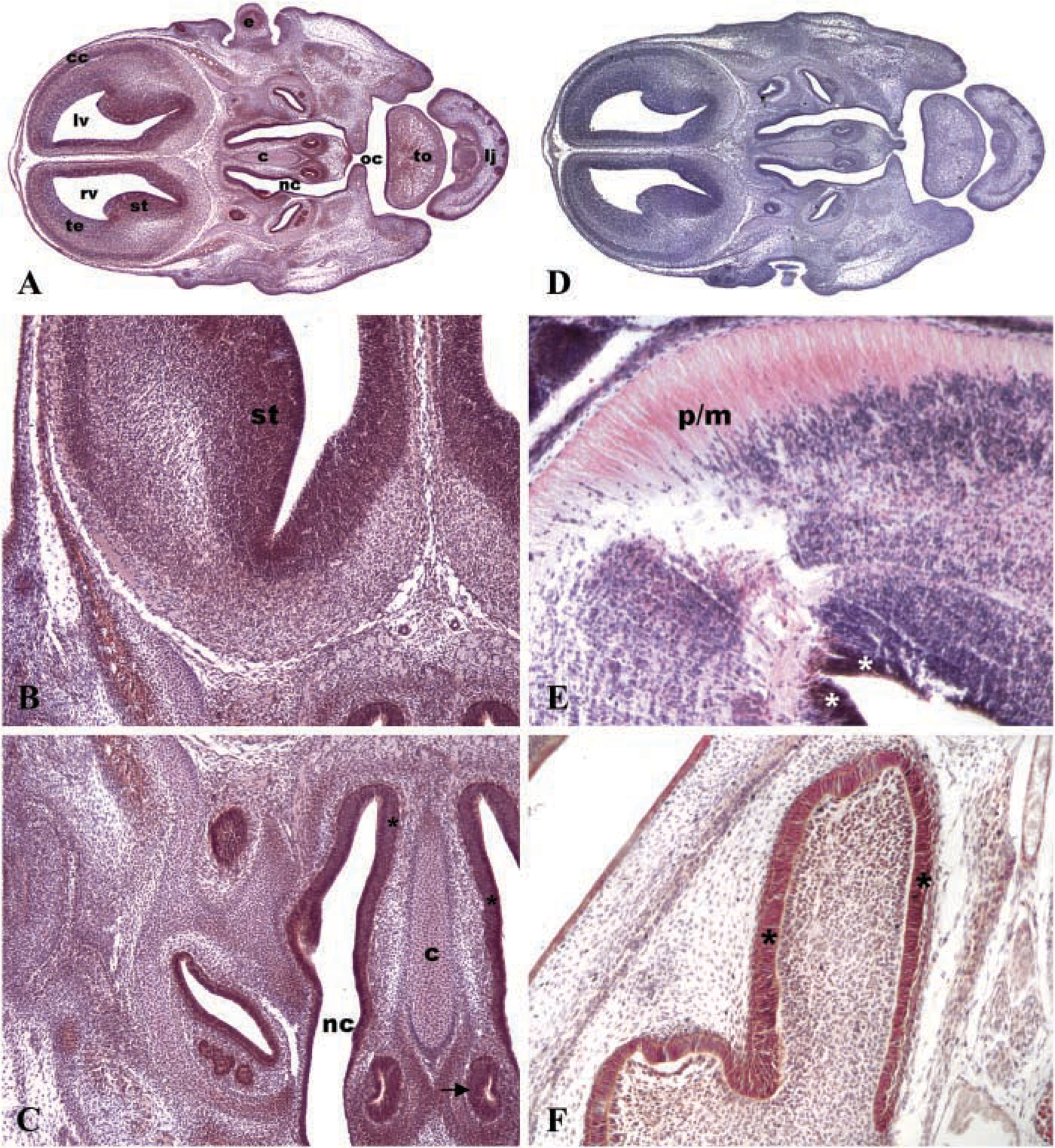
Pattern of expression of HtrA1 in brain of 14.5 dpc mouse embryo. (
A very recent study has shown that TGFβ family members are targets of this protease (Oka et al. 2004). Indeed, the TGFβ family of growth factors are important in the regulation of cell differentiation and tissue morphogenesis as well as in the control of cell division, primarily by regulating the signals by which primary and secondary induction are initiated at different stages of embryogenesis (Millan et al. 1991). Our results show that HtrA1 was expressed during embryonic development in specific areas where signaling by TGFβ family proteins plays important regulatory roles (Montuenga et al. 1998). In particular, we focused our attention on the developing nervous system. Involvement of the TGFβ family of proteins in the correct development of the nervous system is well documented (Unsicker and Strelau 2000). In particular, TGFβ proteins have anti-proliferative effects on dividing neurons in the developing nervous system, while on postmitotic neurons they have prominent modulatory effects in combination with many neurotrophic cytokines (Krieglstein et al. 1998). The expression of HtrA1 documented both at mRNA and protein levels by RT-PCR and IHC is consistent with a possible role of this protein both in dividing and postmitotic neurons, possibly also via the inhibitory effects on TGFβ proteins. Nevertheless, it must be noted that none of the IGFBP and KI (or follistatin-like) domains were required for HtrA1 binding to TGFβ family members (Oka et al. 2004). Therefore, it is conceivable that HtrA1 may also exert its functions through different targets.
The localization of HtrA1 in the developing nervous system raises a number of possibilities for the biological functions of this protein, including roles in neuronal migration, trophic support, axon guidance, or plasticity. To the best of our knowledge, this is the first report describing the localization and distribution of HtrA1 in developing mouse nervous system. An exhaustive knowledge of the differential cell- and tissue-specific patterns of expression of HtrA1 in normal mouse embryos is an essential requirement for a critical evaluation of the exact role played by this protein during development. Nevertheless, a careful identification of the physiological substrates of this protease still remains the most important step towards understanding of its biological functions.
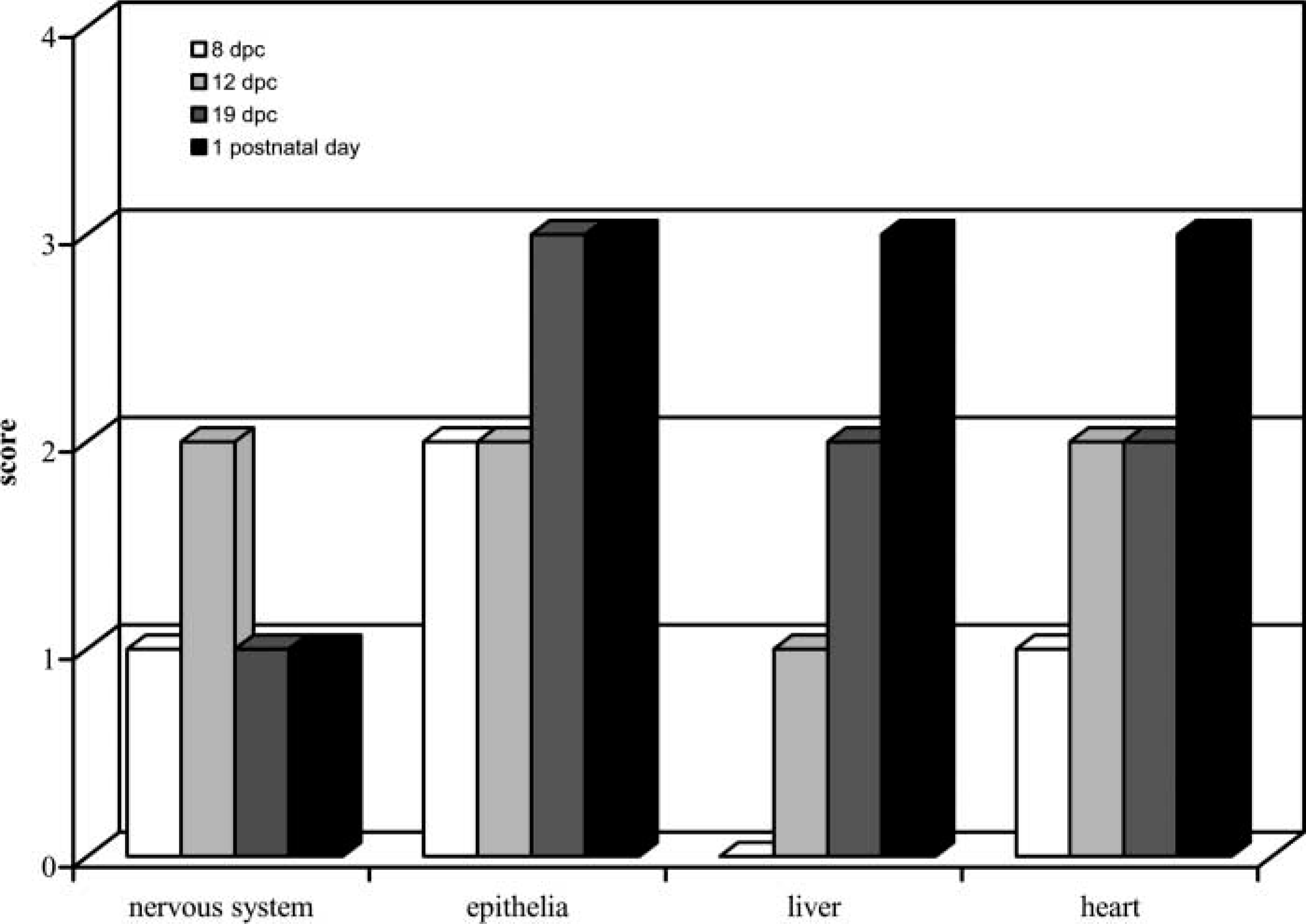
Time course of HtrA1 pattern of expression during mouse development. Ordinate: immunoreactivity scored as described in Materials and Methods. Abscissa: different tissues of different mouse developmental ages.
Footnotes
Acknowledgements
This work was supported in part by AIRC and Ministero della Salute grants to MGP, by Futura-ONLUS, Second University and AIRC grants to ADL and AB.
We thank the International Society for the Study of Comparative Oncology Inc. (president H.E. Kaiser) for its continuous support and Dr L.M. Martins for the generous gift of the HtrA2 cDNA. We also thank Mr Giuseppe Falcone for his contribution to the images elaboration.