
Abstract
The use of 10–15-nm gold probes in freeze-fracture immunocytochemistry sometimes results in poor immunogold labeling. Replica sites are labeled with only one or two gold particles, making it unlikely that the labeling depicts the true distribution of antigen. In this study, the feasibility of using ultrasmall (∼1.4-nm) gold probes for immunocytochemical labeling of replicas was examined. When HLA Class I in neutrophil membrane replicas was labeled with various sized immunogold particles as the secondary detection system, the apparent distribution density was inversely related to the size of the particles (1.4-nm <5-nm <10-nm <15-nm). Indeed, the density of the apparent distribution of HLA Class I labeled with 1.4-nm gold particles was about sevenfold greater than when labeling was carried out with the 10-nm gold particles. Similar results were obtained with CD16, another neutrophil membrane protein. Silver enhancement was required to visualize the 1.4-nm gold particles, but this procedure did not adversely affect replica membranes. These results suggest that, when followed by silver enhancement, 1.4-nm gold particles are effective probes for achieving high-resolution immunocytochemical labeling of replicas.
Keywords
F
Colloidal gold particles of 10–15-nm diameter have been routinely used as a secondary detection system in freeze-fracture immunocytochemistry, because smaller particles (e.g., 5-nm) can not be readily distinguished on the relatively electron-dense, granular replica membranes stabilized by platinum/carbon (Pt/C) evaporation. We previously used this method to visualize the distributions of selected molecules in human neutrophil membranes and observed that immunogold labeling was sometimes poor when larger colloidal gold probes were used. This made it difficult to distinguish whether labeled sites on replicas represented the true distribution of antigen. This article reports on the use of 1.4-nm gold immunoprobes in conjunction with silver enhancement. The results presented here show that, when followed by silver enhancement, 1.4-nm immunogold particles serve very well as immunogold probes and achieve intense labeling of antigens on replicas.
Materials and Methods
Labeling of Human Neutrophil Replicas with Immunogold Probes
Replicas of human neutrophils were prepared as previously described (Takizawa et al. 1998); reagents were the same except as noted. Briefly, unstimulated human neutrophils were rapidly frozen, freeze-fractured at −130C, and subsequently replicated by Pt-Pd/C or Pt/C evaporation. The replicas were then digested in 10–20 mM octyl glycoside (OG)-PBS solution for 1 hr at 4C. After OG digestion, the replicas were incubated in PBS containing 1% bovine serum albumin (BSA) or 10% normal goat serum to block nonspecific protein binding sites and then incubated with the following antibodies for 120 min at 22C: mouse monoclonal anti-HLA Class I (clone W6/32; Dako, Glostrup, Denmark) and anti-CD16 (clone DJ130c; Dako). Detection of bound primary antibody was accomplished by incubating the replicas for 90 min in goat anti-mouse Nanogold (NanoProbes, Stony Brook, NY; 1:40 dilution). This ultrasmall immunogold probe consists of a 1.4-nm gold particle covalently conjugated to an affinity-purified goat anti-mouse Fab' fragment. After immunocytochemical labeling with Nanogold, the replicas were postdigested overnight in 20 mM OG-PBS at 22C, washed five times in PBS, and fixed for 10 min at 22C in 2% glutaraldehyde-PBS. The samples were then washed five times with distilled water, mounted on formvar-coated copper or nickel grids with the Pt-Pd/C cast in contact with the thin formvar film (i.e., the immunostained membrane halves were exposed), and then dried. The 1.4-nm gold particles were then silver-enhanced to facilitate visualization by electron microscopy (Burry 1995). Controls consisted of the primary antibody substituted with nonimmune IgG of the same isotype; IgG isotype control was used at the same concentration as the primary antibody.
For purposes of comparison, goat anti-mouse 5-, 10-, and 15-nm colloidal gold particles (NanoProbes) were also used as secondary antibodies; 5-nm colloidal gold particles were silver-enhanced. In addition, goat anti-mouse 10-nm colloidal gold particles prepared by the method of DeMey (1984) were applied.
Morphometric Analysis of Immunogold in Human Neutrophils
For analysis of the apparent distributions of HLA Class I, the labeling density of various-sized immunogold particles (1.4-, 5-, 10-, and 15-nm diameter) was determined. Negatives of electron micrographs of the replicas were printed at the same magnification (X 61,500); the areas of the exoplasmic halves (E-faces) of plasma membranes were then measured, and the number of individual immunogold particles on the E-faces was counted.
Results
Initially, we characterized the distribution of HLA Class I on replicas of unstimulated human neutrophils using 1.4-nm immunogold particles as the secondary detection system (Figure 1). When silver-enhanced, the E-faces of the plasma membranes were heavily labeled with the ultrasmall gold particles. Labeling of HLA Class I was readily apparent even at low magnification (Figure 1A). Few gold particles were present on nuclear membranes (Figure 1B). In the substitution of the primary antibody with nonimmune IgG, there was no immunocytochemical labeling despite application of 1.4-nm immunogold and subsequent silver enhancement (Figure 1C). The replica membranes, which consisted of split membrane halves and their Pt-Pd/C casts, were unaffected by the silver enhancement procedure (Figure 1).
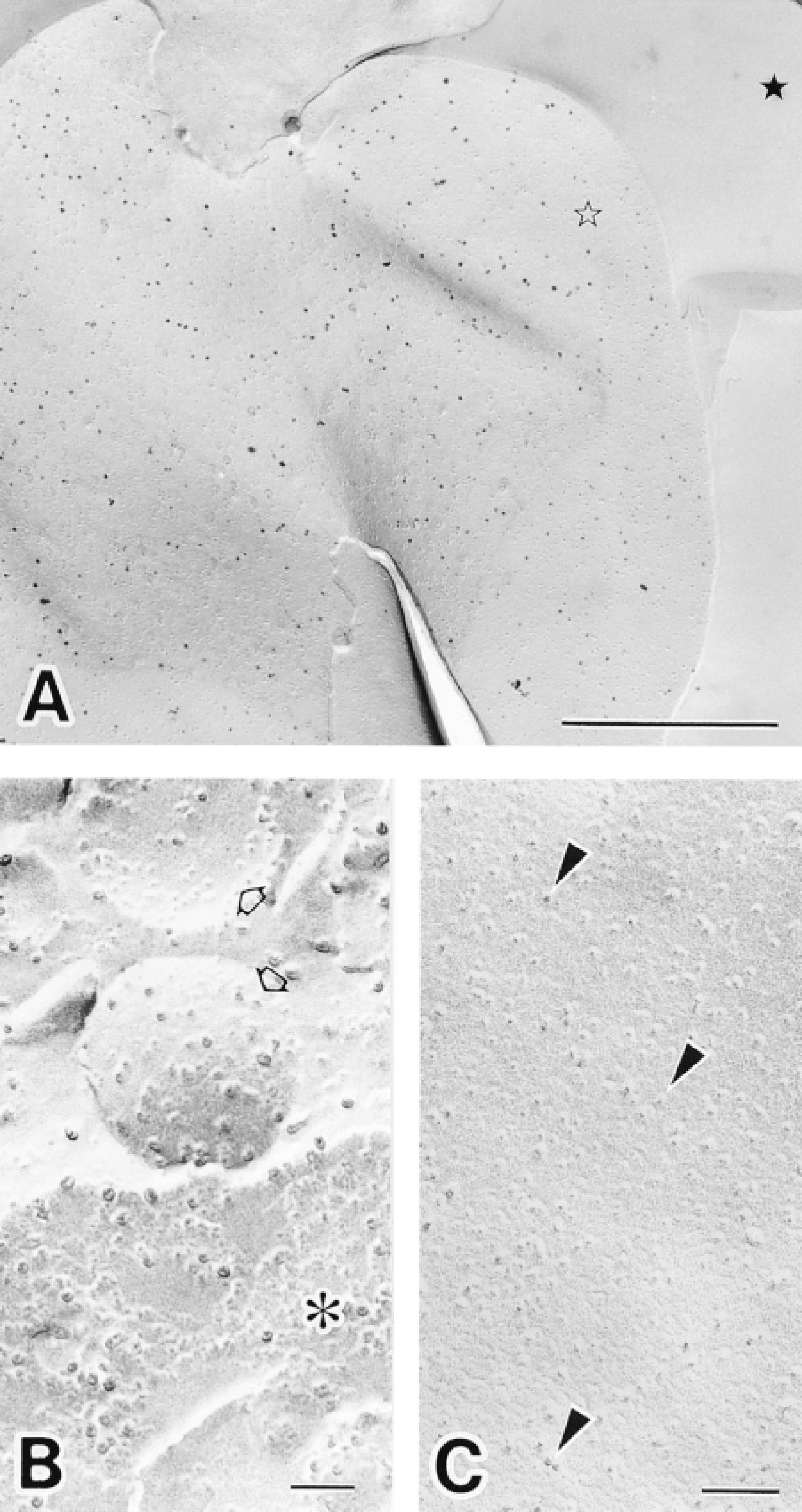
Immunocytochemical labeling of HLA Class I in human neutrophils replicas achieved using 1.4-nm gold-conjugated secondary antibody and subsequent silver enhancement. (A) Low-magnification electron micrograph of a plasma membrane replica showing the distribution of HLA Class I. Silver-enhanced 1.4-nm gold particles are present on the exoplasmic half (E-face) of the plasma membrane (open star). Freeze-fractured extracellular space (filled star) is evident. Bar = 1 μm. (B) Electron micrograph of a nuclear membrane after the immunocytochemical labeling of HLA Class I (∗); few silver-enhanced gold particles are present. Intracellular granules are evident (open arrows). (C) Control experiment in which the primary antibody was substituted with nonimmune IgG; note the absence of silver-enhanced gold labeling on the E-face of a plasma membrane. Intramembranous particles (arrowheads) are evident. Bars = 0.1 μm. Note that the replica membranes are unaffected by silver enhancement in A-C.
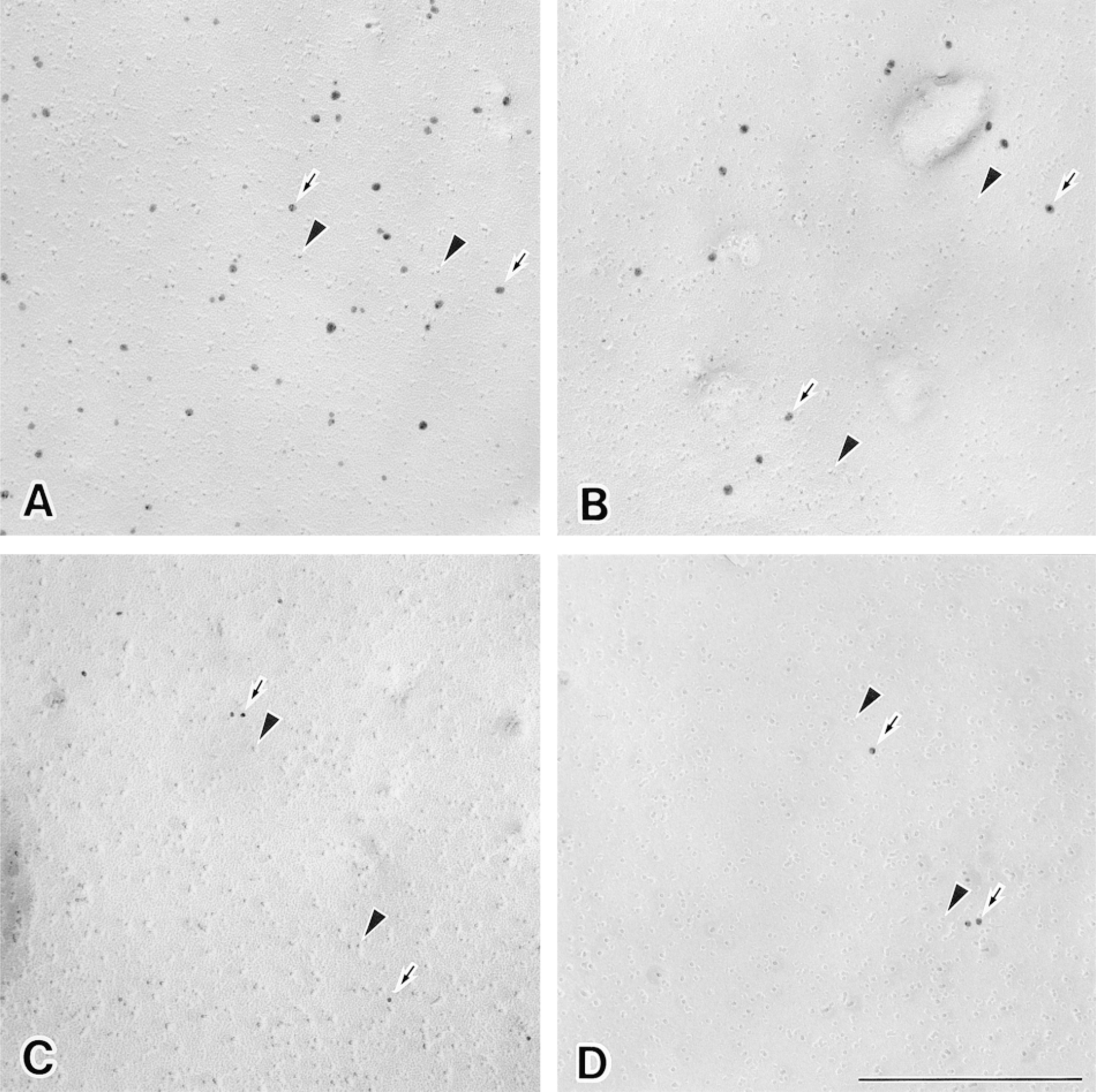
Labeling of HLA Class I on replicas of freeze-fractured plasma membranes (E-faces) using various sizes of immunogold particles. Replicas were incubated with mouse anti-HLA class I under identical conditions and then incubated with the different immunogold particles under the same conditions. Gold of 1.4 and 5 nm was subsequently silver-enhanced. (A) The replica exhibiting the distribution of HLA Class I is labeled with 1.4-nm gold. (B) The replica is labeled with 5-nm gold. (C) The replica is labeled with 10-nm gold. (D) The replica is labeled with 15-nm gold. Immunogold particles (arrows) and intramembranous particles (arrowheads) are evident. Bar = 0.1 μm.
Neutrophil replicas labeled with different-sized immunogold particles (1.4-, 5-, 10-, and 15-nm in diameter) exhibited striking differences in labeling density. As judged by the intensity of the immunogold labeling, the apparent distribution of HLA Class I on the E-faces of plasma membranes was inversely related to the size of the immunogold probe used as the secondary detection system (Figures 2 and 3). Therefore, the distribution of 1.4-nm gold particles was about seven times as dense as that of the 10-nm gold particles routinely used for freeze-fracture immunocytochemistry (Figure 3).
CD16 was also visualized with immunogold probes as the secondary detection system and was found to be located on both plasma membranes (data not shown) and small intracellular granules (Figure 4) in unstimulated neutrophils. CD16 on the E-faces of granules was somewhat difficult to detect when 10-nm gold probes were used (Figure 4B). This was probably due to combined effects of steric hindrance between the comparatively large colloidal gold particles and the relatively low abundance of protein in the granules. In contrast, CD16-containing granules were readily recognized using the ultrasmall gold technique. The E-faces of the granules were heavily labeled with the silver-enhanced 1.4-nm immunogold probes (Figure 4A).
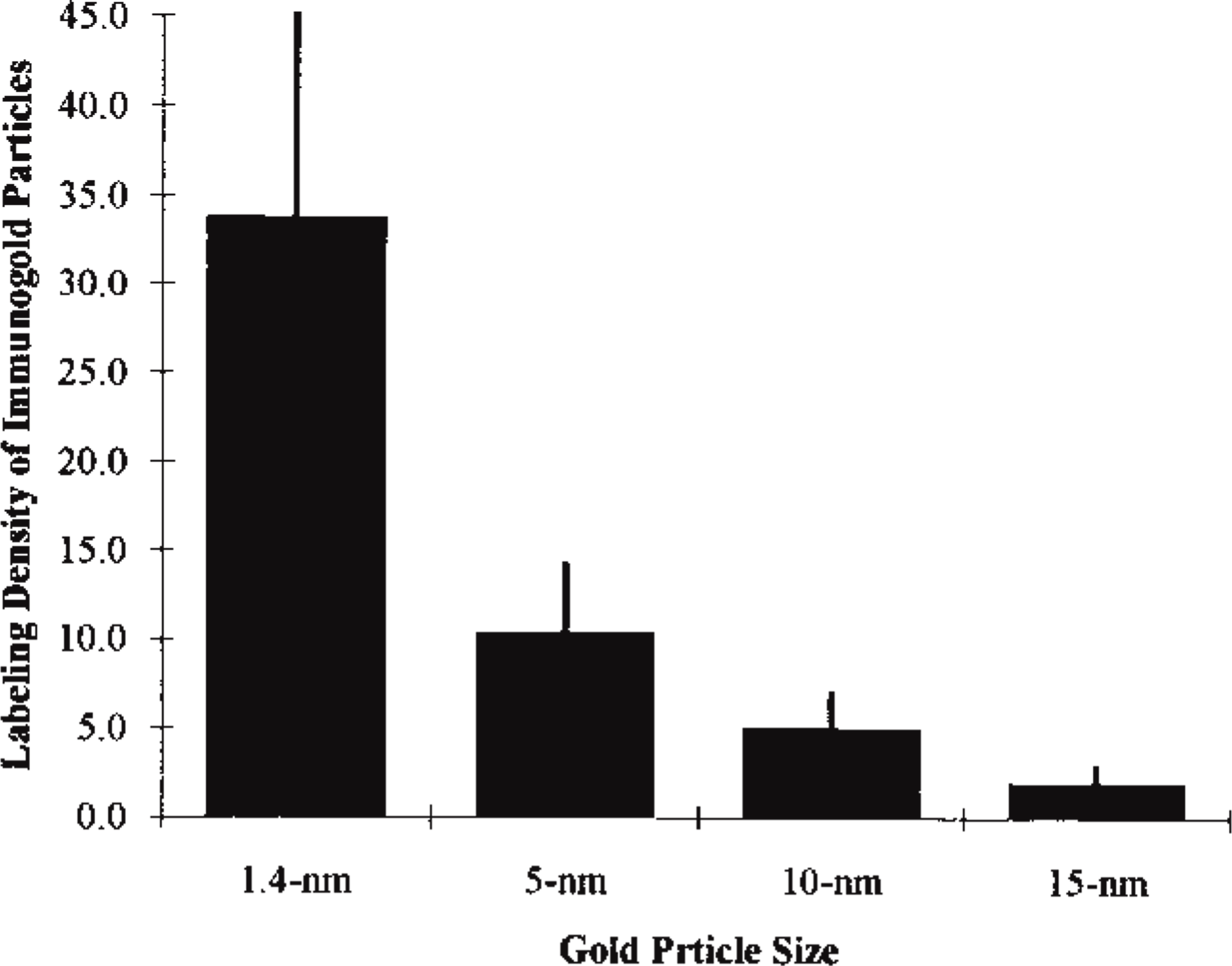
Graph comparing the labeling of HLA Class I using 1.4-, 5-, 10-, and 15-nm immunogold probes. In each case, enlargements of 35 micrographs of human neutrophil split plasma membranes (E-faces) were analyzed. The y-axis is calibrated as the number of gold particles/μm2 of the E-face.
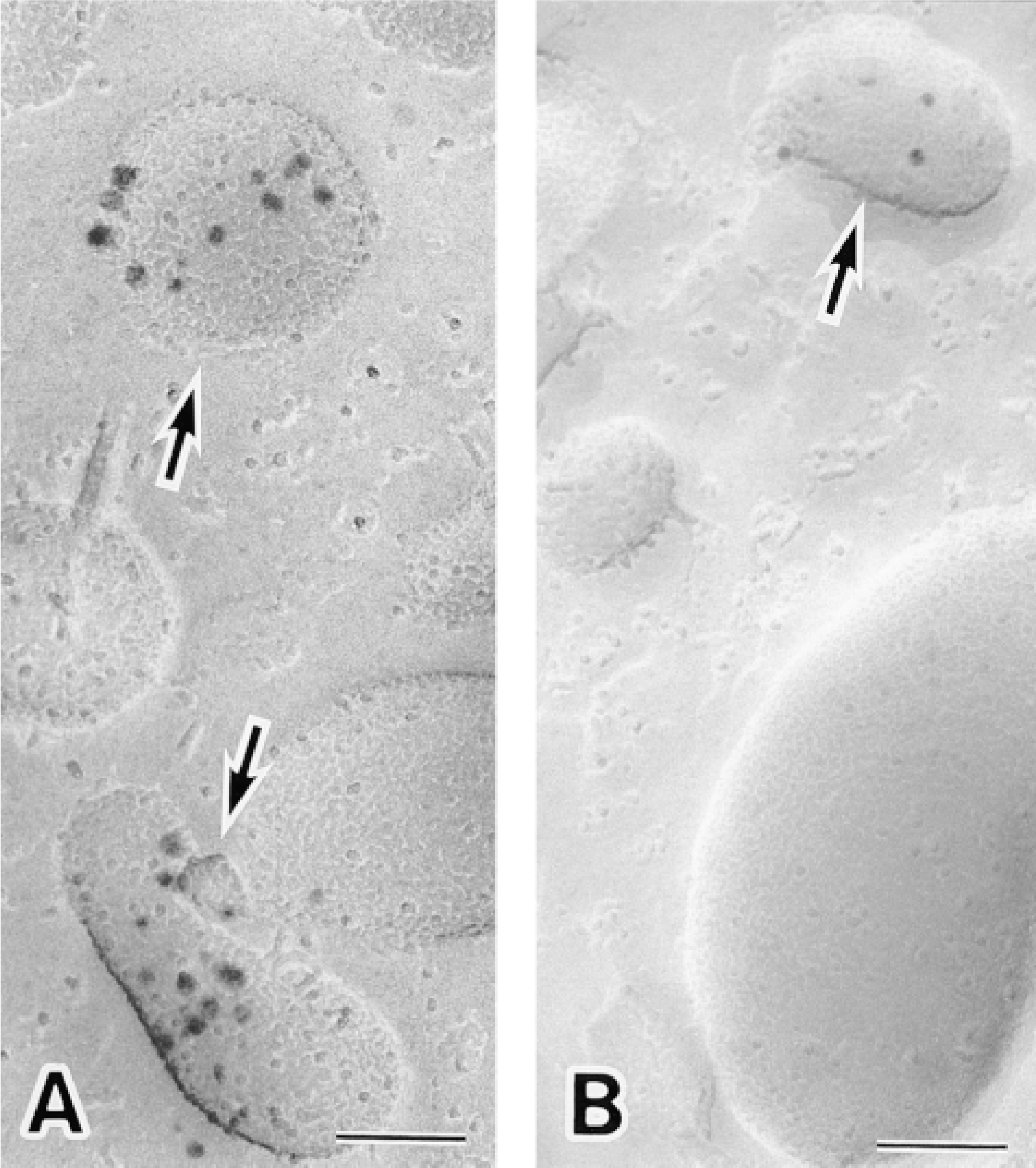
Intracellular localization of CD16 in unstimulated human neutrophils. (A) Replica immunocytochemical labeling of CD16 achieved by the use of 1.4-nm gold-attached secondary antibody with subsequent silver enhancement. Silver-enhanced 1.4-nm gold particles depicting the distribution of CD16 are located on intracellular granules (arrows). CD16-containing granules are readily recognized using this technique. (B) Replica immunocytochemical labeling of CD16 with 10-nm gold. Note that only three gold particles are present on the E-face of this intracellular granule (arrow). Bars = 0.1 μm.
Discussion
Freeze-fracture immunocytochemical techniques that make use of colloidal gold immunoprobes have proved valuable for visualizing the macromolecular organization of biomembranes (Fujimoto 1997; Takizawa et al. 1998). However, when we used 10- or 15-nm colloidal gold probes as the secondary detection system for this type of immunocytochemistry, labeling was sometimes poor. As was seen in the present study with HLA Class I and CD16, this was particularly problematic with several antigens in neutrophil replicas, where antigens might be represented by only one or two gold particles. This low level of labeling led to difficulties in interpretation. We questioned these findings and hypothesized that the apparent distribution of antigen was attributable to characteristics of the immunogold probes (e.g., gold particle size).
We found that the labeling density for HLA Class I was inversely related to the size of the immunogold probe (see Figure 3) and confirmed that greater immunogold labeling efficiency was achieved when smaller gold particles were used. Earlier studies using other immunocytochemical techniques, but also employing immunogold probes, reported a similar inverse relationship between labeling density and the size of the immunogold particles (for review see Robinson et al. 1998).
We also discovered that replica membranes were neither degraded nor themselves enhanced by the silver enhancement method of Burry (1995), which we employed. This meant that all of the observed electron-dense particles on the replicas were attributable to silver-enhanced Nanogold localization of either HLA Class I or CD16. Therefore, when followed by silver enhancement, 1.4-nm gold was an effective immunoprobe for replica immunolabeling.
We have demonstrated that 1.4-nm immunogold probes used in conjunction with silver enhancement can yield reliable immunocytochemical labeling of replicas. Because freeze-fracture cytochemistry, especially freeze-fracture immunocytochemistry, is an important technique for studying the topology and dynamics of membrane molecules, the method described here should prove to be a useful addition.
Footnotes
Acknowledgements
Supported by grants from the Kazato Research Foundation and the Nippon Foundation, and by Grants-in-Aid for Scientific Research from the Ministry of Education, Science, Sports, and Culture of Japan.
I am deeply indebted to Dr John M. Robinson of Ohio State University for critical review of the manuscript. I am grateful to Ms Kiyomi Inose, Ms Michiyo Soutome, and Ms Megumi Yatabe for excellent technical assistance.