
Abstract
We investigated the development of innervation of the pulmonary neuroendocrine cell (PNEC) system composed of single cells and organoid cell clusters, neuroepithelial bodies (NEB) in rabbit fetal and neonatal lungs. To visualize the nerve fibers and their contacts with PNECs/NEBs, we used confocal microscopy and multilabel immunohistochemistry (IHC) with pan-neural marker, synaptic vesicle protein 2 (SV2), and serotonin (5-HT) as markers for PNECs/NEBs, and smooth muscle actin or cytokeratin to identify airway landmarks. The numbers and distribution of PNEC/NEB at different stages of lung development (E16, 18, 21, 26, and P2) and the density of innervation were quantified. First PNECs immunoreactive for 5-HT were identified in primitive airway epithelium at E18 as single cells or as small cell clusters with or without early nerve contacts. At E21 a significant increase in the number of PNECs with formation of early innervated NEB corpuscules was observed. The overall numbers of PNECs/NEBs and the density of mucosal, submucosal, and intercorpuscular innervation increased with progressing gestation and peaked postnataly (P2). At term, the majority of NEBs and single PNECs within airway mucosa possessed neural contacts. Such an extensive and complex innervation of the PNEC system indicates a multifunctional role in developing lung and during neonatal adaptation.
T
Because NEBs are prominent in fetal/neonatal lungs, their role may be particularly important during lung development and neonatal adaptation (Cutz et al. 1984; Cutz and Jackson 1999). We have previously documented the distribution and frequency of NEBs in developing rabbit lung, with peaking of NEB numbers soon after birth and decline in the adult (Cho et al. 1989).
Here we investigated the ontogeny of innervation of the PNEC system in developing rabbit lungs because this species has been used extensively to characterize NEB function both in vivo (Lauweryns and Cokelaere 1973; Lauweryns et al. 1978) and in vitro (Youngson et al. 1993; Fu et al. 2002). Furthermore, there are well-known species differences in lung innervation (Dey and Hung 1997). We examined lung samples from the fetal to the early postnatal period using confocal microscopy on sections immunostained with an antibody to a pan-neural marker, synaptic vesicular protein 2 (SV2) (Xu and Bajjalieh 2001), combined with immunostaining for 5-HT, a marker of PNECs/NEBs (Cho et al. 1989). To define airway anatomic landmarks, we used an antibody to smooth muscle actin as a marker of airway smooth muscle and cytokeratin as a general epithelial cell marker. Our findings indicate that PNECs/NEBs differentiate in primitive airway epithelium concurrently with the appearance of mucosal innervation. During the late fetal/early neonatal period, most solitary PNEC and NEB clusters become innervated, with highest intramucosal nerve density occurring on postnatal day 2. We also found that the majority of solitary PNECs in the major bronchi appear innervated and that individual NEBs may be interconnected via submucosal neural connections. These observations suggest that the PNEC system might play an important role in neurohormonal regulation of airway function in developing lung, particularly in the transduction of intraluminal stimuli (i.e., hypoxia) via complex innervation.
Materials and Methods
Animals and Tissue Preparation
Timed-pregnant New Zealand rabbits at E16, 18, 21, and 26 days of gestation, and neonates on day 2 and 7, as well as young adults on postnatal day 30, were obtained from the Riemens Fur Ranch, (Waterloo, ON, Canada). All animal studies were approved by our institutional Animal Care Committee in accordance with accepted guidelines for animal care and experimentation. Does were sacrificed by lethal injection of euthanol and the fetuses by decapitation. Dissected lungs were washed three times in CO2-independent medium and embedded in polyethylene glycol (OCT medium) (Lab-Tek Products; Naperville, IL) and then snapfrozen on dry ice. All tissue blocks were sealed and stored at − 80C until use.
Immunofluorescence Labeling
Cryostat sections of medial lobes were cut at 100 μm under low working temperature (–12C). The sections were immediately transferred to a dish with zinc formalin fixative (Newcome Supply; Middleton, WI) at room temperature (RT). After three changes of fresh fixative (10 min each at RT), the sections were washed in PBS and stored at 4C. Immunofluorescence labeling was performed on sections permeablized with 1% Triton X-100 in PBS for 10 min and incubated with 0.03% of H2O2 in PBS for 5 min to block endogenous peroxidase. To suppress nonspecific binding, the sections were incubated in 20% normal donkey serum in 4% BSA plus avidin/biotin blocking solution for 60 min at RT. This was followed by incubation with two primary antibodies, first with mouse monoclonal anti-synaptic vesicle 2 (SV2) antibody (1:200 dilution; Developmental Studies Hybridoma Bank, Iowa City, IA) followed by goat anti-5-HT antibody (1:500 dilution; DiaSerin, Stillwater, MN) at 4C overnight using an orbital shaker. As secondary antibodies we used donkey antibodies conjugated with either horseradish peroxidase (HRP) for SV2 or fluorescein isothiocyanate (FITC) for 5-HT (Jackson ImmunoResearch; West Chester, PA) for 2 hr at RT.
The signal for SV2 immunolabeling was enhanced by a catalyzed reporter deposition (CARD) amplification system. The N-hydroxysuccinimide ester of biotin [sulfosuccinimidyl-6-(biotinimide) hexanoate] (BIO-NHS) (Boehringer-Mann-heim; Mannheim, Germany) was coupled to tyramide-HCl (Sigma Chemical; St Louis, MO) according to the method of Hopman et al (1998) to produce a biotin/tyramide substrate for HRP reaction. After incubation with HRP-conjugated secondary anti-mouse IgG antibody, this complex was then amplified with the biotin/tyramide substrate in sodium borate buffer, pH 8.3, with 0.0003% H2O2 for 10 min at RT. To visualize the signal, we applied a 1:400 dilution of either streptavidin/FITC conjugate or streptavidin-Texas Red X conjugate (Molecular Probes; Eugene, OR) for 30 min at RT. To identify anatomic landmarks i.e., airway wall smooth muscle or airway epithelium, we used a monoclonal antibody against smooth muscle actin or low molecular weight cytokeratin labeled with FITC or RITC (Sigma Chemical).
Confocal Microscopy
All preparations for immunofluorescence double or triple labeling were examined under a Leica confocal laser scanning microscope (model TCS 4D) with SCANWARE software. The excitation wavelengths of the krypton/argon laser were 488 nm for FITC and 568 nm for Texas Red. The tissue slices were optically sectioned by scanning at depth of focus (in steps of 1 μm) under a ×25 objective lens to follow the path of the neural network towards NEBs or PNECs in relation to the airway smooth muscle or airway epithelium. Many fields were scanned and multiple areas selected for detail observation of NEB and PNEC innervation under a ×40 objective lens. The maximal intensity of the corresponding pixels in each optical section was used to generate a single topographic image (2-D projection) from a stack of images obtained at varying depth (41 μm on average). FITC- or Texas Red-labeled images of nerves (including NEBs, PNECs, and ganglia) were merged with images of airway smooth muscle or epithelium (either RITC or FITC) to produce a dual-color nerve/smooth muscle airway epithelium image. Image processing including merging and montage of fields was carried out using Adobe PhotoShop 5.0 software. Single fields were collated and assembled, creating a comprehensive bronchial nerve/smooth muscle airway epithelium image.
Morphometric Analysis
For quantification of PNECs and NEBs we used a method according to Van Lommel and Lauweryns (1997). The integrated surface area of bronchioles of different sizes, expressed in square millimeters of the section (100-μm thickness) was measured using the NIH-Scion program standardized by an internal scale bar in each counted confocal image. The numbers of PNECs/NEBs were assessed in three sections of medial lobe immunostained for SV2 and 5HT. The total number of NEBs and PNECs in each section was divided by the integrated surface area and the relative number expressed as a mean ± SD per mm3 of lung tissue based on calculated volume of three thick (100-μm) frozen sections. Morphometric analysis of the density of airway innervation was performed using NIH-Scion Image software as modified by Baluk et al. (1992). The density of innervation of airway smooth muscle and submucosa was defined by the rectangular selection tool as integrated reading density per 100 μm2 of nerve immunoreactivity in airways of different sizes. The sizes of airways were determined from measurement of the external diameter of the smooth muscle coat. The selected measured areas excluded large nerve trunks and PNECs/NEBs. Because immunostaining with pan-neural marker SV2 does not differentiate between immunostaining of nerve fibers and cytoplasmic staining of PNECs/NEBs, we subtracted the overlapping signal (SV2+5-HT) from total SV2 immunoreactivity. To determine the nerve density in corpuscular NEBs, the areas of signal overlap (yellow) due to 5-HT (red) and SV2 (green) were highlighted by a Magic Wand tool from Photo Shop 5.0. The rest of the green images were converted to black- and white-field and then measured using NIH-Scion Image and expressed as intracopuscular nerve density in individual NEB (n = 5 for each developmental stage). The real standard regions measured were normalized by an internal scale bar in each confocal image. The density of NEB innervation was measured under ×40 objective from a stack of images (5 μm) with five single laser scans. A freehand selection tool in the NIH-Scion program was used to highlight the whole NEB defined by positive immunostaining for 5-HT (Texas Red), and then switched to the channel showing SV2 expression normalized by an internal scale bar.
Results
Development and Innervation of PNECs/NEBs
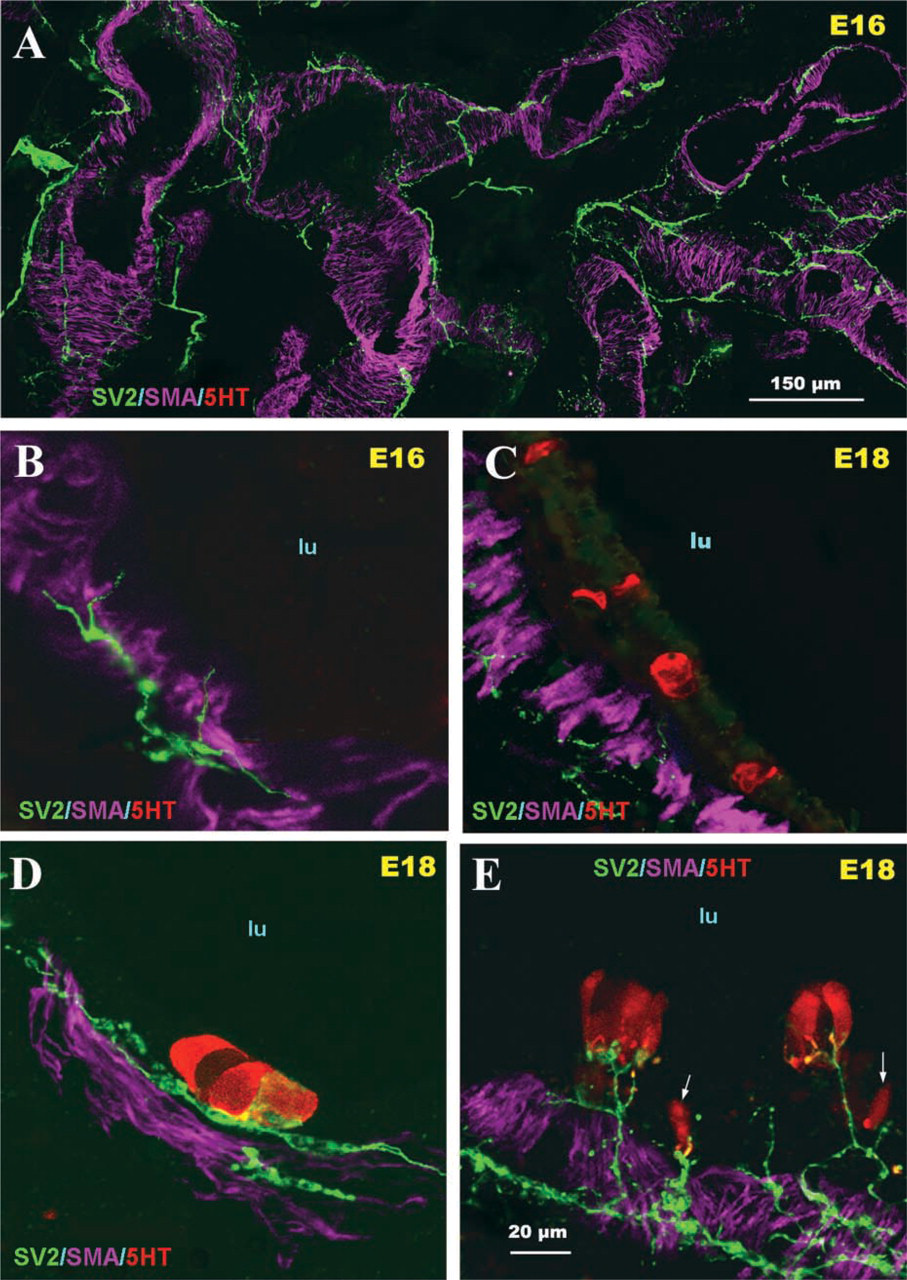
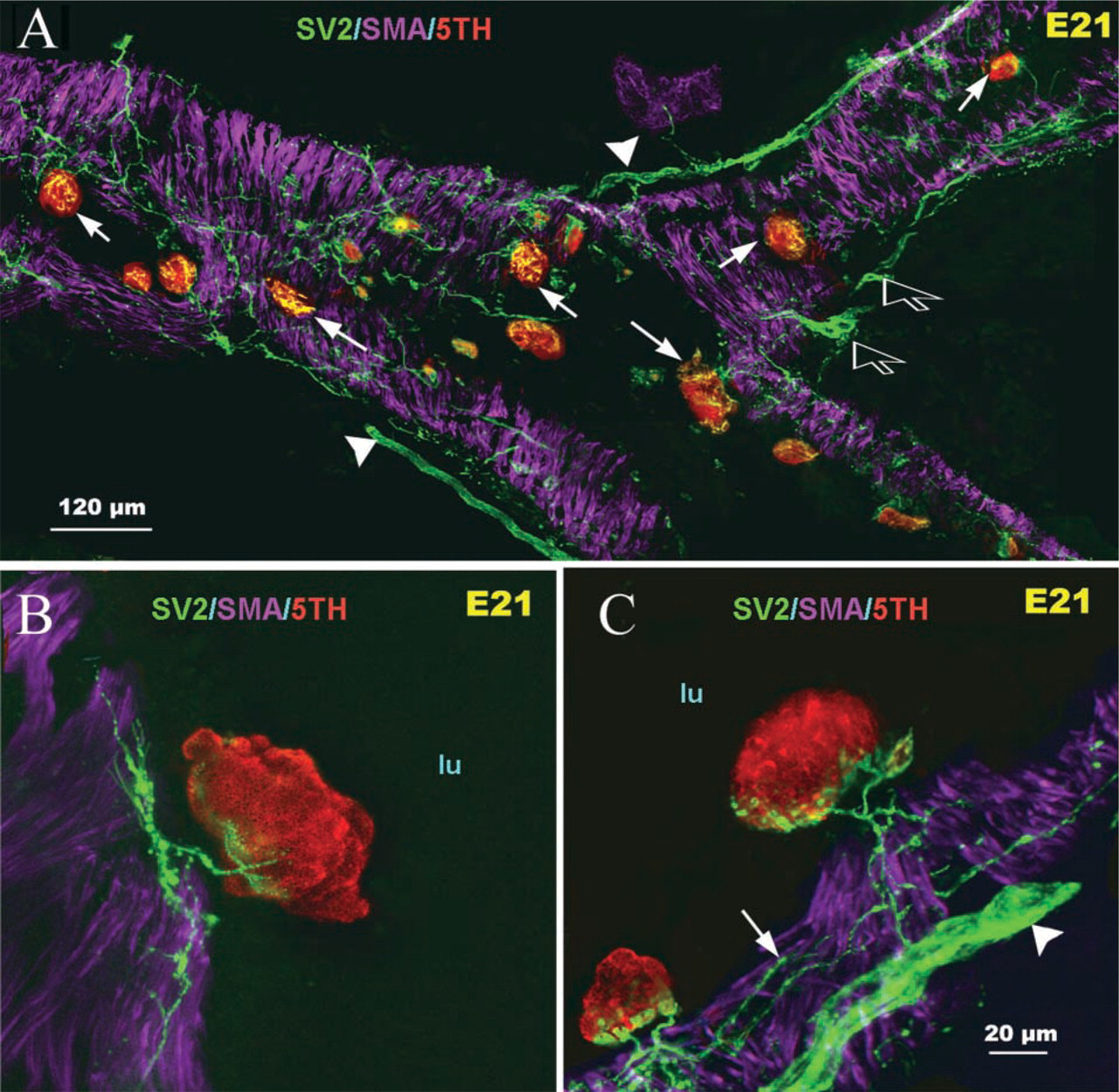
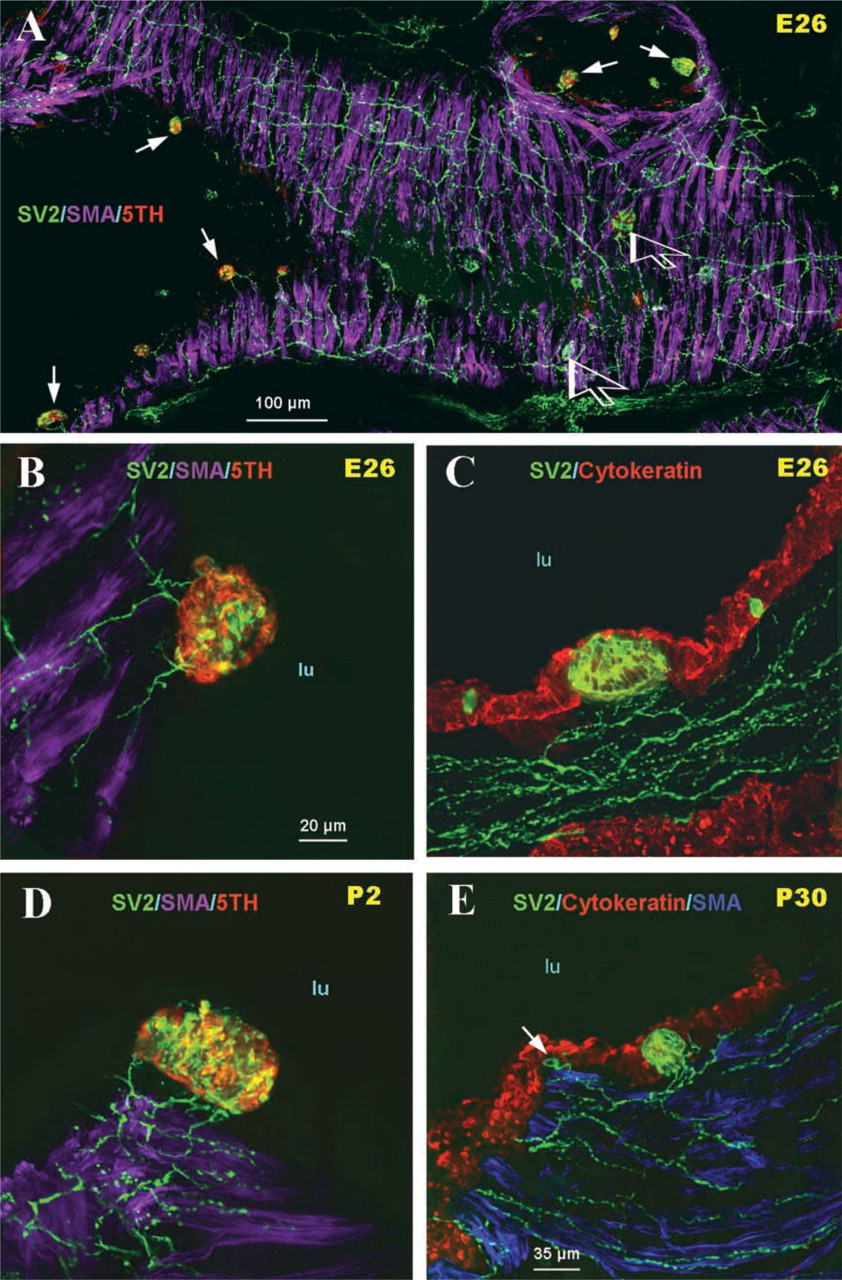
During the early stages of lung development (E16), large nerve trunks were apparent in the lung hilus, which subsequently grew into the lung parenchyma along the main airway branches and generated an extensive neural plexus within developing airway smooth muscle (Figure 1A). At this stage of development, neither 5-HT-immunoreactive cells within airway epithelium nor mucosal innervation was observed (Figure 1B). At E18, a few solitary PNECs and small PNEC cell clusters immunoreactive for 5-HT were apparent in the primitive airway epithelium. Some were without neural contacts (Figure 1C), whereas others were forming early contacts with submucosal nerve endings (Figures 1D and 1E). At E21, a significant burst in the number of 5-HT immunoreactive PNECs occurred and the first PNEC clusters forming compact NEBs as well as small peribronchial ganglia were noted (Figure 2A). This was followed by a gradual increase in the density and complexity of airway innervation, including PNECs/NEBs. Fine nerve fibers entering the base of NEBs and forming early intracorpuscular branches were evident (Figures 2B and 2C). In an appropriate plane of section, neural connections between several NEBs in close proximity became apparent (Figure 2C). At E26, the fine nerve fibers around the smooth muscle coat became more complex and more uniformly distributed in larger airways (>250 μm in diameter) (Figure 3A). At this stage of development a more prominent submucosal nerve plexus was evident, with several nerve fibers entering corpuscular NEBs (Figures 3B and 3C). By postnatal day P2, NEBs appeared fully innervated and exhibited an extensive intracorpuscular network of fine fibers (Figure 3D). The nerve fibers entering NEBs originated from a complex submucosal neural network of fine varicose fibers in apparent continuity with nerve fibers innervating the bronchial smooth muscle (Figures 3C-3E).
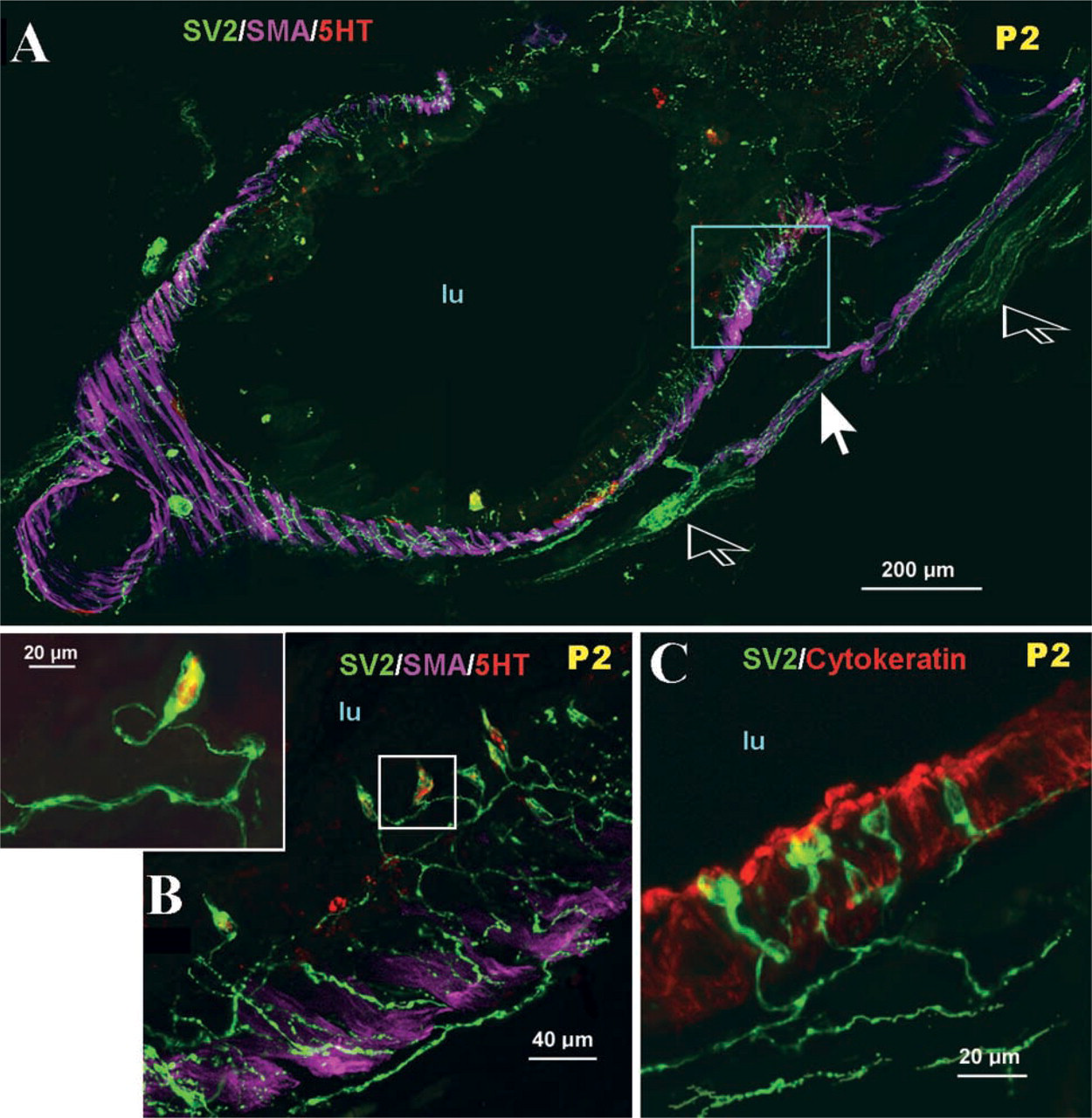
During the early postnatal stage, large numbers of solitary PNECs appeared in the mucosa of large bronchi (Figure 4A). Most single PNECs formed a thin apical or basal process extending to the airway lumen or, alternatively, towards the basement membrane, and most of them appeared innervated by a complex network of interconnecting fine varicose nerve fibers originating in the submucosa (Figures 4B and 4C).
Quantitative Studies
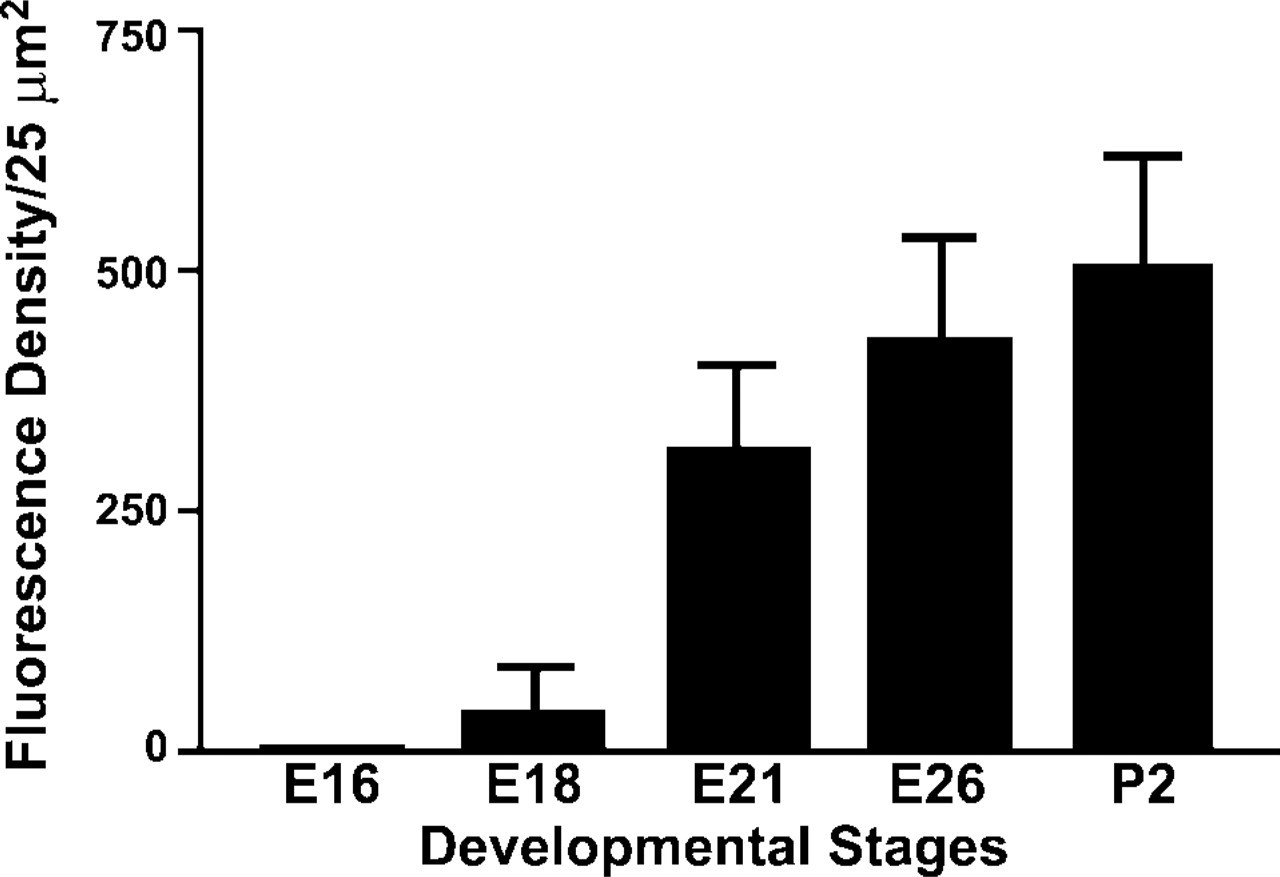
In agreement with previous studies (Sorokin et al. 1997), PNEC/NEB development progressed in a proximal to distal airway pattern. At E18 only occasional isolated PNECs or small PNECs clusters were identified in the mucosa of the larger proximal airways. The mean number of PNECs/mm3 of lung parenchyma was 20 ±6 in airways >250 μm and 5±3 in airways <250 μm (Table 1). At E21, the numbers of both solitary PNECs and early NEBs had increased significantly in most airways present at this stage of lung development. In airways >250 μm, the mean number of PNECs/mm3 was 311 ±49, whereas in airways <250 μm it was 228±33. At the E21 stage of lung development, the first corpuscular NEBs were observed and their mean numbers (i.e., 168 vs 199) were comparable among airways of different sizes (Table 1). At E26, many PNECs (∼698 ±145) appeared in large proximal airways (>500 μm), whereas in more distal airways the number of PNECs remained relatively constant (∼158±60 in >250 μm vs ∼255±32 in <250 μm). The mean number of NEBs (∼377± 55) was higher in more distal airways (<250 μm) compared with proximal airways (∼228±43 in >250 μm) and in airways >500 (∼196±29) (Table 1). At postnatal day P2 there was an increase in the frequency of PNECs in large airways and NEBs in smaller airways (Table 1).
(
(
(
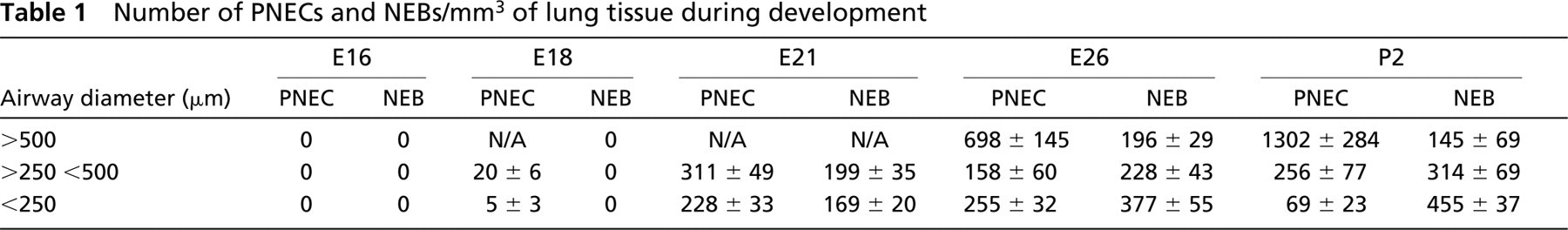
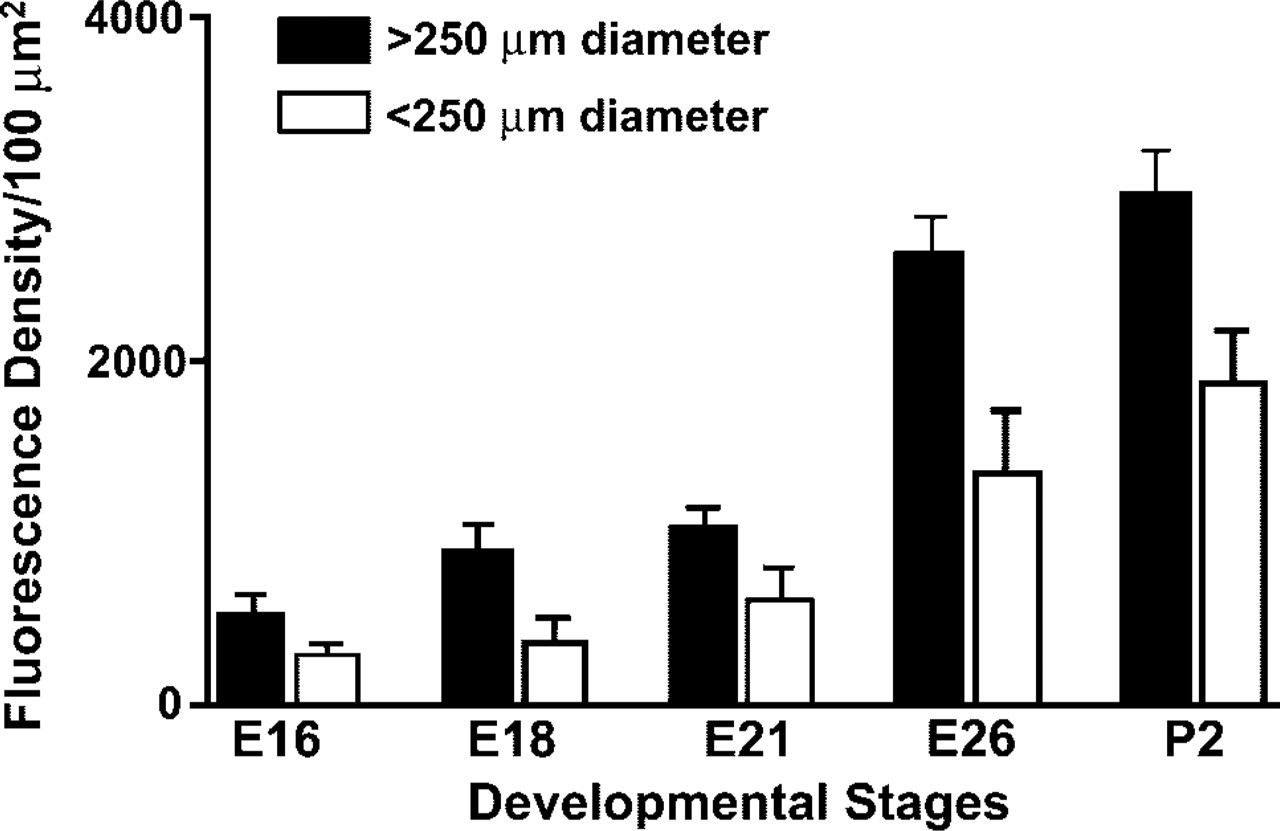
The quantitative assessment of the density of airway innervation was expressed as integrated fluorescence density of SV2-immunolabeled nerve fibers. The overall density of innervation of the airway submucosa tended to be higher in larger airways (>250 μm) during all stages of lung development, with significant increases close to term and postnatally (Figure 5). The density of SV2-immunolabeling in NEB reached its plateau by postnatal day P2, indicating that NEBs become fully innervated after birth (Figure 6).
Discussion
The development of PNECs/NEBs in humans and various animal species has been reported in a number of previous studies (Cutz et al. 1984; Cho et al. 1989; Van Lommel and Lauweryns 1997). Quantitative studies on the distribution and frequency of NEBs in late fetal and neonatal rabbit lungs revealed striking developmental changes, with prominence of NEBs during the perinatal period and a decline postnatally (Cho et al. 1989). Similar findings were reported for developing human lungs, with peaking of PNECs/NEBs numbers at birth and decline with advancing age (Cutz et al. 1984). Other studies have shown that the relative number of PNECs/NEBs in human lung did not change over time but rather became “diluted” with postnatal lung growth (Gosney 1993). These findings were confirmed by more recent investigations comparing the number of PNECs/NEBs in animal species with different lengths of gestation (Van Lommel and Lauweryns 1997). Stereological estimate of the total volume of NEBs in neonatal rabbit lung was three times that of the carotid bodies (the principal arterial chemoreceptor) and was close to the volume of the parathyroid glands, indicating a significant physiological function for these cells (Bolle et al. 1999). Our quantitative studies on the frequency and distribution of PNECs/NEBs in developing rabbit lung reported here are in general agreement with the above studies. Sorokin et al. (1997) described centrifugal waves of PNEC/NEB differentiation progressing from the trachea to the intrapulmonary airways. The present study suggests that in the developing rabbit lung the first significant burst of PNEC differentiation occurs at E21, corresponding to the late glandular and early cannalicular period, before extensive airway branching and development of terminal airways. The first PNECs immunoreactive for 5-HT were detected at E18, with some PNECs without innervation whereas others showed early contacts with submucosal nerve endings. This observation suggests several possibilities, including that the differentiation of PNECs may be independent of innervation, may be synchronized with mucosal innervation or, alternatively, may be innervation-dependent. The former interpretation is supported by studies of Sorokin et al. (1993), using explants of rat fetal lungs, that have shown that the differentiation of PNECs proceeded at the same rate in vitro as in intact animals. These lung explants also developed mucosal nerve fibers derived from intrinsic neuroblasts, but their possible role in PNEC differentiation was not explored. More recently, Ito et al. (1997) have shown differentiation of PNECs in explants of hamster lung buds devoid of mesenchyma and nerves, suggesting that PNEC precursor cells and endocrine differentiation are intrinsic to lung epithelium. Therefore, it is conceivable that neural contacts may not be an absolute requirement for PNEC differentiation but that they may be important for functional reasons in an intact animal.
The first intramucosal nerve fibers in close contact with 5-HT-immunoreactive PNEC were noted at E21 when the first NEB clusters became apparent, suggesting that these structures may guide their own innervation. Weichselbaum et al. (1996) and Sparrow et al. (1999) reported studies on the development of innervation of airway smooth muscle coat in human and porcine lungs using confocal microscopy and various neural markers. These studies demonstrated an extensive neural network enveloping the fetal airways, supporting earlier physiological studies on fetal lungs that reported strong smooth muscle constrictor responses after electrical or agonist stimulation (Sparrow et al. 1994). This indicates the presence of functional innervation of the airways in early fetal life. There are no comparable studies on the innervation of PNECs/NEBs in the developing lung, and its possible contribution to the airway constrictor responses is not known.
(
Number of PNECs and NEBs/mm3 of lung tissue during development
The density and complexity of NEB innervation increased with advancing gestation, reaching a plateau postnatally. These findings are in agreement with studies of Bolle et al. (2000), who used electron microscopy to assess the density of NEB innervation in neonatal rabbit lungs. In this study, the density of afferent-like and efferent-like nerve endings in NEBs was higher compared to that of the carotid bodies in the same animals, suggesting more advanced functional maturation in NEBs. Overall, these findings are consistent with the postulated role of NEBs as auxiliary O2 sensors, particularly at birth when carotid body function is relatively immature.
The use of immunostaining for SV2 and confocal microscopy on thick frozen sections allowed a detailed examination of airway mucosal innervation, including PNECs/NEBs, not feasible by conventional microscopy. In neonatal rabbit lungs we observed that the great majority of NEBs were innervated and that individual NEBs may be interconnected with other NEBs within the same airway. Such an arrangement is reminiscent of “connection in parallel”, perhaps allowing signal amplification from a dispersed population of small cell clusters. Other studies reported variable innervation of NEBs in adult rat lungs with a subpopulation lacking neural connections (Larson et al. 2003). These were interpreted as representing possible different NEBs functional subgroups, i.e., innervated NEB functioning as chemoreceptors, whereas those without innervation representing endocrine/paracrine cells (Adriaensen et al. 2003; Brouns et al. 2003; Larson et al. 2003). However, the possibility that this apparent lack of innervation in some NEBs may be due to technical problems or may reflect immature or senescent NEBs has not been excluded. In addition, some of these discrepancies could be related to well-known species variations in lung innervation (Dey and Hung 1997). For example, in the rat lungs NEBs show prominent CGRP-immunoreactive nerve fibers derived from the spinal ganglia, but in the rabbit lungs only sparse CGRP-immunoreactive nerve fibers are observed (unpublished observation). The present study used SV2 as a pan-neural marker that identifies all nerve fibers regardless of their neurotransmitter content. To further characterize the nerve fibers innervating NEBs and their precise origin in the rabbit, further studies are required.
Quantitative assessment of nerve density expressed as integrated fluorescence density of SV2-immunoreactive nerve fibers in airway walls of different sizes during lung development.
It appears likely that airway mucosal innervation is guided by neurotrophic factors such as glial-derived neurotrophic factor (GDNF). Studies by Tollett et al. (2002) demonstrated that exposure to GDNF in vitro enhanced neural outgrowth in organ cultures of fetal mouse lung. Furthermore, expression of both mRNA for GDNF and its receptor has been demonstrated in mouse lung (Towers et al. 1998). Most intramucosal nerve fibers appear to make contact with PNECs/NEBs rather than being randomly distributed throughout the airway mucosa, suggesting that specific neurotrophins could be secreted by PNECs/NEBs, a highly specialized population of airway epithelial cells. Further studies are required to test this hypothesis.
Quantitative assessment of intracorpuscular nerve density, expressed as integrated fluorescence density of SV2 immunofluorescence in NEBs at different stages of lung development.
It is noteworthy that the control of spontaneous tone in guinea pig tracheal preparations appears to be dependent on intact airway epithelium, including PNECs, (Skogvall et al. 1999). To further define the role of PNECs/NEBs, innervation studies using physiological methods are needed to complement histomorphological observations. Such studies are essential to confirm the postulated multifunctional role for PNECs/NEBs as hypoxia-sensitive chemoreceptors as well as locally active endocrine/paracrine cells responding to mechanical, chemical, or other stimuli.
Footnotes
Acknowledgements
Supported by funding from the Canadian Institutes of Health Research (CIHR) and Group Grant on Lung Development (EC and HY, MOP12742 and MGP15270).