
Abstract
The distribution of platelet endothelial cell adhesion molecule (PECAM-1, CD31) in vascular endothelium has been disputed. Originally reported to be highly concentrated at interendothelial cell contacts, recent studies have claimed that CD31 is distributed evenly over the entire endothelial cell surface. We re-investigated this question with two different murine anti-CD31 antibodies (MEC 13.3 and M-20), using a pre-embedding immunonanogold electron microscopic procedure that allowed precise label quantitation. MEC 13.3 reacted strongly with the luminal and abluminal plasma membranes of the endothelial cells lining microvessels in normal tissues and in angiogenic vessels induced by a tumor and vascular endothelial growth factor (VEGF-A164). Lateral plasma membranes were significantly less labeled. Conversely, M-20 strongly labeled the cytoplasmic face of the lateral plasma membranes of endothelial cells, although sparing specialized junctions, and only weakly labeled the luminal and abluminal plasma membranes. Both antibodies stained a significant minority of vesicles and vacuoles comprising the vesiculovacuolar organelle (VVO). Neither antibody was reactive in CD31-null mice. We conclude that CD31 is distributed over the entire endothelial cell surface, exclusive of specialized junctions, and in VVOs, but is not equally accessible to different antibodies in all locations.
Platelet endothelial cell adhesion molecule (PECAM-1, CD31) is a ∼130 kD glycoprotein that is constitutively expressed by endothelial cells and by platelets, neutrophils, monocytes, and some T-lymphocytes (Muller et al. 1989,1993; Vaporciyan et al. 1993; Carlos and Harlan 1994; Newman 1994,1997; Muller 1995a; Muller and Randolph 1999; Liao et al. 1999; Newton-Nash and Newman 1999; Henshall et al. 2001; Ilan et al. 2001; Newman et al. 2001; Jackson 2003; Cicmil et al. 2002; Newman and Newman 2003). It is an integral transmembrane protein that belongs to the immunoglobulin superfamily and that mediates both homotypic and heterotypic cell adhesion. These functions have implicated CD31 in a variety of important biological processes including inflammation, angiogenesis and wound healing (Schimmenti et al. 1992; Muller et al. 1993; Muller 1995a; Murohara et al. 1996; DeLisser et al. 1997; Newman 1997; Zhou et al. 1999; Cao et al. 2002; Jackson 2003; Newman and Newman 2003). In particular, CD31 has an important role in neutrophil and monocyte emigration across vascular endothelium, processes that can be inhibited, both in vitro and in vivo, with anti-CD31 antibodies (Bogen et al. 1992; Muller et al. 1993; Liao et al. 1995; Rival et al. 1996; Zocchi et al. 1996; Christofidou-Solomidou et al. 1997; Newman 1997).
Immunohistochemical (IHC) studies demonstrated that CD31 is distributed diffusely on the surfaces of inflammatory cells and platelets (Ohto et al. 1985; Metzelaar et al. 1991; Bogen et al. 1992; Newman et al. 1992). However, initial light microscopic studies of cultured endothelial cells in vitro and of the vasculature in vivo reported that CD31 was concentrated at the lateral borders of endothelial cells, i.e., at sites of interendothelial cell contact (Muller et al. 1989,1993; Albelda et al. 1990; Metzelaar et al. 1991; Vaporciyan et al. 1993; Ayalon et al. 1994; Carlos and Harlan 1994; Newman 1994,1997; Zocchi et al. 1996). These findings were believed to reflect homotypic binding in which CD31 joined together adjacent endothelial cells. They were also consistent with studies demonstrating that anti-CD31 antibodies inhibited leukocyte extravasation, a process that was believed to involve the passage of inflammatory cells through interendothelial cell junctions (Muller et al. 1989,1993; Muller 1995a,b; Muller and Randolph 1999).
Not all studies have agreed that CD31 is concentrated on the lateral membranes of endothelial cells. Some reports have found CD31 to be distributed diffusely on the surfaces of vascular and lymphatic endothelium (Ioffreda et al. 1993; Grafe et al. 1994; Erhard et al. 1996; Scholz and Schaper 1997; Lossinsky and Wisniewski 1998; Sauter et al. 1998; Marszalek et al. 2000; Ebata et al. 2001). Moreover, it has been claimed that CD31 is normally distributed widely over the surface of vascular endothelium in vivo but that, in response to TNF-α or other types of activation, CD31 is redistributed to the lateral plasma membranes (Ioffreda et al. 1993).
Our interest in CD31 was prompted by our studies of neutrophil emigration at sites of inflammation in guinea pig skin (Feng et al. 1998). Making use of serial electron microscopic sections and computer-assisted reconstructions, we demonstrated that, in response to an inflammatory stimulus, f-Met-Leu-Phe (FMLP), neutrophils extravasated by a transcellular, parajunctional pathway through greatly thinned endothelial cells, not by passing through interendothelial clefts. Interendothelial clefts were not widened and endothelial cell junctions remained tightly closed. Our findings therefore appeared to predict that anti-CD31 antibodies acted to inhibit neutrophil transmigration across endothelium in some fashion other than that of interfering with the opening of interendothelial junctions.
To further investigate this question, we investigated the ultrastructural distribution of CD31 in the microvasculature of a number of different tissues and organs in normal mice, in mouse tumor vessels, and in the new angiogenic vessels induced in flank skin by an adenovirus engineered to express vascular permeability factor/vascular endothelial growth factor (VPF/VEGF, VEGF-A164) (Pettersson et al. 2000). To our surprise, we found that two antibodies directed at different portions of the CD31 molecule gave distinctly different distributions in vascular endothelium, suggesting that these differences may be important for understanding the physiological and pathological functions of CD31.
Materials and Methods
Animals, Tumors, and Adenoviral Vector
We studied blood vessels in 4-6-week-old female mice, as follows: three normal A/Jax mice, two CD31-null mice (B6;129S-Pecamtm1Lex ), two B6;129S-F2/J mice that served as controls for the CD31-nulls (all obtained from the Jackson Laboratory; Bar Harbor, ME), and two athymic Nu/Nu mice (National Cancer Institute; Bethesda, MD). For studies of tumor blood vessels, 1 × 106 TA3/St mammary carcinoma cells were injected into the SC space of three syngeneic A/Jax mice and tumors were harvested 9 days later (Feng et al. 2000b). For additional studies of angiogenic blood vessels, an adenoviral vector engineered to express VEGF-A164 (Ad-VEGF-A164) was prepared as previously described (Pettersson et al. 2000). Plaque-forming units (108) were injected into the flank skin of two 4-6-week-old-female athymic Nu/Nu mice in a volume of 5 μl through a 30-g needle and tissue was harvested for study of new blood vessels 4 days later.
Antibodies to CD31
A rat monoclonal antibody (MAb) to CD31 (clone MEC 13.3, IgG2ak ) was obtained from Pharmingen (San Diego, CA). MEC 13.3 was prepared by immunizing with a polyoma middle T transformed endothelial cell line (Vecchi et al. 1994). It has been widely used to stain CD31 on vascular endothelium (Vecchi et al. 1994; Christofidou-Solomidou et al. 1997; Zhou et al. 1999), effectively blocks neutrophil accumulation in the inflamed mouse peritoneum and in TNF-α-stimulated skin grafts (Christofidou-Solomidou et al. 1997), and blocks FGF-induced angiogenesis (DeLisser et al. 1997). An affinity-purified goat polyclonal IgG antibody raised against a 20-amino-acid peptide mapping at the carboxy terminus of mouse CD31 (M-20) was obtained from Santa Cruz Biotechnology (Santa Cruz, CA). A goat polyclonal IgG directed against an epitope at the carboxy terminus of human VE-cadherin (C-19) was also obtained from Santa Cruz.
Tissue Fixation and Processing for Immunocytochemistry
Mice were sacrificed by CO2 narcosis and tissues were immediately fixed in 4% paraformaldehyde in 0.02 M PBS, pH 7.4, for 4 hr at room temperature (RT), washed in 0.02 M PBS, pH 7.4, and immersed overnight in 4C 30% sucrose in 0.02 M PBS, pH 7.4 (Feng et al. 2000a). Issues were then embedded in OCT compound (Miles; Elkhart, IN) and stored in liquid nitrogen.
Immunonanogold Silver Staining Protocol for Electron Microscopy
Tissue was processed for immunonanogold staining and electron microscopy as previously described (Feng et al. 2000a). In brief, 10-μm cryostat tissue sections were cut, collected on pre-cleaned glass slides, and air-dried for 20 min. All following steps were performed at RT: (a) wash in 0.02 M PBS, pH 7.4, 5 min; (b) immerse in 50 mM glycine in 0.02 M PBS, pH 7.4, 10 min; (c) wash in 0.02 M PBS, pH 7.4, 5 min; (d) immerse in 5% normal goat serum (NGS) (Vector Laboratories; Burlingame, CA), 20 min; (e) incubate in the primary antibody (MEC 13.3, M-20, or C-19) at dilutions of 1:25, 1:50, or 1:100 in 0.02 M PBS, 60 min; (f) three washes in 0.02 M PBS, pH 7.4, 5 min each; (g) incubate in the secondary antibody (affinity-purified goat anti-rat or rabbit anti-goat Fab1 conjugated to 1.4-nm nanogold) (Nanoprobes; Stony Brook, NY), 1:50 or 1:100 in 0.02 M PBS, pH 7.4, 60 min; (h) three washes in 0.02 M PBS, pH 7.4, 5 min each; (i) postfix in 1% glutaraldehyde in 0.02 M PBS, pH 7.4, 2 min (j) three washes in distilled water, 5 min each; (k) develop with HQ silver enhancement solution (Nanoprobes) for 6 min in the darkroom; (l) two washes in distilled water, 2 min each; (m) fix in 5% sodium thiosulfate, 1 min; (n) three washes in distilled water, 5 min each; (o) postfix in 1% osmium tetroxide in sym-collidine buffer, pH 7.4, 10 min; (p) one wash in 0.05 M sodium maleate buffer, pH 5.2, 5 min; (q) stain with 2% uranyl acetate in 0.05 M sodium maleate buffer, pH 6.0, 5 min; (r) one wash in distilled water, 5 min; (s) dehydrate in graded ethanols and infiltrate with a propylene oxide-eponate (Ted Pella; Redding, CA) sequence; (t) embed by inverting eponate-filled plastic capsules over the slide-attached tissue sections; (u) polymerize at 60C for 16 hr; (v) separate eponate blocks from glass slides by brief immersion in liquid nitrogen; (w) cut thin sections with a diamond knife with an ultratome (Reichert; Vienna, Austria), and collect sections on uncoated 200-mesh copper grids (Ted Pella); (x) view grids with a transmission electron microscope (CM-10; Philips, Einhoven, The Netherlands) without additional staining.
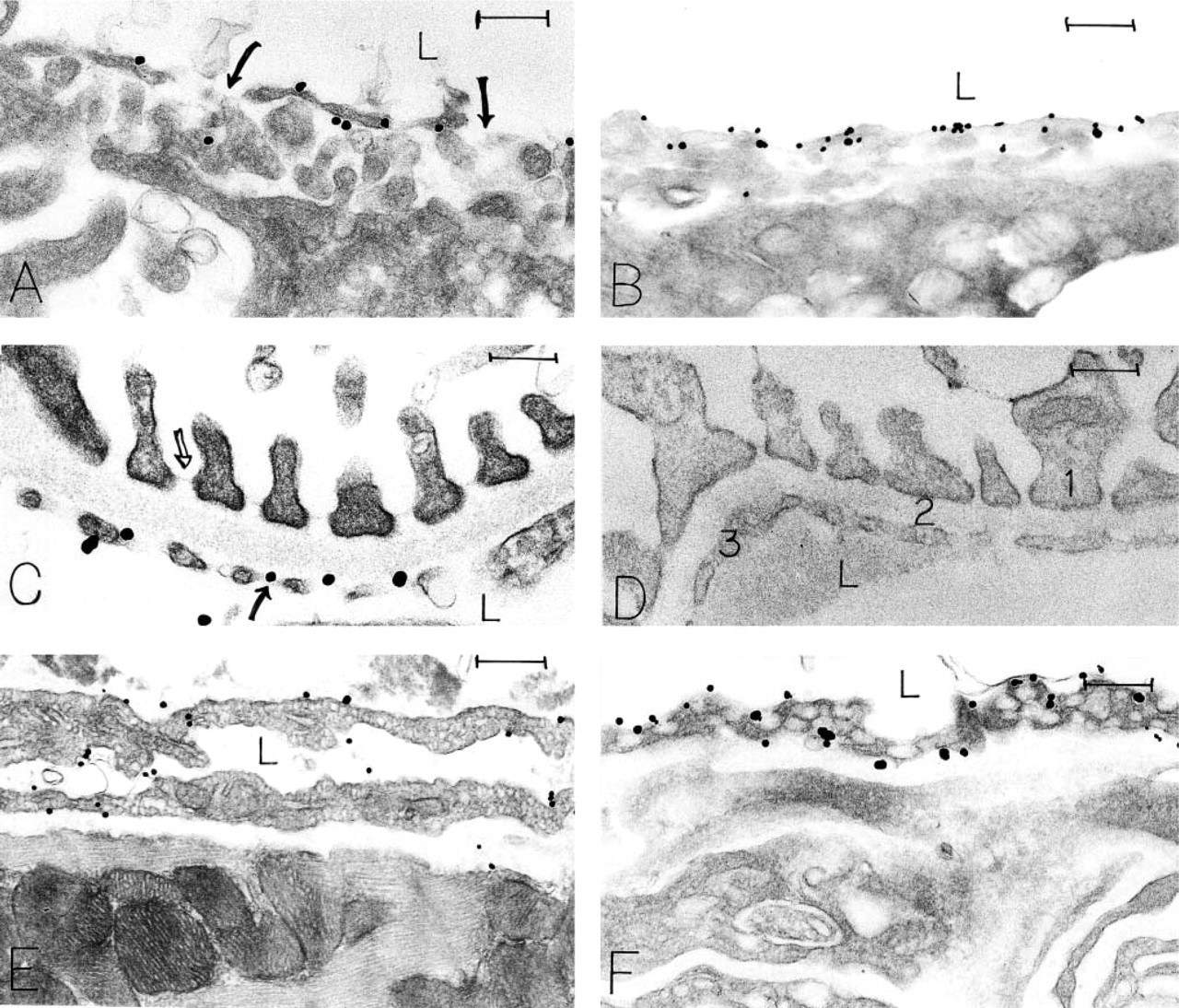
Localization of CD31 in mouse endothelial cells with a rat MAb (13.3) and an ultrastructural immunonanogold method. (
Controls for Immunonanogold Staining
Four controls were performed to ensure the specificity of staining: (a) replacement of the primary antibody by an irrelevant goat anti-rat or rabbit anti-goat antibody; (b) omission of specific primary antibody; (c) omission of the secondary antibody; and (d) omission of the HQ silver enhancement solution.
Quantitation of Immunonanogold Label
Silver-enhanced immunonanogold particles were counted on randomly selected electron micrographs of capillary and venule endothelial cells of normal mouse skin and heart, nude mouse lung, and hyperpermeable vessels induced by tumors or Ad-VEGF-A164. The subcellular localization of label on luminal, abluminal, and lateral plasma membranes was expressed as particles/linear micrometer of membrane or as percent of labeling of individual vesicles and vacuoles comprising VVOs. Statistical analysis was performed using the Mann-Whitney U-test.
Higher-magnification views localize CD31 to continuous, non-fenestrated endothelium in capillaries of normal A/Jax mouse heart (
Results
Ultrastructural Distribution of CD31 on Normal Microvessels with Rat MAb MEC 13.3
The ultrastructural distribution of CD31 was studied in the microvessels of normal organs that displayed a continuous (skin, heart, lung), fenestrated (kidney, adrenal), or discontinuous (liver) endothelium (Feng et al. 2002). MEC 13.3 gave a consistent staining pattern in all of these tissues, i.e., strong labeling of the luminal and abluminal plasma membranes, significantly less labeling of the lateral plasma membranes, and no staining of interendothelial cell tight or adherens junctions (Figures 1–3). These findings were most thoroughly documented in the continuous, non-fenestrated endothelium of skin, lung, and heart, where plasma membrane labeling was quantitated (p<0.001, Table 1).
CD31 localization to continuous, non-fenestrated endothelium in mouse skin with antibody 13.3. (
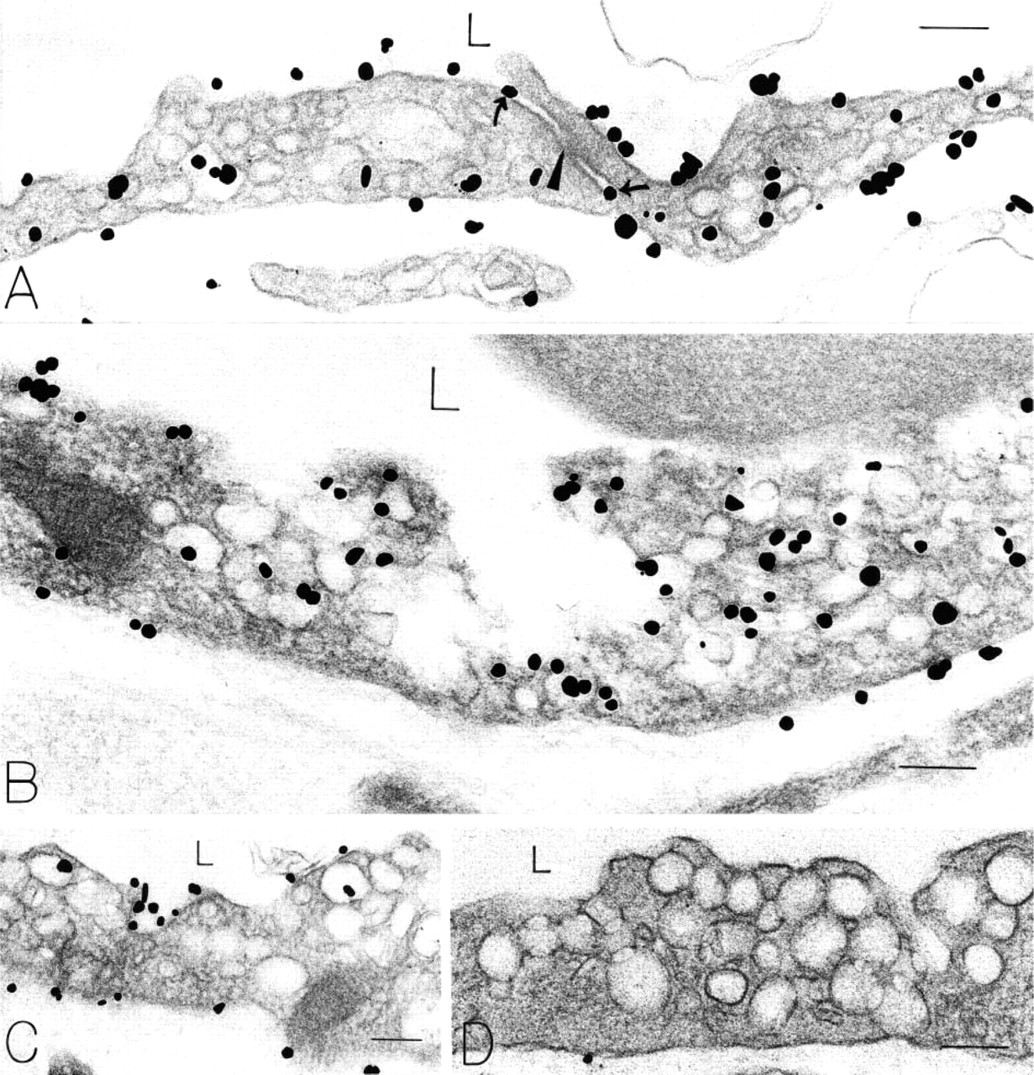
CD31 was also localized (Figure 3) to a significant fraction of the vesicles and vacuoles comprising vesiculovacuolar organelles (VVOs) (12-16% in several normal mouse tissues; Table 1). VVOs are clusters of uncoated, largely parajunctional vesicles and vacuoles found in venular endothelial cells. They are interconnected by stomata that are normally closed by thin diaphragms but that open in response to VEGF-A and other vasoactive agents to provide a transcellular pathway for plasma extravasation. Focal deep invaginations of the endothelial cell plasma membranes were also extensively labeled (Figure 3B). These structures may derive from VVOs (Feng et al. 1999). Other endothelial cell cytoplasmic organelles, such as mitochondria and multivesicular bodies, were not labeled. Parenchymal cells, pericytes, and basal laminae did not stain with this antibody, but platelets and neutrophils incidentally present in vascular lumens exhibited strong plasma membrane staining (not shown).
Monoclonal antibody MEC 13.3 immunonanogold quantitation of CD 31 in microvessel endothelial cell plasma membranes and VVOs of normal mouse skin, lung, and heart and in new blood vessels induced by VEGF-A164 or TA3/St tumor cells
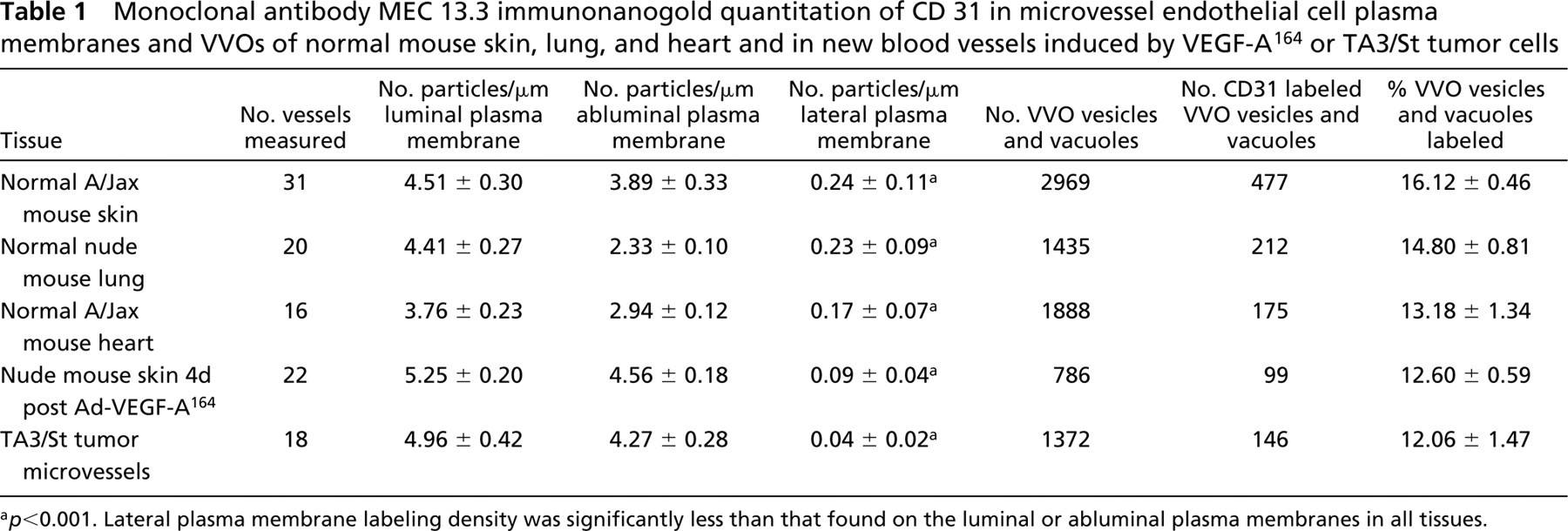
a p<0.001. Lateral plasma membrane labeling density was significantly less than that found on the luminal or abluminal plasma membranes in all tissues.
Ultrastructural Distribution of CD31 on Angiogenic Microvessels with Rat MAb MEC 13.3
The distribution and intensity of CD31 labeling in the new microvessels induced by either the TA3/St mammary adenocarcinoma (Figure 4) or by an adenoviral vector engineered to express VEGF-A164 (Figure 5) was similar to that observed in normal blood vessels (Table 1). The luminal and abluminal surfaces of vascular endothelium showed strong and roughly equivalent labeling, whereas the lateral surfaces had comparatively little labeling (p<0.001). Approximately 12% of VVO vesicles and vacuoles were labeled, comparable to that observed in normal microvessels. Specialized interendothelial cell junctions (adherens, tight) exhibited no staining (Figures 5A and 5C). Pericytes, basal laminae, tumor cells, and other tissue cells did not stain with this antibody. However, as in normal tissues, circulating monocytes and neutrophils did exhibit plasma membrane labeling as well as labeling of some internal cytoplasmic endocytic/secretory vesicles and granule membranes (not shown).
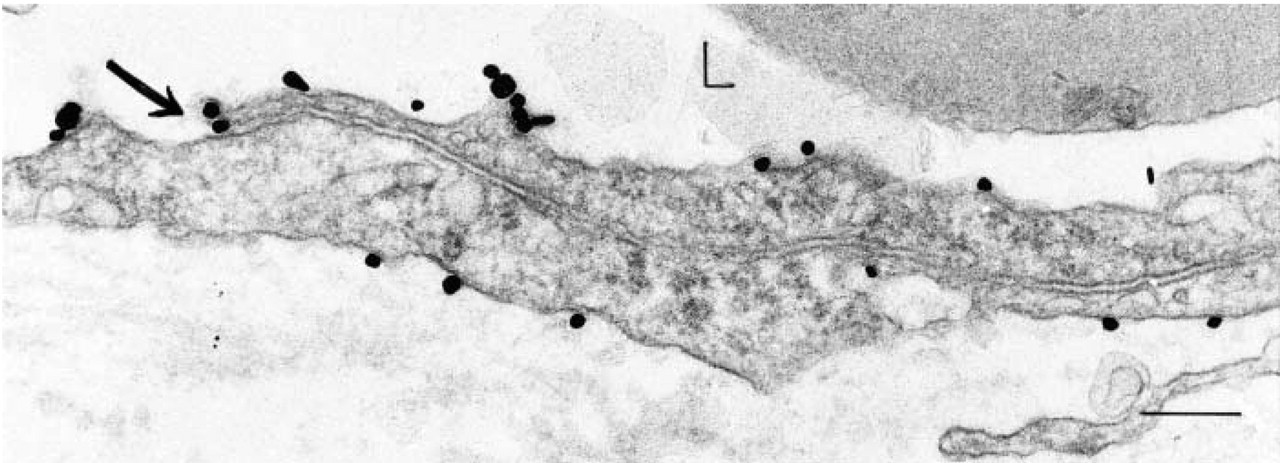
CD31 localization with antibody 13.3 in subcutaneous tumor-associated vessels 9 days after implantation of TA3/St mouse mammary tumor. Luminal, and to a lesser extent abluminal, plasma membrane is labeled, whereas the long segment of lateral plasma membranes that form the intercellular cleft between two endothelial cells is not. Two particles are bound to the luminal flap of one endothelial cell at the luminal introit of the lateral interendothelial cell cleft (arrow). L, lumen. Bar = 260 nm.
Ultrastructural Distribution of CD31 on Microvessels with a Goat Polyclonal Antibody Directed Against a CD31 Terminal Peptide (M-20)
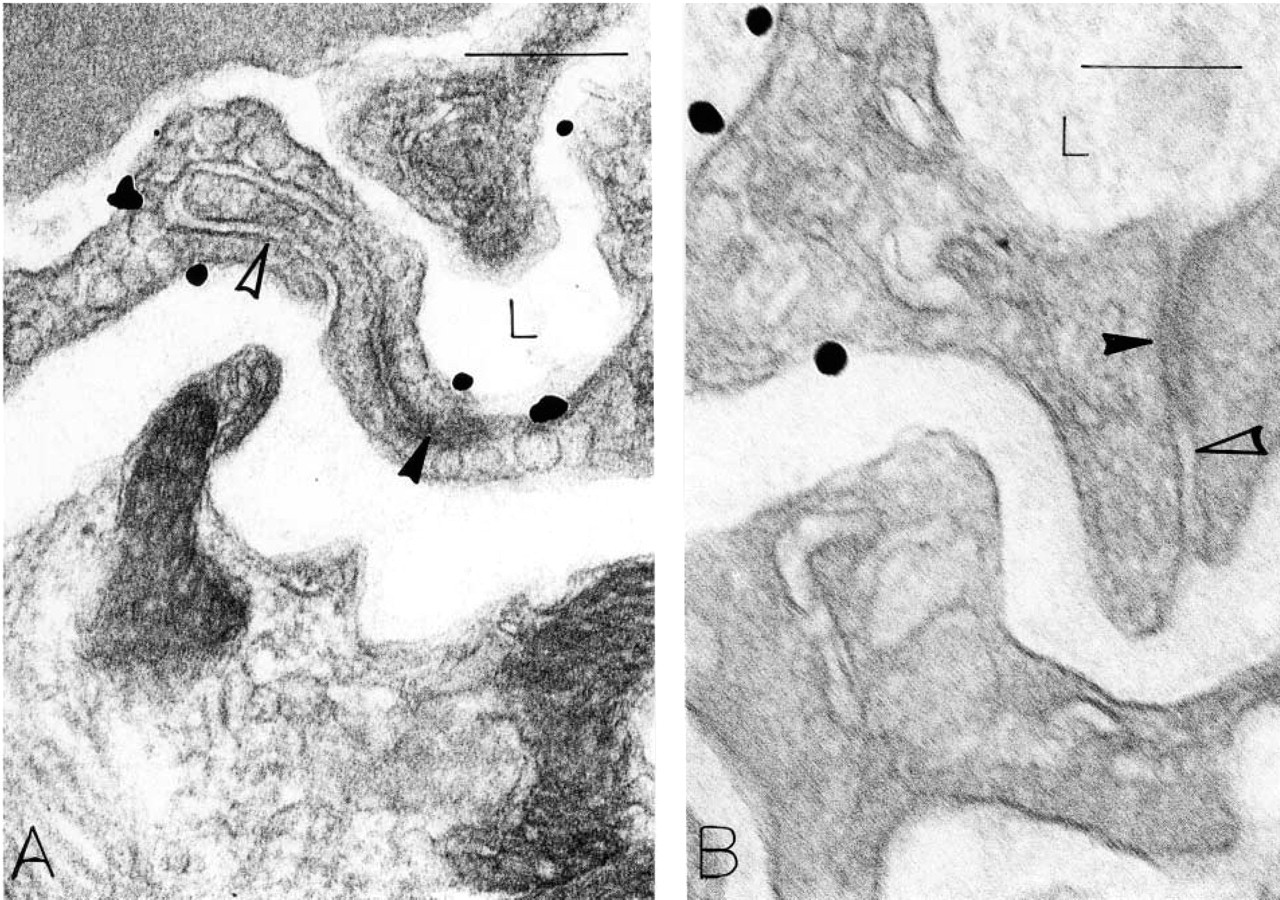
Antibody M-20 gave a labeling distribution that was distinctly different from that of MEC 13.3 ((Figures 6 and 7). The lateral plasma membranes of apposed microvascular endothelial cells were strongly and selectively labeled, both in normal tissues and in the new microvessels induced by Ad-VEGF-A164, whereas the luminal and abluminal plasma membranes were largely spared (Table 2). However, despite intense and selective labeling of the lateral endothelial membranes, tight junctions and adherens junctions were not labeled (Figures 6A-6D). Consistent with the C-terminal localization of the antigen, staining was localized to the inner face of the plasma membrane (Figures 6 and 7A). M-20 also stained a significant minority of VVO vesicles and vacuoles (17-23%; Table 2), predominantly on their cytoplasmic face (Figure 7B).
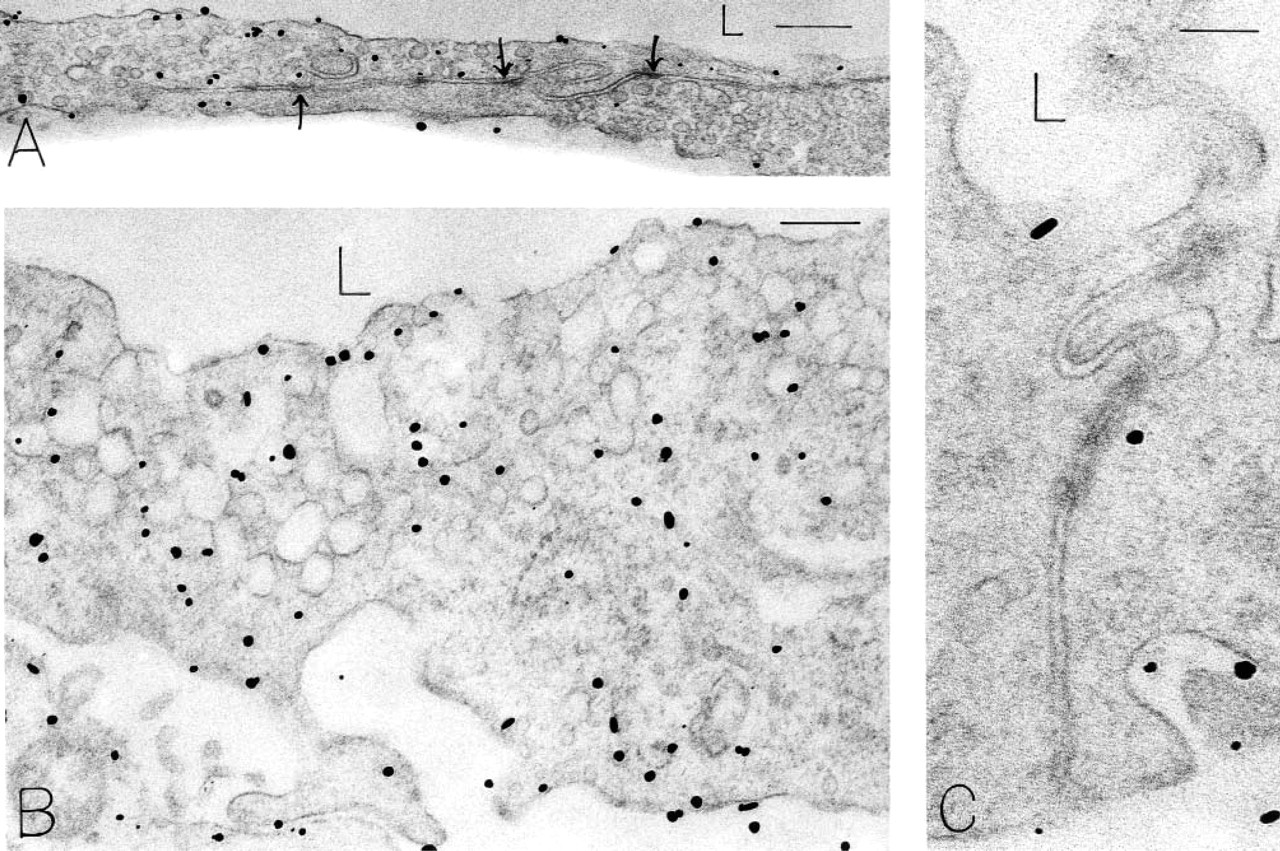
CD31 localization with antibody 13.3 in angiogenic vessels generated in nude mouse skin 4 days after injection of Ad-VEGF-A164. (
Studies in CD31-null Mice
We considered the possibility that the disparate staining pattern observed was attributable to a lack of specificity of one or both of our antibodies. Therefore, we repeated our immunocytochemical studies on the normal tissues (skin, kidney) of CD31-null and control mice. In contrast to strong staining in B6;129S-F2/J controls, neither antibody stained the microvessels of CD31-null B6;129S-Pecamtm1Lex mice (Figure 8).
Ultrastructural Distribution of VE-Cadherin in Microvascular Endothelium
New blood vessels induced by tumors or Ad-VEGF-A164 localized VE-cadherin to the cytoplasmic faces of adherens junctions (Figures 9A and 9C). Normal microvessels exhibited a similar pattern of staining, although somewhat less intense. Luminal and abluminal plasma membranes, VVOs, and coated vesicles were not stained (Figures 9A, 9C, and 9D). An irrelevant antibody control was negative (Figure 9B).
Specificity Controls for the Immunogold Procedure
All four controls (see Materials and Methods) were negative. Illustrated are omission of the primary anti-body (Figure 1D) and substitution of an irrelevant IgG for specific primary antibodies (Figures 3D and 9B). We also examined skin endothelial cells in CD31 knockout mice (Figures 8C and 8D), which gave no staining of plasma membranes or of VVOs with either MEC 13.3 or M-20.
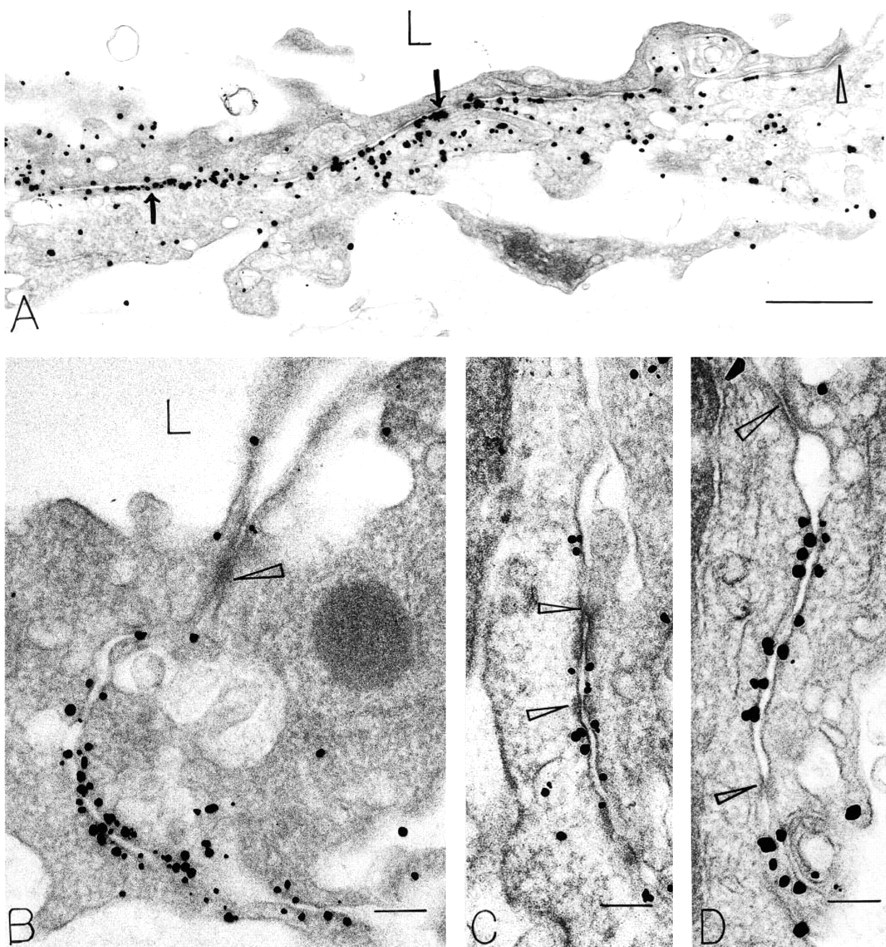
Immunonanogold staining of endothelial cells in Ad-VEGF-A164 expressing (
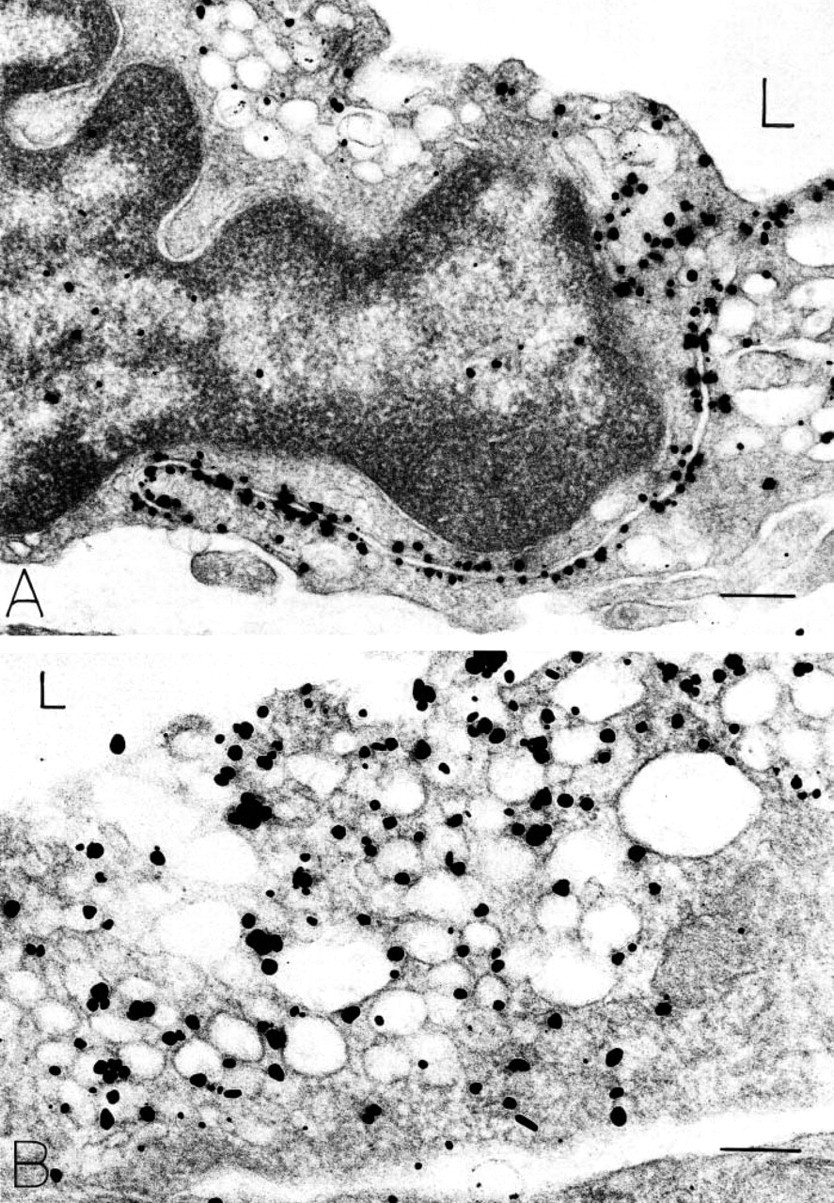
Immunonanogold staining of endothelial cells in normal mouse skin with antibody M-20. (
Discussion
The distribution of CD31 on vascular endothelium has been a matter of dispute. Although distributed evenly over the surface of non-confluent cultured endothelial cells, CD31 was reported to become highly concentrated at interendothelial cell contacts both in vivo and in vitro (Muller et al. 1989; Albelda et al. 1991), and this view has been widely accepted in the literature (see Scholz and Schaper 1997 for complete references). On the other hand, Grafe et al. (1994), Scholz and Schaper (1997), and others (Erhard et al. 1996; Sauter et al. 1998) have reported that CD31 was homogeneously distributed over the entire endothelial cell surface, luminal and abluminal as well as lateral, both in vitro and in vivo. Scholz and Schaper (1997) used confocal and immunocytochemical electron microscopy to study five different mouse MAbs and a rabbit polyclonal antibody directed against human CD31. Some of the antibodies used in these studies had been provided by authors who had previously used them to localize CD31 to the cell-cell interface. Scholz and Schaper (1997) noted that most of the earlier studies that claimed a junctional localization had been performed on cultured endothelium using light microscopic IHC or immunofluorescence and may not have fully taken into account anatomic features that would artifactually accentuate staining at intercellular contacts (e.g., overlapping membranes of apposed endothelial cells could give the appearance of increased interface staining). However, none of the investigators to date made any attempt to quantitate CD31 epitope density in different regions of the endothelial cell plasma membrane.
Polyclonal goat antibody M-20 immunonanogold quantitation of CD31 in microvessel endothelial cell plasma membranes and VVOs in normal mouse skin and in the new blood vessels induced by VEGF-A164
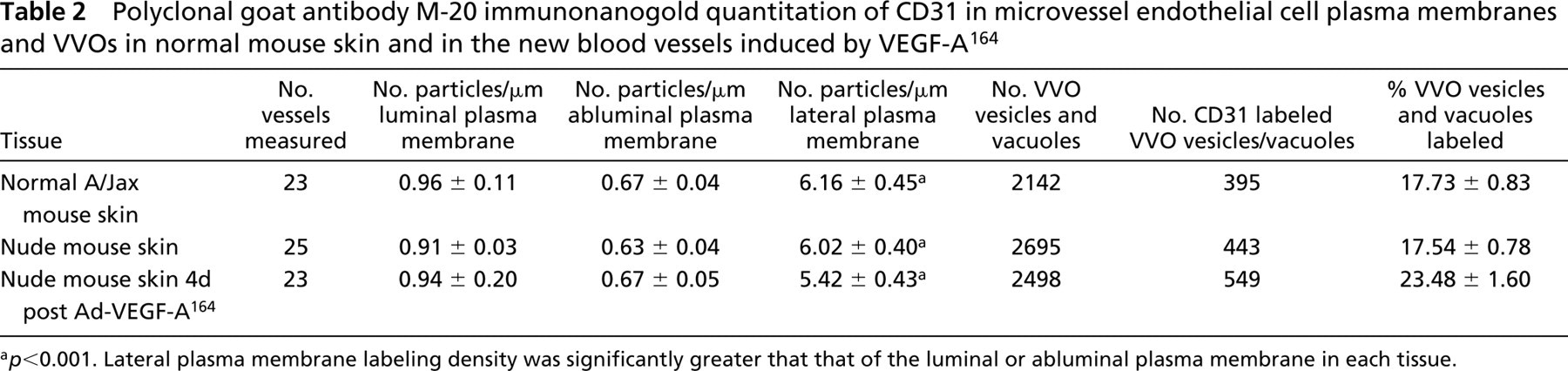
a p<0.001. Lateral plasma membrane labeling density was significantly greater that that of the luminal or abluminal plasma membrane in each tissue.
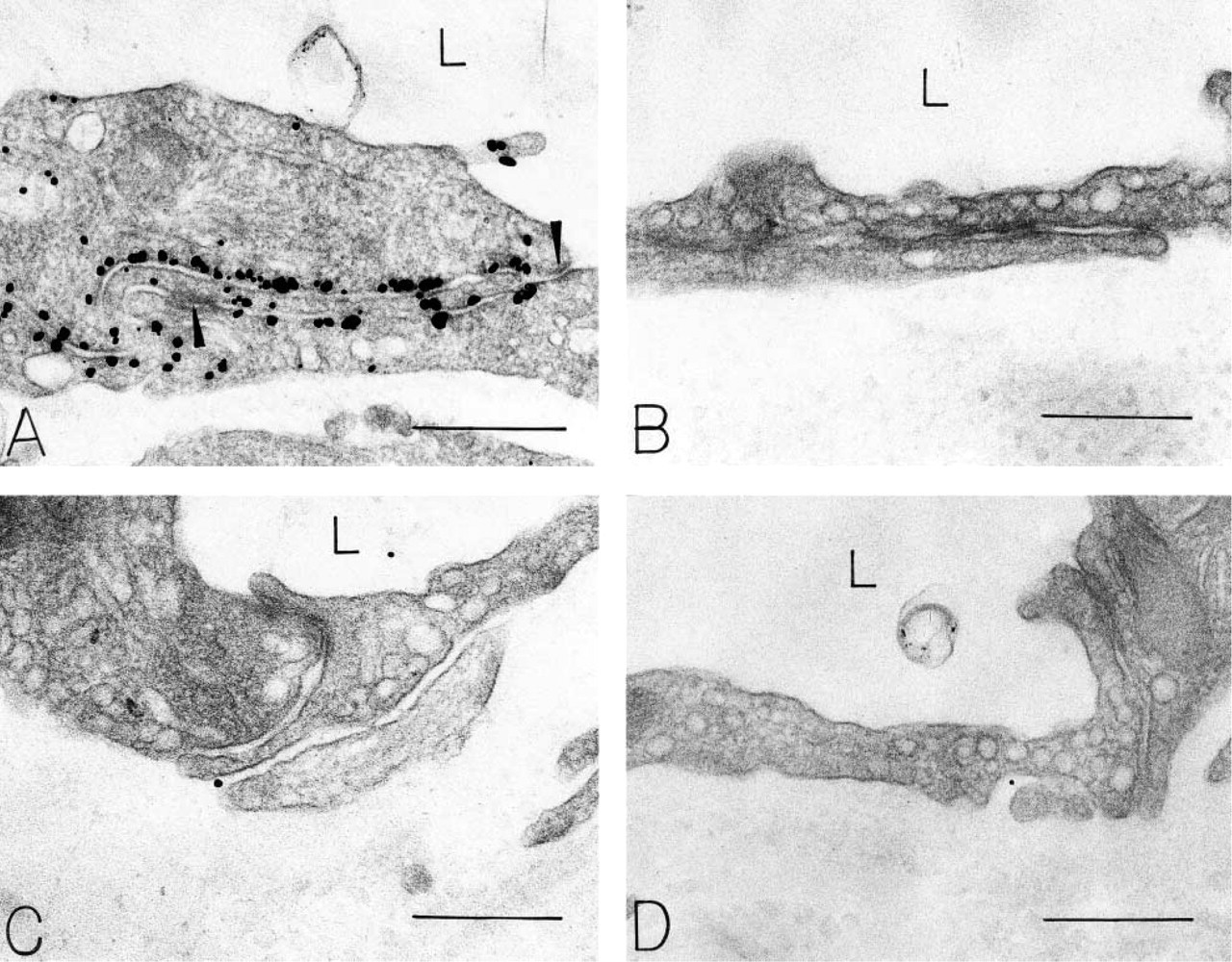
Immunonanogold preparations of skin endothelium of wild-type (
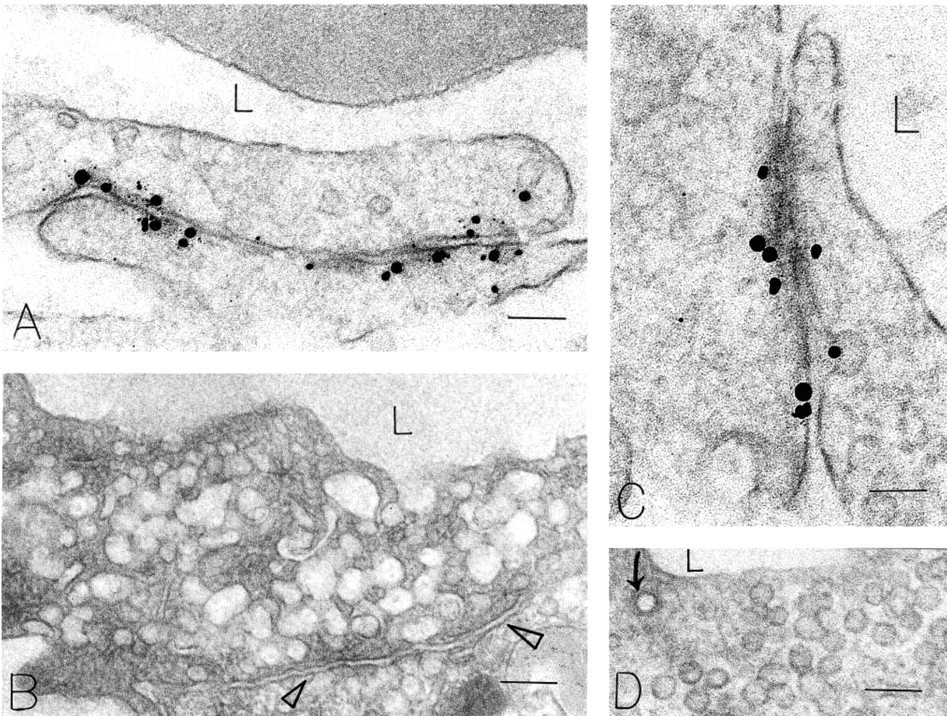
VE-cadherin immunonanogold localization in microvessels from Ad-VEGF-A164 injected mouse skin at 5 days (
In an attempt to clarify the issue of CD31 distribution on vascular endothelium, we used a pre-embedding immunonanogold electron microscopic procedure that allowed precise subcellular localization and quantitation of CD31 epitope distribution (Feng et al. 2000a). Using two different antibodies to mouse CD31 that have not been previously investigated at the electron microscopic level, we found strikingly different labeling patterns in mouse microvascular endothelium. A much studied blocking antibody, MEC 13.3, reacted strongly with the extracellular face of the luminal and abluminal plasma membranes of microvascular endothelial cells but displayed only minor labeling of the lateral plasma membranes (Figures 1–5; Table 1). This labeling pattern was similar in several normal tissues, in TA3/St tumor vessels, and in the angiogenic vessels induced by Ad-VEGF-A164. In contrast, a polyclonal antibody, M-20, directed against a C-terminal peptide of CD31, strongly labeled the cytoplasmic face of the lateral plasma membranes of endothelial cells, although sparing tight and adherens junctions. The luminal and abluminal membranes were labeled to a much lesser extent (Figures 6 and 7; Table 2). Given the significantly different distributions of CD31 observed with these two antibodies (Figure 10), we considered the possibility that one or both were not specific for CD31. However, neither antibody reacted with vascular endothelium (or leukocytes or platelets) in CD31-null mice (Figure 8), providing strong evidence that both were indeed specific for CD31.
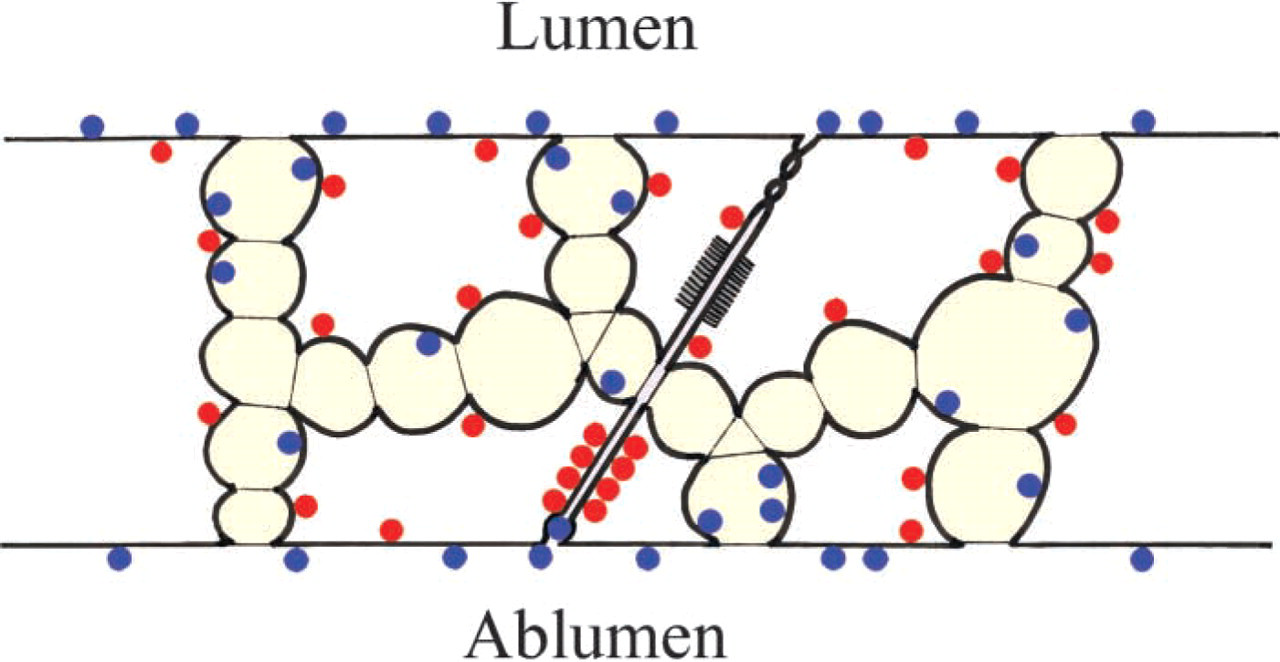
Schematic diagram illustrating labeling pattern of MEC 13.3 (blue dots) and M-20 (red dots) in vascular endothelium. MEC 13.3 primarily labels the external surfaces of luminal and abluminal surface membranes, whereas M-20 primarily labels the cytoplasmic face of lateral plasma membranes. Both antibodies label a fraction of VVO vesicles and vacuoles. M13.3 label was found predominantly on the inner vesicular/vacuolar face, whereas M-20 staining was predominantly on the cytoplasmic face, Neither labeled specialized adherens or tight junctions.
We conclude that CD31 is distributed over the entire endothelial cell surface, luminal, abluminal, and lateral, with the exception of specialized intercellular junctions, as also previously reported (Lampugnani et al. 1992; Ayalon et al. 1994). Failure to stain specialized junctions with either antibody cannot be attributed to poor antibody penetration because adherens junctions were strongly and selectively stained with an antibody to VE-cadherin (Figure 9).
We do not have a clear explanation for the striking quantitative differences in CD31 distribution exhibited by the two antibodies we studied, although they probably relate to their recognition of different CD31 epitopes. The polyclonal M-20 antibody recognizes a cytoplasmic epitope of CD31. Therefore, one possibility is that the cytoplasmic portion of CD31 molecules distributed in the luminal and abluminal plasma membranes form complexes with other cytoplasmic proteins that make them inaccessible to the M-20 antibody. Another possible explanation relates to the fact that the cytoplasmic portion of CD31 has multiple splicing variants (Sheibani et al. 1999; Jackson 2003; Newman and Newman 2003). It is not known whether these different splicing variants are distributed differently in different sites in the vascular endothelium. If so, it is possible that the C-terminal epitope of CD31 that is recognized by the M-20 antibody is preferentially expressed by CD31 molecules localized in the lateral plasma membrane. New antibodies directed against different CD31 cytoplasmic epitopes will be needed to address this question.
Alternative splicing might not have been expected to prevent MEC 13.3 from labeling the lateral plasma membranes more strongly, because splice variants described thus far have been confined to the cytoplasmic portion of CD31. However, different splicing variants of the cytoplasmic tail of CD31 have been shown to affect CD31 binding (Baldwin et al. 1994; DeLisser et al. 1994; Yan et al. 1995; Famiglietti et al. 1997). A more likely explanation for the failure of MEC 13.3 to label lateral endothelial cell plasma membranes is that MEC 13.3 reacts with extracellular epitopes of CD31 that are engaged in homotypic bonds between adjacent endothelial cells and therefore are unavailable for binding to antibody.
In addition to labeling the plasma membrane of endothelial cells, both MEC 13.3 and M-20 antibodies labeled a significant fraction (12-20%) of VVO vesicles and vacuoles. VVOs represent an important pathway for transcellular passage of macromolecules and fluid in response to vascular permeabilizing mediators such as VEGF-A (Dvorak et al. 1996; Feng et al. 1996,1999). They may also have a role in neutrophil diapedesis (Feng et al. 1998) and may provide a substantial store of membrane that can be rapidly mobilized to the cell surface early in angiogenesis to allow the rapid expansion in cell surface area that accompanies vessel enlargement (formation of “mother” vessels) (Pettersson et al. 2000; Nagy et al. 2002). CD31 has been reported to have a role in both vascular permeability and angiogenesis (Zhou et al. 1999; Cao et al. 2002; Graesser et al. 2002), and CD31 represented on VVOs could be involved. Finally, because VVOs are often parajunctional, their staining with antibodies to CD31 may also have contributed to the idea that CD31 is preferentially located at the interface between adjacent endothelial cells. By light microscopy, it may not have been possible to distinguish IHC reaction product located in VVOs from that localized to the lateral plasma membrane of endothelial cells.
It remains unclear exactly how antibodies to CD31 inhibit leukocyte migration. A recent report (O'Brien et al. 2003) describes five mechanisms by which CD31 could modulate the neutrophil migration across endothelial cell monolayers that is induced by cytokines (e.g., IL-1β). The authors also note that transendothelial migration induced by neutrophil chemoattractants (LTB4) or chemokines (IL-8) is CD31-independent. Blocking antibodies are reported to have limited access to CD31 molecules at the interface between adjacent endothelial cells (Muller et al. 1993). Therefore, such antibodies may act in other ways, binding to CD31 expressed on leukocytes, on the luminal surface of endothelial cells, or on VVOs. The Muller laboratory has provided strong evidence that antibodies to CD31 that inhibit transmigration do not prevent adherence of leukocytes to the apical surface of vascular endothelium (Muller et al. 1993; Liao et al. 1995). Therefore, attachment of leukocytes to endothelium is not dependent on CD31, whereas transmigration of leukocytes is, at least for certain stimuli. Blocking antibodies could also act by interfering with cell signaling mechanisms (Newton-Nash and Newman 1999; Henshall et al. 2001; Ilan et al. 2001; Newman et al. 2001; O'Brien et al. 2003).
Our finding of CD31 labeling of VVO vesicles and vacuoles relates to a recent paper that described a population of CD31-positive vesicles located just beneath the plasmalemma at the borders of cultured human endothelial cells (Mamdouh et al. 2003). These vesicles were found to recycle evenly along endothelial cell borders but, during leukocyte emigration, were targeted to segments of the border across which monocytes were migrating. They were believed to facilitate monocyte emigration because blocking antibodies to CD31 specifically inhibited the recruitment of these vesicles to zones of leukocyte migration. The authors concluded that their vesicles were not caveolae or associated with VVOs because CD31 and caveolin staining did not co-localize. However, VVO vesicles and vacuoles are commonly parajunctional, and individual VVO vesicles not uncommonly fuse to the lateral plasmalemma (see Figure 1 in Feng et al. 1997). It therefore remains to be determined whether the vesicles described by Mamdouh et al. (2003) represent an entirely new population of vesicles or one that is related to VVOs.
In summary, we have shown that CD31 is localized to the entire surface of vascular endothelial cells in both normal tissues and in tumor and Ad-VEGF-A164-induced angiogenesis. However, two different authenticated antibodies gave significantly different distributions on the endothelial cell surface (Figure 10). In addition, both antibodies reacted with a significant fraction of VVO vesicles and vacuoles. Further work will be required to elucidate the reasons for these differences and to work out the role played by CD31 in inflammatory cell diapedesis.
Footnotes
Acknowledgements
Supported in part by US Public Health Service grants CA-50453, HL-59316, and P01 CA92644 (HFD), AI-33372 and AI-44066 (AMD), and by a contract from the National Foundation for Cancer Research (HFD).