
Abstract
It has recently been postulated that platelet/endothelial cell adhesion molecule-1 (PECAM-1/CD31) might play a role in vascular tube formation. To evaluate the role of PECAM-1/CD31 in the formation of the capillary network in vivo, we conducted an ultrastructural immunohistochemical evaluation of the localization of PECAM-1/CD31 and its developmentally regulated expression in the periphery of the lungs of fetal, newborn, and adult rats. PECAM-1/CD31 was present mainly on luminal surfaces and at the junctions between endothelial cells. Moreover, in fetal lung, products of the immunoreaction were also found on the abluminal surfaces of endothelial cells. To relate those findings to the developmental changes in the capillary area of the lung, we performed a morphometric study of electronmicrographs. The cross-sectional area of blood vessels at the periphery of the lungs was significantly greater in 15–19-day-old fetuses than in postpartum animals (p < 0.0001). Disappearance of the expression of PECAM-1/CD31 on the abluminal endothelial surface paralleled the changes in the cross-sectional area of blood vessels that occurred during the perinatal period.
P
Quantitative investigations of the postpartum development of the rat lung have been performed since appropriate morphometric tools became widely available (Burri et al. 1974). During the past decade, great progress has been made in the development of computerized tools for quantitative morphology, but studies focusing on lung structure have been limited (Bolender et al. 1993; Marszalek et al. 1999; for review see Burri and Moschopulos 1992). Such studies have focused mainly on developmental changes in the correlation between air space and vascular volume (Adamson and King 1984; Ting et al. 1998). There have been few morphometric studies on the development of lung tissue compartments in correlation with factors involved in lung growth.
The aim of this study was to investigate the maturation of the vascular network of the rat lung by monitoring the presence of PECAM-1/CD31 on endothelial cell surfaces using an immunoelectron microscopic technique. Furthermore, an ultrastructural morphometric analysis of changes in vascular area was performed during lung development using 15-day-old fetuses, postnatal, and adult rats.
Materials and Methods
Animals
Five-day-pregnant, specific pathogen-free Wistar rats (body weight 245–280 g) were provided by Seiwa Experimental Animals (Fukuoka, Japan). Rats were determined to be pregnant upon detection of spermatozoa in vaginal smears after overnight mating. The first 24 hr after detection of spermatozoa were defined as Day 1. During the experiment, each dam was kept with her offspring in a single cage with a 12 hr/12 hr light/dark cycle at 20–24C, with relative humidity maintained at 60 ± 10%. Standard rat chow and water were provided ad libitum. Before removal of fetuses, dams were anesthetized with an IP injection of sodium pentobarbital (Abbott; Dainabot, Osaka, Japan). Immediately after removal by cesarean section, fetuses were sacrificed by spinal dislocation. Before the thorax was opened, the trachea was clamped and was occluded until the end of fixation. Fetal lungs were removed from the thorax by thoracotomy and immediately placed in fixative. Tissue removal and fixation were performed carefully to avoid any compression. Lungs from newborn and adult animals were removed by thoracotomy after IP injection of sodium pentobarbital. We prepared lungs from 15-, 17-, 19-, and 21-day-old fetuses and from 1-, 3-, 5-, 7-, 14-, 21-day-old and 2-month-old animals (body weight >240 g). We studied tissues from five animals in each group. No more than two animals in any group were from the same litter.
Electron Microscopy
For transmission electron microscopy, tissues were prepared by standard methods. In brief, after removal, lungs were immersed immediately in freshly prepared Karnowsky fixative at 4C and incubated for 8 hr (with several changes of fresh fixative). Small samples of tissue (1 × 1 × 1 mm) were taken and washed in several changes of phosphate buffer (PB) at 4C. After overnight washing in PB, these samples were post-fixed in 1% OsO4 for 2 hr and dehydrated in a graded series of ethanol–propylene solutions. Finally, the lung samples were embedded in Epok 812 resin (Oken; Tokyo, Japan). After polymerization of the resin, semithin sections were cut, stained with toluidine blue, and examined by light microscopy. Ultrathin sections were stained with uranyl acetate and lead citrate and were examined by transmission electron microscopy (JEM 100CX; JEOL, Tokyo, Japan).
Immunoelectron Microscopy
For immunoelectron microscopy, we used lungs that had been fixed in 4% paraformaldehyde in 0.2 M PBS. Immediately after removal, lungs were incubated in freshly prepared cold (4C) fixative for 4 hr (with at least three changes of fixative). Then samples (5 × 5 × 10 mm) were cut from the lower lobes and washed for 12–18 hr in several changes of 0.2 M PBS that contained 10% sucrose (PBS-S). The samples were finally washed in PBS-S containing 7% glycerol for 1 hr, mounted in Tissue-Tek OCT compound (Sakura Finetechnical; Tokyo, Japan), and rapidly frozen in dry ice and acetone. They were stored at −80C before analysis. For immunostaining, 5-μm sections that had been air-dried for 30 min were washed four times (for 5 min each) in PBS at 4C. Endogenous peroxidase activity was quenched by incubation in 0.2% H2O2 at room temperature (RT) for 30 min. The samples were washed in PBS at 4C. To block free aldehyde groups, samples were incubated in a 0.05% solution of potassium borohydrate (Chameleon Reagent; Osaka, Japan) in PBS at RT for 1 hr. After washing in PBS at 4C (four times for 5 min), samples were incubated with 10% normal goat serum for 15 min at RT. They were next incubated overnight with primary antibody (mouse monoclonal antibody against rat PECAM-1/CD31) (MA-3107; Endogen, Woburn, MA) at a dilution of 1:50 in a humid chamber at 4C. The next day, after washing in PBS at 4C (four times for 5 min), the samples were incubated with the peroxidase-conjugated F(ab')2 fragment of goat antibody against mouse IgG (ICN Pharmaceuticals; Aurora, OH) at 1:100 dilution for 2 hr at RT. After incubation with the second antibody, the samples were washed in PBS at 4C (four times for 5 min), postfixed in 1% glutaraldehyde for 5 min at 4C, and washed again. They were treated with a 0.02% solution of 3,3′-diaminobenzidine tetrachloride (DAB) and 1% DMSO in 0.05 M Tris-HCl (Sigma Chemical; St Louis, MO) buffer (pH 7.2) for 30 min and then with a 0.02% solution of DAB in 0.05 M Tris-HCl buffer (pH 7.2) that contained 0.005% H2O2 for 6–10 min. The samples were washed in PB, post-fixed in 1% OsO4 for 1 hr, and washed again in PB. Finally, after dehydration in a graded ethanol series, the sections were embedded in Epok 812. Epok blocks were trimmed, cut into ultrathin sections, and observed, without additional staining, by electron microscopy. In all cases, negative controls for immunohistochemical staining were provided by replacement of primary antibodies by PBS or by nonimmune mouse serum, or by omitting the chromogen (DAB).
Morphometry
To fulfill the requirements for uniformity and randomness of the sample, we used criteria described earlier by Bolender et al. (1993). For morphometric measurements, we selected lung tissue samples taken only from the base of of the lower lobes in all groups, because they can represent the same part of the lung. It is extremely important for the developing lung, as described earlier (Burri and Moschopulos 1992), that different parts show different stages of development in the same growing organ. All selected vascular profiles contained endothelial cells with a nucleus and had an open lumen. From several tissue blocks, 50 electron micrographs were selected for each age group. No more than 10 pictures were chosen from the blocks of tissue for each animal. Selected pictures were taken from at least five animals in each age group. All electron micrographs had the same magnification (×3400). On the positives (negatives during processing were magnified ×6.0), the borders of the capillaries (lumen plus endothelium) and the borders of the capillary lumina were manually delineated. Then, through the use of a scanner (GT-9000; Epson, Tokyo, Japan), pictures were saved on the hard disk (as 8-byte ∗.pict files) of a computer (Power Macintosh 8600250; Apple Japan, Tokyo, Japan). Using NIH Image 1.52 software (National Institutes of Health, Bethesda, MD), we transformed and converted the pictures into binary pictures. On the two-color images (black and white), using a wand, we selected previously delineated borders of the area of each capillary [endothelium plus lumen; S(el)] and that of the capillary lumen [S(l)], and these areas were measured automatically. The area occupied by endothelial cells [S(e)] that formed the capillary wall was calculated as S(e) = S(el) – S(l) for each vessel.
Statistical Analysis
The comparison of multiple groups was made according to the rules described by Ryder and Robakiewicz (1998). As a test of statistical significance, we performed an analysis of variance (ANOVA). For individual comparisons, we used the Bonferroni test. The statistical significance of differences was recognized at p < 0.0001. For all statistical calculations, we used Data Desk v4.1 (Data Description; Ithaca, NY).
Results
Immunolocalization of PECAM-1/CD31
Products of a positive immunoreaction specific for PECAM-1/CD31 were found in the blood vessels of all age groups studied. There was no immunostaining in the negative controls (data not shown). Reaction products were seen mostly on the luminal surfaces of endothelial cells (Figures 1A and 1D) and also at junctions between neighboring endothelial cells in all groups (Figures 1B and 1C). In specimens from fetuses between 15 and 21 days after conception, reaction products were also noted on the abluminal surfaces of endothelial cells (Figures 1A–1C), but such was not the case in postpartum rats (Figure 1D). Weak evidence of immunoreaction products was infrequently noted in pinocytotic vesicles of endothelial cells (Figures 1B and 1C) in fetuses and newborns. PECAM-1/CD31 was detected by light microscopy in all types of blood vessels (except controls) in the materials studied (data not shown), but the intensity of immunostaining decreased with increasing age of the rats.
Morphometry
Examples of capillaries selected for morphometric studies are shown in Figure 2. Morphometric results for all groups are shown in Figure 3. The areas of all measured compartments tended to decrease with increasing age of fetuses and in postpartum animals. The initial mean vessel area in a cross-section of capillaries in lung buds was (mean ± SE) 79.6 ± 6.6 μm2 on fetal Day 15, and in adult animals it had fallen to 23.32 ± 1.9 μm2. In the capillary lumen, the mean areas for the youngest (fetal Day 15) and oldest (adult) groups were 45.9 ± 5.3 and 14.2 ± 1.4 μm2, respectively. The area of the endothelial compartment also decreased from an initial value of 33.7 ± 2.2 μm2 on fetal Day 15 to 9.1 ± 1.0 μm2 in adults. The most dramatic decrease in measured areas occurred from fetal Day 15 to delivery (fetal Day 21 and 1-day-old newborns). In general, areas of entire capillaries, their lumina, and endothelial cells were significantly greater in 15–19-day-old fetal rats than in rats just before birth and in newborn to adult rats (for details see Figures 3A–3C). The decrease in mean areas was clearest in endothelial cells when 19-day-old and 21-day-old fetuses were compared (Figure 3C). It represented the flattening of the endothelial cells in capillaries that prepared them for gas exchange. There were no significant differences among the various groups of postpartum animals, although small fluctuations were observed. The largest standard errors were obtained from the first three groups (15-, 17-, and 19-day-old fetuses) in measurements of all analyzed parameters (Figures 3A–3C). This phenomenon might be related to the extraordinary variability in the size of vessels in the mesenchyme of the dynamically developing lung. In 21-day-old fetuses and postpartum animals, smaller standard errors reflected the greater uniformity in terms of size of the septal capillaries.
Discussion
In this study, using an in vivo model of developmental angiogenesis, we found that expression of the PECAM-1/CD31 molecule changes during lung formation. The exact function of PECAM-1/CD31 in vivo remains unclear, even though several investigations in vitro and in vivo have been reported in the past decade (Albelda et al. 1990, 1991; DeLisser et al. 1993, 1997). The fetal expression of PECAM-1/CD31 on the abluminal surfaces of endothelial cells found in this study might reflect a role of that molecule in endothelial cell migration in vivo. It was also suspected by Sheibani and Frazier (1999) that this cell adhesion molecule might play a role in cell–matrix interactions and that vascular formation also depends on extrinsic factors (Noden 1990; Baldwin et al. 1994). The phenomenon of positively immunostained regions of abluminal endothelial membranes was not observed in postpartum animals when air–blood barriers were formed. This might reflect the PECAM-1/CD31 redistribution that was described by Schimmenti et al. (1992) in vitro, which occurs during vascular tube formation. Similar findings, as described above, during lung formation were found during development of the murine blood–brain barrier (Vorbrodt et al. 1986; Lossinsky and Wisniewski 1998; Lossinsky et al. 1999). Those changes in the expression of the adhesion molecules in different biological barriers probably reflect a uniform model of formation of vascular networks.
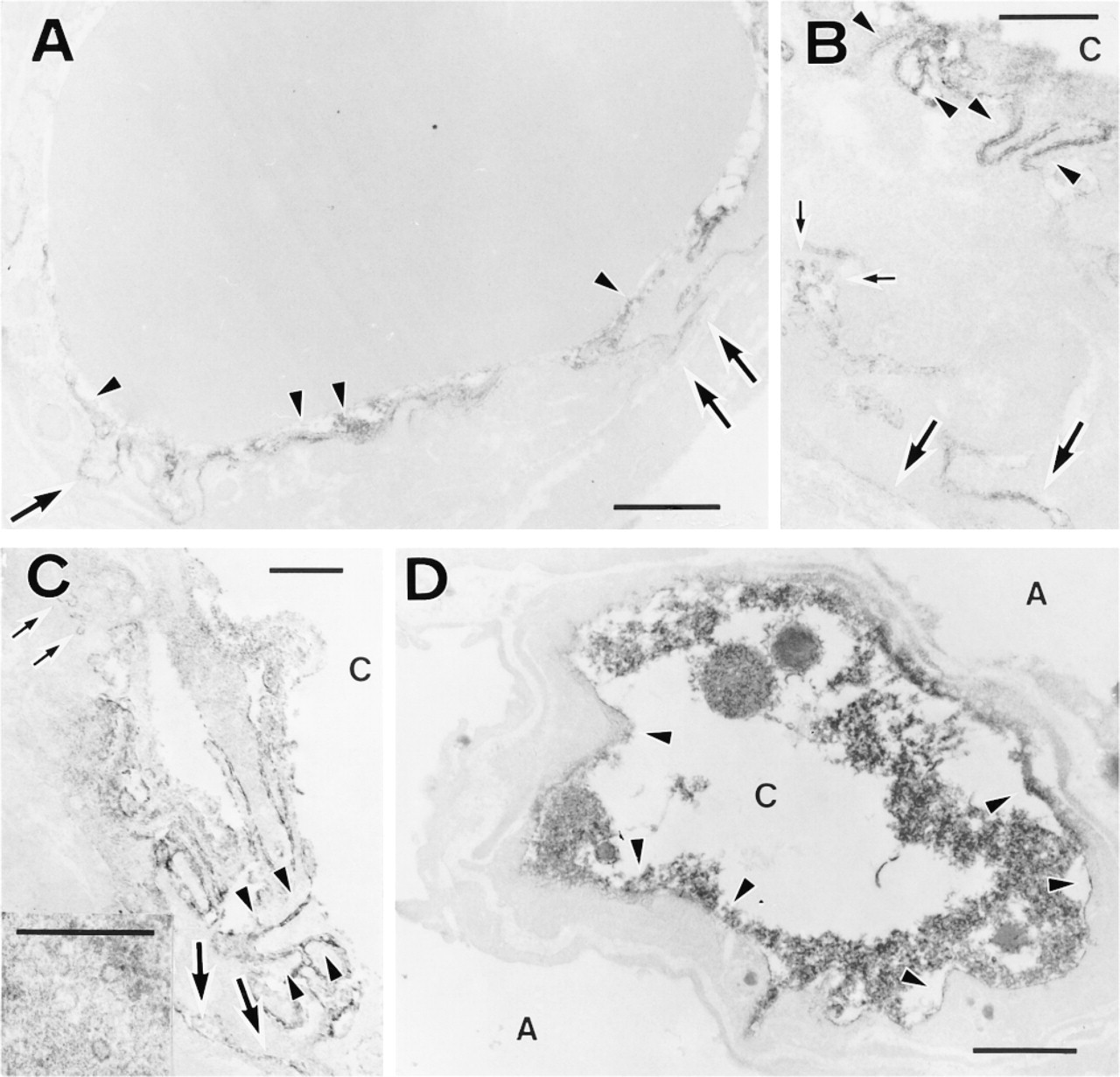
Electron microscopic immunoperoxidase localization of PECAM-1/CD31 in capillaries of the developing lung. (
Formation of the capillary tubes depends on endothelial cell–cell contacts. PECAM-1/CD31 has been described as one of the key molecules in that process (Albelda et al. 1991; Fawcett et al. 1995). In this study, we found expression of PECAM-1/CD31 in the junctions between neighboring endothelial cells in both fetal and postpartum tissues. We observed different levels of expression of that molecule in the capillaries of developing lungs at the light microscopic level (unpublished data). This is consistent with previously published reports that PECAM-1/CD31 expression is developmental regulated (Watt et al. 1995; Sheibani et al. 1999). Recently published data (Matsumura et al. 1997; Yang et al. 1999) suggest that PECAM-1/CD31 is only a part of a multimeric protein structure called the adherens junction. The same authors provide evidence that this structure is formed by the interaction between VE-cadherin (cadherin-5/CD144), β-catenin, F-actin, and PECAM-1/CD31. Molecules in the PECAM-1 and VE-cadherin complex were described as critical for endothelial tube formation (Matsumura et al. 1997; Yang et al. 1999). These data require future confirmation using in vivo developmental models.
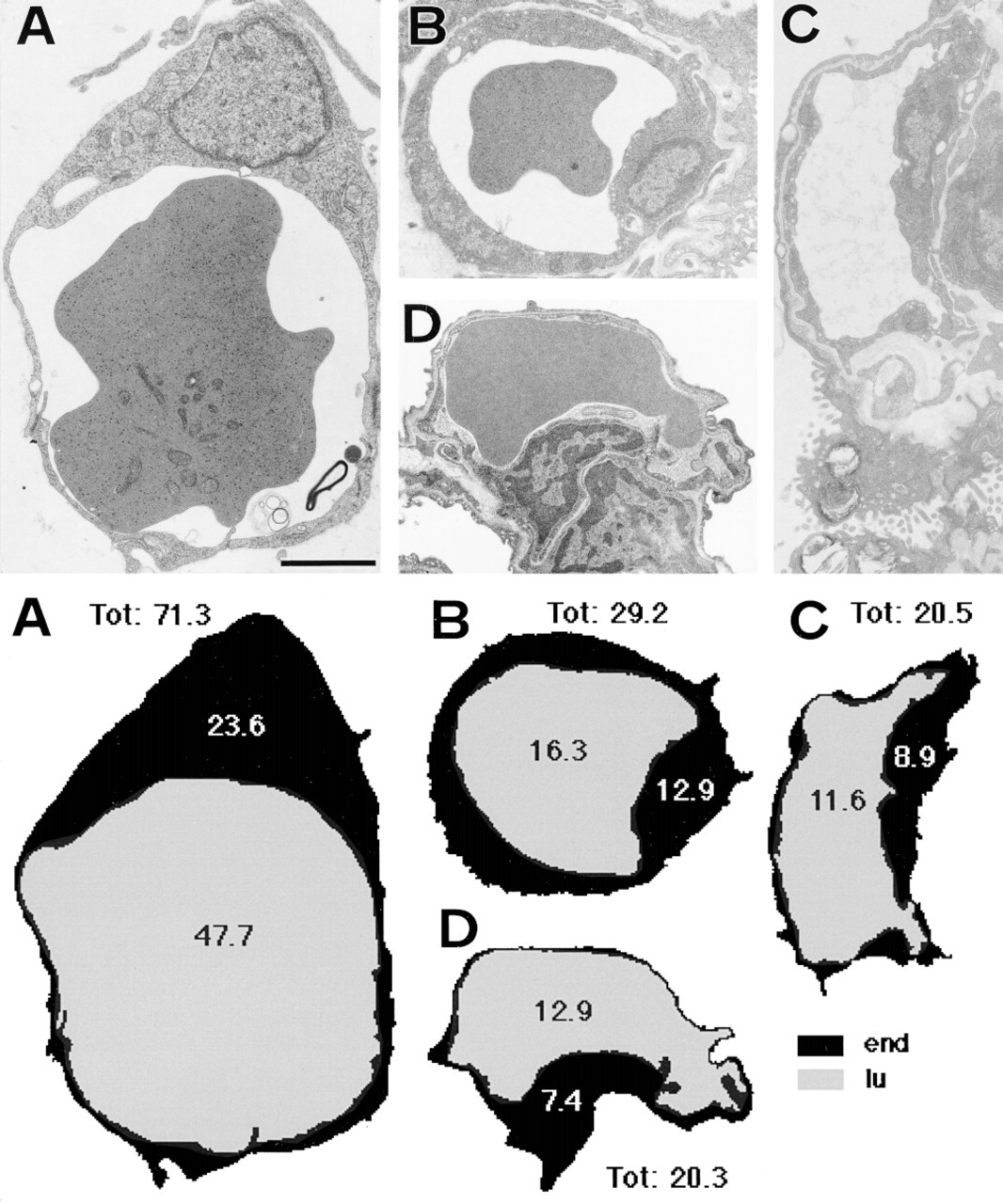
Examples of electron micrographs of lung vessels selected for morphometric studies from Karnovsky-fixed tissue samples and related schematic images with the results of measurements (in μm2) on given electron micrographs. The time period of lung tissue samples is as follows: (
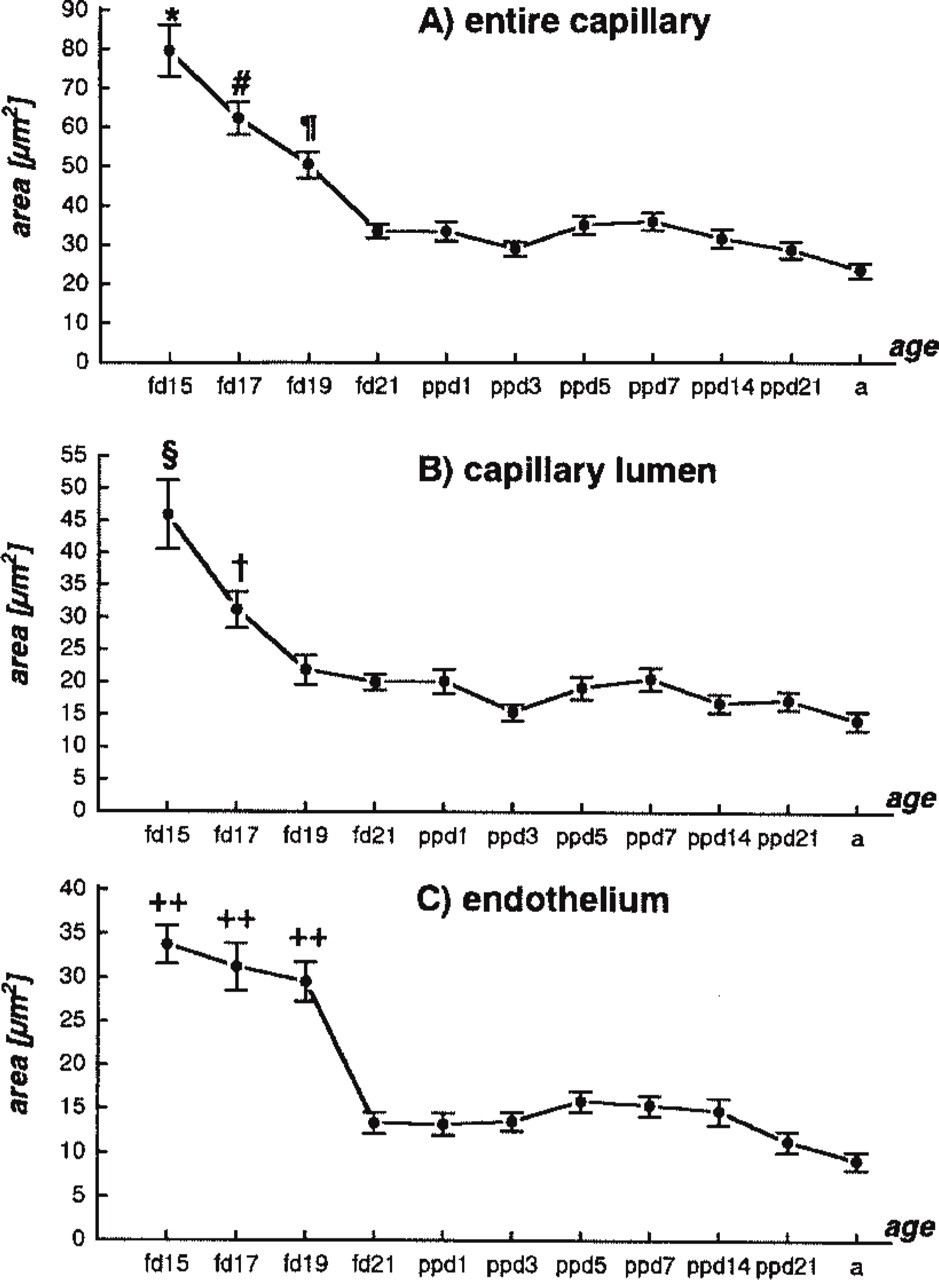
Morphometric measurements of capillary area during fetal and neonatal lung development. Values represent means ± SE. Significant difference (p < 0.0001) is found in
Because the morphometric estimation of whole-lung volume changes during development in rats had been published earlier (Burri et al. 1974; Moschopulos and Burri 1993), we focused on the changes in the capillary cross-sectional area in the periphery of the lung during fetal and postpartum life. Knowing the limitations and the validity of tissue sampling for the morphometric studies of the lung published earlier (Gil 1978, 1988; Hislop et al. 1984; Zeltner et al. 1990; Ciurea and Gil 1996), we applied those data to the observed changes in the expression of PECAM-1/CD31. The development of lung capillary networks leads to the formation of an air–blood barrier. First barriers in rats can be observed just before delivery (Han et al. 1992; Wasowicz et al. 1996). In our study, this was accompanied by a significant decrease in capillary area parameters. However, as seen earlier, the total vascular surface and volume in the lungs increases dramatically during the perinatal period (Moschopulos and Burri 1993; Marszalek et al. 1999). Moreover, the morphometric changes found in the present study paralleled those in the expression of the PECAM-1/CD31 molecule. This leads us to conclude that PECAM-1/CD31 plays an important role during the proliferative phase of vascular network formation in the lung until air–blood barriers appear. It is then downregulated after birth.
This report is the first, to our knowledge, on the ultrastructural localization and expression of PECAM-1/CD31 during development of the lung vasculature. However, in the light of recent publications, new studies on the role of other molecules that might act in conjuction with PECAM-1/CD31 are needed for a better understanding of the developmental growth of these vascular networks in vivo.
Footnotes
Acknowledgements
A.M. was a recipient of a scholarship from the Japanese government (MONBUSHO; Ministry of Education).
We wish to thank Mr S. Yano for excellent assistance in the handling of the animals, and for technical support.