
Abstract
We examined the expression and localization of the prohormone convertases, PC1 and PC2, in the ultimobranchial gland of the adult bullfrog using immunohistochemical (IHC) and in situ hybridization (ISH) techniques. In the ultimobranchial gland, PC1-immunoreactive cells were columnar, and were present in the follicular epithelium. When serial sections were immunostained with anti-calcitonin, anti-CGRP, anti-PC1, and anti-PC2 sera, PC1 was found only in the calcitonin/CGRP-producing cells. No PC2-immunopositive cells were detected. In the ISH, PC1 mRNA-positive cells were detected in the follicle cells in the ultimobranchial gland. No PC2 mRNA-positive cells were detected. RT-PCR revealed expression of the mRNAs of PC1 and the PC2 in the ultimobranchial gland. However, very little of the PC2 mRNA is probably translated because no PC2 protein was detected either by IHC staining or by Western blotting analysis. We conclude that the main prohormone convertase that is involved in the proteolytic cleavage of procalcitonin in the bullfrog is PC1.
Keywords
C
Alternative processing of RNA transcripts from the calcitonin gene leads to the production of distinct mRNAs encoding calcitonin or CGRP (Amara et al. 1982). A sequence analysis of these mRNAs revealed that procalcitonin is cleaved at the Lys-Arg and Gly-Lys-Lys-Arg sites to produce calcitonin, CCP, and 83-amino-acid N-terminal peptide, and that proCGRP is cleaved at two cleavage sites, Lys-Arg and Gly-Arg-Arg-Arg, to generate CGRP (Amara et al. 1980,1982).
Prohormone convertases, PC1 (also called PC3) and PC2, are responsible for the proteolytic cleavage of a large precursor molecule at paired basic sites to yield bioactive peptides in endocrine cells and neurons (Seidah and Chrétien 1992). Immunocytochemical and ISH studies have shown that these convertases are expressed in the pancreatic islets (Malide et al. 1995; Tanaka et al. 1996; Rawdon and Larsson 2000), in the pituitary gland (Braks et al. 1992; Marcinkiewicz et al. 1993; Kurabuchi and Tanaka 1997; Uehara et al. 2001), and in the brain (Schafer et al. 1993; Birch et al. 1994; Vieau et al. 1998; Gangnon et al. 1999). Recently, we demonstrated that both PC1 and PC2 were located in the mouse calcitonin/CGRP-producing parafolliclular cells and that a subpopulation of the PC1-immunoreactive cells contains CGRP in the mouse respiratory tract, suggesting that procalcitonin is proteolytically cleaved by PC2 together with PC1, and proCGRP by PC1 alone. In anuran amphibians, we showed that ultimobranchial calcitonin regulates expression of otoconin mRNA, which is involved in the formation of calcium carbonate crystals in the endolymphatic sac (Yaoi et al. 2003b). Therefore, amphibian calcitonin has a unique biological action, but it has not yet been fully confirmed whether PC1 and/ or PC2 are involved in the limited proteolysis of the precursors of this calcitonin. Immunocytochemical examination of PC1 and PC2 would also provide important evidence for the substrate specificity of these convertases in processing of the procalcitonin.
In this study we examined the expression and localization of PC1 and PC2 in the bullfrog ultimobranchial gland and found a correlation between the presence of PC1 and the presence of calcitonin.
Materials and Methods
Animals
Adult male bullfrogs (Rana catesbeiana) were purchased from Ouchi (Misato, Japan). They were acclimated under normal laboratory conditions for at least 1 week before use. They were fed pieces of porcine liver twice a week. The ultimobranchial glands, dissected under anesthesia with MS-222 (Nacalai Tesque; Kyoto, Japan), were used for histochemical examination and RT-PCR analysis. All animal experiments were in compliance with the Guide for Care and Use of Laboratory Animals in Shizuoka University.
Antibodies
The antibodies used in this study were prepared in rabbits and characterized as described previously: anti-PC1 serum (ST-28) and anti-PC2 serum (ST-29) against synthetic peptides covering amino acids 442–459 and amino acids 613–629 of the respective mouse proteins (Tanaka et al. 1996), anti-salmon calcitonin serum (a gift from Prof. K. Wakabayashi, Gunma University), and anti-synthetic rat CGRP (1–37) serum (Cambridge Research Biochemicals; Cheshire, UK).
Light Microscopic Immunocytochemistry
The glands were fixed by immersion in Bouin-Hollande for 2 days. After dehydration and embedding in Paraplast, serial 4-μm-thick sections were cut and mounted on gelatin-coated slides. The deparaffinized sections were incubated in a solution of 0.3% H2O2 in methanol for 30 min to inhibit endogenous peroxidase activity. After rinsing with distilled water (DW) followed by PBS (0.01 M sodium phosphate buffer and 0.14 M NaCl, pH 7.5), the sections were immunostained by the indirect enzyme-antibody method. To amplify the signals for PC1 and PC2, the sections were treated with Gomori's oxidation mixture according to the procedure of Kurabuchi and Tanaka (1997). The sections were incubated sequentially at room temperature (RT) with the following reagents; 10% normal goat serum (NGS) for 1 hr, rabbit anti-PC1 (1:2000) or PC2 (1:2000) serum for 16 hr, and peroxidase-conjugated goat anti-rabbit IgG for 1.5 hr. Peroxidase activity was detected using 15 mg 3,3′-diaminobenzidine tetrahydrochloride (Dojin Lab; Kumamoto, Japan) and 0.01% H2O2 in 100 ml of 0.05 M Tris-HCl buffer, pH 7.6. The sections were lightly counterstained with Mayer's hematoxylin, dehydrated with ethanol, and then mounted in Entellan (Merck; Gobbstown, NJ).
Adjacent serial sections were immunostained to identify calcitonin-producing cells using the same method with rabbit anti-salmon calcitonin antiserum (1:8000) and anti-CGRP serum (1:2000), respectively.
The specificity of the anti-PC1 and anti-PC2 sera was also tested using an immunocytochemical (ICC) preabsorption test. Diluted antisera were preabsorbed with their corresponding peptides at a final concentration of 10 μg/ml at 4C for 16 hr before ICC studies.
Western Blotting Analysis
The ultimobranchial glands or pituitary neurointermediate lobes from the bullfrogs were homogenized in cell lysis buffer [50 mM Tris-HCl (pH 8.0), 0.15 M NaCl, 1% Triton X-100, 0.1 mg/ml PMSF, 1 μg/ml aprotinin] and centrifuged in a microcentrifuge for 5 min to remove insoluble materials. The proteins were quantified with a BCA Protein Assay Kit (Pierce; Rockford, IL). The supernatant protein (10 μg) was denatured at 100C for 10 min in denaturation buffer comprising 2% SDS, 25 mM Tris-HCl, pH 7.5, 25% glycerol, and 0.005% bromophenol blue, subjected to electrophoresis on a 10% polyacrylamide gel, and then transferred to an Immobilon-P membrane (Millipore; Tokyo, Japan). The proteins in the membrane were reacted sequentially with rabbit anti-PC1 or PC2 serum diluted at 1:5000, biotinylated anti-rabbit IgG (DAKO; Kyoto, Japan), and streptavidin-conjugated horseradish peroxidase (DAKO). The reaction product on the membrane was visualized with an ECL Western blot detection kit (Amersham Pharmacia Biotech; Poole, UK). As a control, the primary antibody was replaced with anti-PC1 serum preincubated with 10 μg/ml of the antigen peptide.
RT-PCR of Bullfrog Ultimobranchial Gland
The tissue expression of PC1 and PC2 mRNAs was analyzed by RT-PCR. Total RNA was prepared, using the TRIZOL reagent (Life Technologies; Rockville, MD), from the bullfrog ultimobranchial gland and neurointermediate lobe. After treatment of 20 μg total RNA with DNase I (4 U; Takara, Kyoto, Japan), a 10-μg aliquot of the total RNA was reverse-transcribed in 20 μl of reaction buffer containing 1 mM each of dNTP, 9.9 U of RAV-2 reverse transcriptase (Takara), 20 U of RNase inhibitor (Toyobo; Osaka, Japan), 7.5 mM of oligo-dT(19)primer (Life Technologies) at 42C for 1 hr and then at 52C for 30 min. RT-PCR was performed using degenerate primers, designed based on the amino acid sequence of the bullfrog PC1 and PC2, respectively: PC1 sense 5′-GTAGGAGGCATTCGGATGTTA-3′ (809–829 b) and antisense 5′-GAAGATTGAGCCTTT-TCCATTT-3′ (994–1015 b); PC2 sense, 5′-TCTTCC-ACTTTAGCCTCTACAT-3′ (1197–1218 b) and anti-sense 5′-CTCTAGGGCTAATGCAAACA-3′ (1333–1352 b). The RT-PCR products were analyzed on a 2% agarose gel containing ethidium bromide (EtBr; 0.5 μg/ml) with Marker 6 (λ/Sty1 digest; Wako Pure Chemicals, Osaka, Japan) for molecular weight markers.
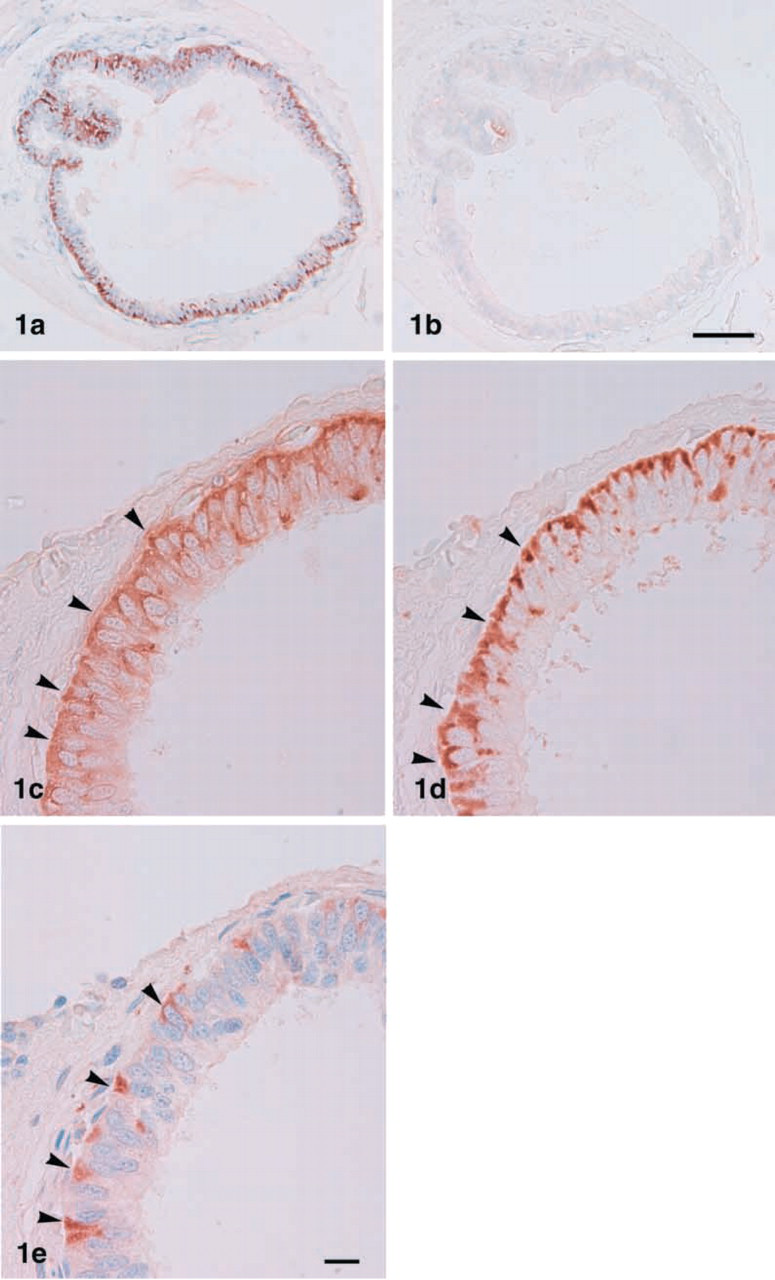
Light micrographs showing immunostaining for PC1 (
ISH Histochemistry
DIG-labeled antisense and sense cRNA probes were prepared from the full-length coding region of PC1 and PC2 cDNAs by in vitro transcription, as described previously (Saito et al. 2002). Bullfrog ultimobranchial glands were fixed with 4% paraformaldehyde (PFA) in 0.1 M phosphate buffer, pH 7.4, overnight at 4C. After fixation the tissues were dehydrated through a graded alcohol series, cleared in methyl benzoate-celloidin, and embedded in Paraplast. Sections were cut at 4-μm thickness and mounted on silane-coated slides. ISH was carried out according to a method described previously (Saito et al. 2002). Briefly, the deparaf-finized sections were digested with 5 μg/ml proteinase K for 20 min, fixed in 4% PFA for 20 min, and then incubated with the DIG-labeled cRNA at 50C for 15 hr. After hybridization, the sections were treated with 1 μg/ml RNase solution for 30 min and then incubated with alkaline phosphatase-conjugated sheep anti-DIG Fab antibody (Roche) for 15 hr. The label was detected with nitroblue tetrazolium chloride and 5-bromo-4-chloro-3-indolylphosphate (Roche).
Dual mRNA and Protein Staining
After the PC1 mRNA was stained as described above, the sections were washed with PBS and incubated with rabbit anti-salmon calcitonin overnight, followed by Cy3-labeled donkey anti-rabbit IgG (Jackson Immunoresearch; West Grove, PA) for 2 hr. The sections were washed with PBS and then mounted in PermaFluor (Immunon; Pittsburgh, PA), and examined under an Olympus BX50 microscope equipped with a BX-epifluorescence attachment (Olympus Optical; Tokyo, Japan).
Results
Localization of Prohormone Convertase and Calcitonin
The bullfrog ultimobranchial gland consists of one to several follicles. Calcitonin-producing cells were visible in the pseudostratified epithelium forming the follicle. PC1-immunopositive cells were observed in the ultimobranchial glands, but no PC2 cells were seen (Figure 1a and 1b). In the absorption test, the PC1-immunopositive materials obtained with the antiserum were completely eliminated when 10 μg/ml of the respective antigen peptide was used as an adsorbent (data not shown). When consecutive sections were immunostained for PC1 and calcitonin, PC1 immunopositive cells were found in calcitonin-producing cells (Figure 1c and 1d). Careful observation revealed intense PC1 immunoreactivity in the basal cytoplasm side of the calcitonin-producing cells. In addition, a subpopulation of calcitonin-producing cells was also immunostained with anti-CGRP (Figure 1e).
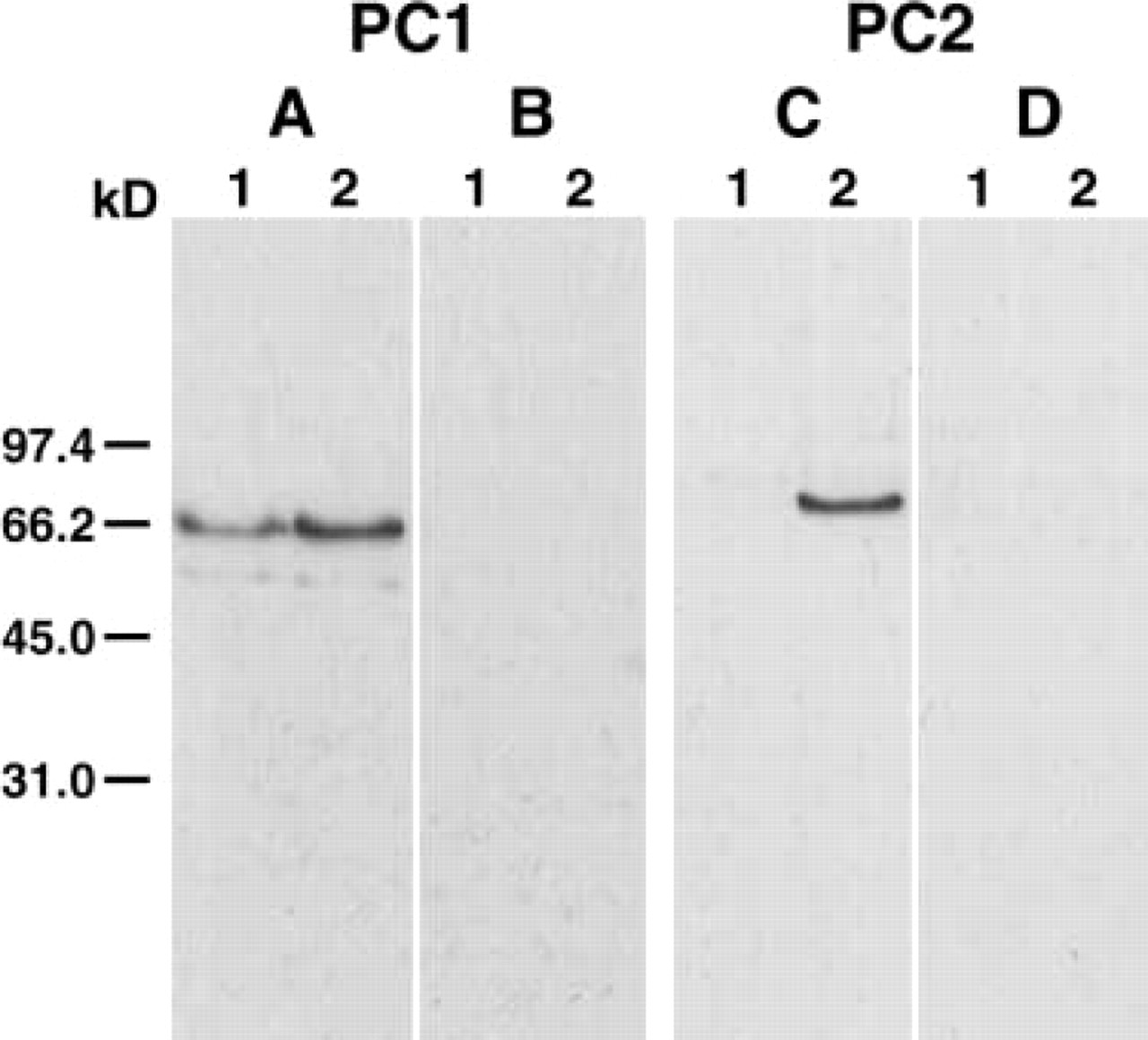
Characterization of anti-PC1 and anti-PC2 sera by Western blotting analysis. (
Antibody Specificity
To test the specificity of the antiserum to the bullfrog ultimobranchial gland, we conducted Western blotting analysis of the extracts. In the extract of ultimo-branchial gland, the antiserum against PC1 detected a major band at 65 kD and a minor band at 59.5 kD (Figure 2), and the same result was obtained using the extract of the neurointermediate lobe as control. These bands were not detected when anti-PC1 was preabsorbed with the peptide used as the immunogen. No bands were seen in the ultimobranchial gland when anti-PC2 was incubated, although the antiserum against PC2 detected a band at 70 kD in the neurointermediate lobe (Figure 2), which disappeared in the absorption test.
RT-PCR Expression of Bullfrog PC1 and PC2 mRNAs in Ultimobranchial Gland
To investigate whether bullfrog PC1 or PC2 mRNA is expressed in the ultimobranchial gland, RT-PCR was performed using total RNA from the ultimobranchial gland, neurointermediate lobe, and skeletal muscle. Both PC1 and PC2 mRNAs were observed in the ultimobranchial glands and in the neurointermediate lobe (Figure 3). No PC1 and PC2 mRNAs were detected in skeletal muscle. This RT-PCR result was confirmed by Southern blotting analysis (data not shown).
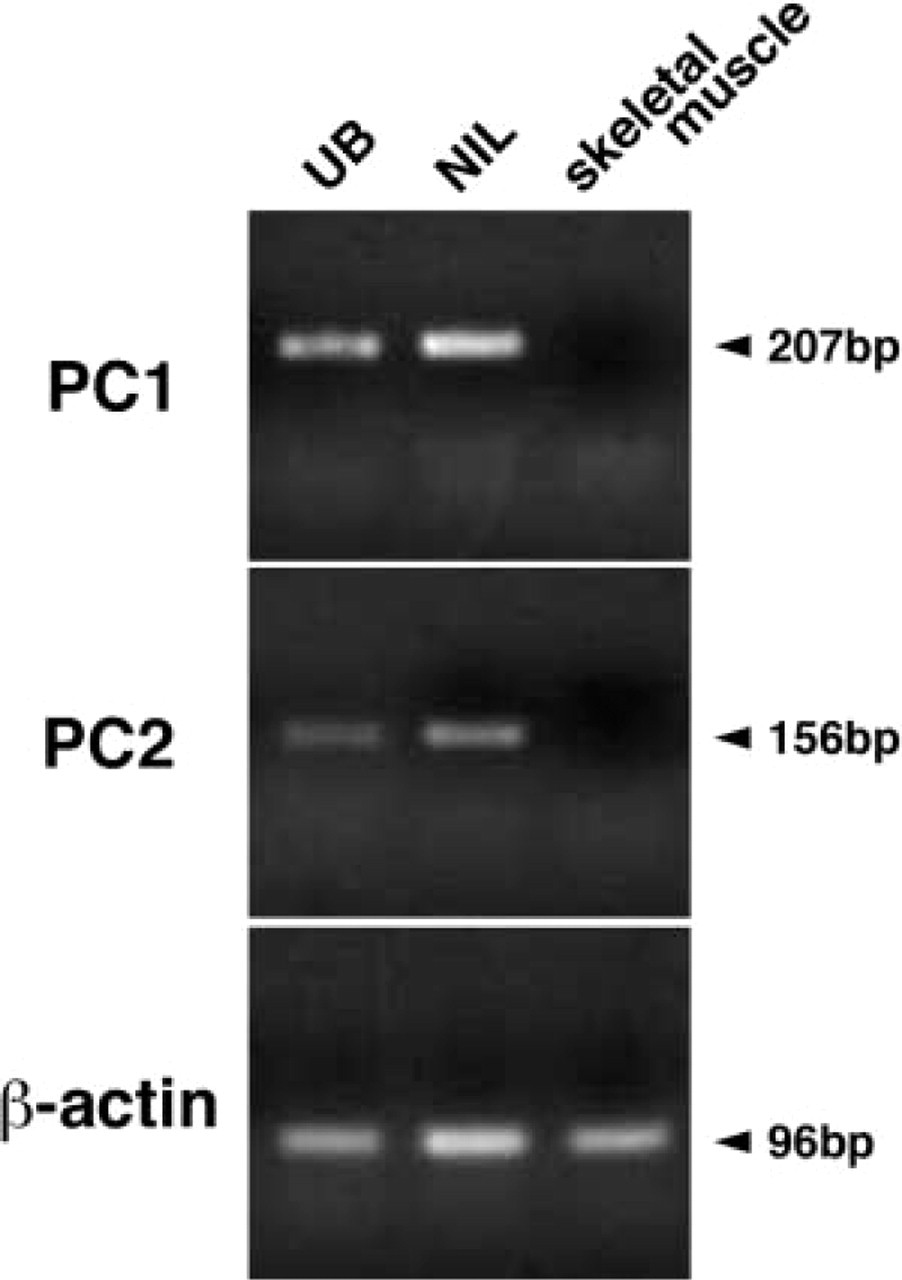
RT-PCR of PC1 and PC2 mRNAs in the ultimobranchial gland, the neurointermediate lobe, and muscle. RT-PCR products using primers as described in Materials and Methods were separated on a 2% agarose gel and stained with ethidium bromide.
Spatial Expression of PC1 mRNA in Calcitonin-producing Cells
To confirm the IHC results, we conducted ISH. PC1 mRNA-expressing cells were detectable in the follicle cells in the ultimobranchial gland (Figure 4a). No PC2 mRNA-expressing cells were seen in any cells of the gland (Figure 4b). The sense probe for PC1 mRNA showed no positive reaction (Figure 4c). To identify cells that express PC1 mRNA in the ultimobranchial gland, we applied fluorescence staining with rabbit anti-salmon calcitonin to the same section. Most PC1 mRNA-expressing cells were also reactive with anti-calcitonin (Figure 5).
Discussion
The present study, using IHC and ISH techniques, clearly demonstrates that calcitonin-producing cells in the ultimobranchial gland of bullfrog express one of two prohormone convertases, i.e., PC1. The antisera used in this study were developed according to the sequence corresponding to the portion of mouse PC1 and PC2 proteins as antigens. The amino acid sequence (442–459) of mouse PC1 used as the immunogen is identical to that of bullfrog PC1 except for one amino acid substitution at 459 (Lys in place of Arg in mouse sequence; DDBJ/EMBL/GenBank accession no. AB105175; Yaoi et al. 2003a). Similarly, the amino acid sequence (613–629) of mouse PC2 protein used as the immunogen is the same as that of bullfrog PC2 except for one amino acid substitution at 616 (Glu in place of Gln in the mouse sequence; DDBJ/EMBL/ GenBank accession no. AB105176; Yaoi et al. 2003a). Western blotting analysis of the extracts of bullfrog ultimobranchial gland using antiserum against PC1 showed one major band of 65 kD and a minor band of 59.5 kD, which are nearly consistent with those of mammalian PC1 proteins (Tanaka et al. 1996). The minor band may be a degraded form of PC1 protein because the molecular mass of 59.5 kD is lower than that of the active form (c. 65 kD). On the other hand, in Western blotting with the PC2 antiserum we did not detect any protein in the ultimobranchial gland, although this antiserum recognizes a specific band in the extract of the neurointermediate lobe. In the present study, the specificity of positive reaction in Western blotting was proved by the absorption test using the corresponding antigen peptides. The Western blotting results were also consistent with the present IHC results, indicating the presence of only PC1 protein in ultimobranchial calcitonin-producing cells. We also obtained similar results by ISH, which revealed PC1 mRNA in the ultimobranchial gland and the absence of PC2 mRNA. Moreover, we demonstrated that PC1 mRNA is expressed in the calcitoninproducing cells. In the present study we also showed the presence of PC1 in the CGRP-positive cells of the ultimobranchial gland, a situation similar to the previous study showing that, in the mouse respiratory tract, CGRP may be processed by PC1 alone (Kurabuchi and Tanaka 2002).
In the present RT-PCR study, we detected PC1 mRNA and PC2 mRNA in the ultimobranchial gland but we did not visualize PC2 mRNA using the ISH technique. Because RT-PCR allows one RNA to be amplified a billionfold, this technique may enable a very small amount of PC2 mRNA that is not detectable by ISH to be visualized. On the other hand, we did not detect PC2 protein by IHC staining or by Western blotting analysis. Consequently, we assumed that very little PC2 mRNA is translated.
It is known that alternative processing of RNA transcripts from the calcitonin gene results in the production of distinct mRNAs encoding calcitonin or CGRP (Amara et al. 1982). A sequence analysis of these mRNAs revealed that procalcitonin is cleaved at the Lys-Arg and Gly-Lys-Lys-Arg sites to produce calcitonin, CCP, and 83-amino-acid N-terminal peptide, whereas proCGRP is cleaved at two cleavage sites, Lys-Arg and Gly-Arg-Arg-Arg, to generate CGRP (Amara et al. 1980,1982). Tomita (2000) showed the presence of PC1 and PC2 in normal human C-cells and in medullary thyroid carcinomas. Similarly, our previous data indicated that thyroid C-cells in mouse express both PC1 and PC2, suggesting that procalcitonin is proteolytically cleaved either by PC2 alone or by PC1 together with PC2. On the other hand, the present study demonstrated co-expression of PC1 with calcitonin in the follicular cells of ultimobranchial glands, suggesting that procalcitonin might be processed by PC1 alone. This discrepancy may be due to the substrate specificity at the cleavage sites of procalcitonin between mammals and amphibians. However, the exact explanation remains to be clarified because the amino acid sequence of procalcitonin in the bullfrog has not yet been determined. Therefore, it was proved that these antisera are specific for bullfrog PC1 and PC2 proteins.
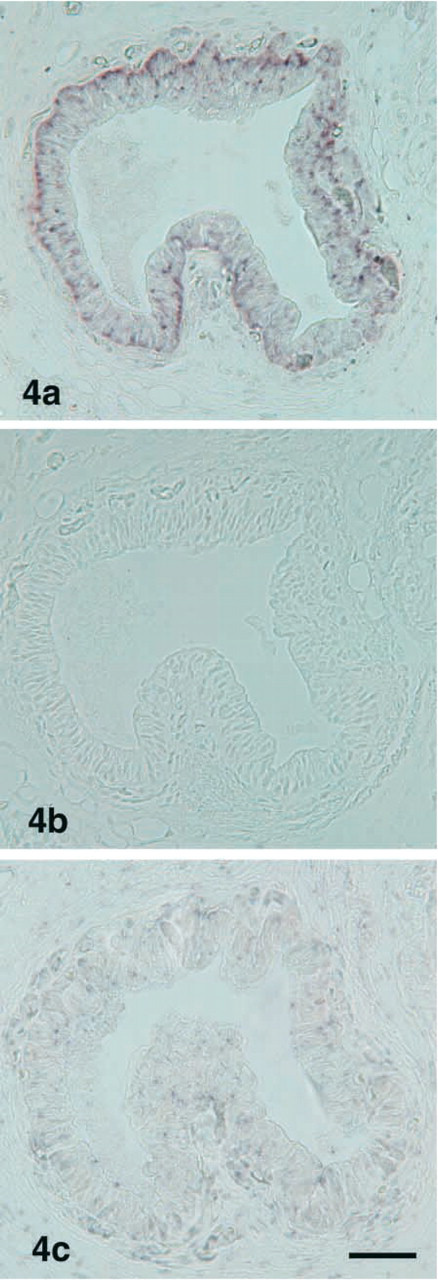
Light micrographs showing localization of PC1 mRNA and PC2 mRNA in the ultimobranchial gland. PC1 mRNA is visible in the follicular cells (
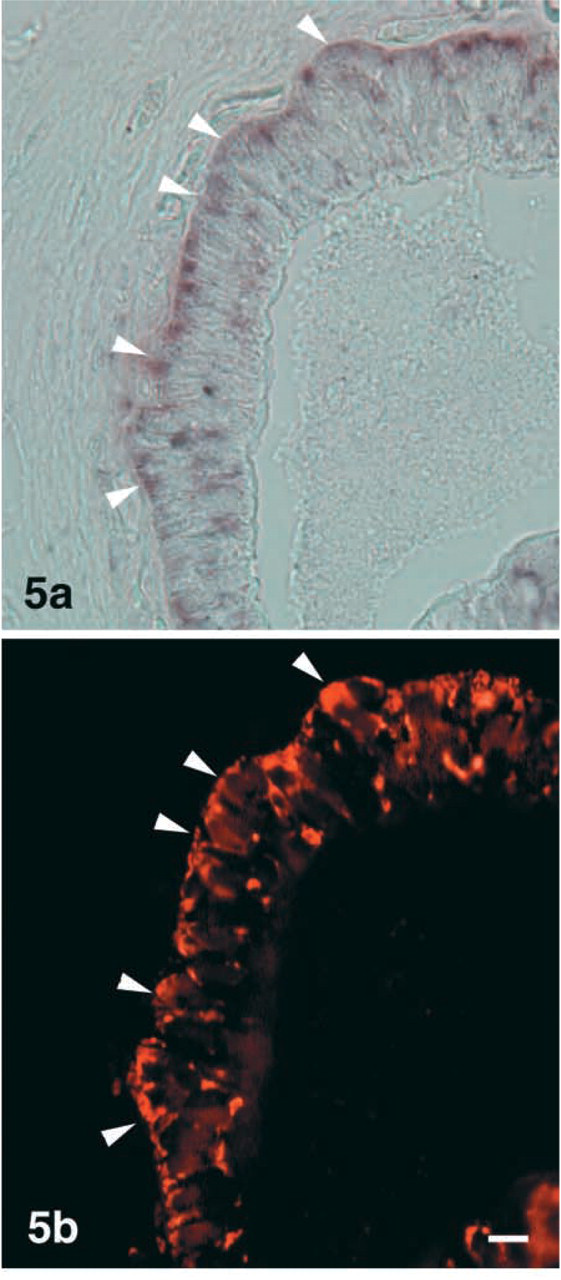
Light micrographs showing dual staining for PC1 mRNA (
Because there are no biochemical studies on the specificity of PC1 and PC2 in the cleavage of calcitonin, our ICC observation provides further argument for the substrate specificity of PC1 in the proteolytic cleavage of procalcitonin. In this connection, it is of interest that very high levels of serum procalcitionin is present in patients with severe systemic inflammation and sepsis, and this finding is used as a clinical marker for systemic infection responses (Assicot et al. 1993; Dandona et al. 1994; Russwurm et al. 1999). Although the cellular sources of the high levels of serum procalcitonin have not been fully identified, various cells, including blood mononuclear cells, are presumed to secrete the procalcitonin (Russwurm et al. 2001). Therefore, it is important to identify the precise cellular sites at which procalcitonin is synthesized during inflammatory conditions. The location may be found by determining which cells possess prohormone convertases, because procalcitonin should be processed to calcitonin in cells expressing PC1 and/or PC2.
Amphibians have a well-developed endolymphatic sac containing tiny crystals of calcium carbonate in the form of aragonites (Simkiss 1967). Frog calcitonin stimulates incorporation of serum calcium into the endolymphatic sac, thereby accelerating the formation of calcium carbonate crystals in the endolymphatic sac (Oguro et al. 1984; Srivastav and Rani 1989). We recently cloned cDNA encoding otoconin-22, a protein involved in the formation of calcium carbonate crystals in the endolymphatic sac, and then demonstrated that ultimobranchial calcitonin regulates otoconin-22 mRNA (Yaoi et al. 2003b).
These data suggest that proteolytic cleavage of procalcitonin to calcitonin is caused by PC1 in the calcitonin-producing cells of the bullfrog ultimobranchial gland, and that the calcitonin produced then stimulates expression of otoconin-22 mRNA, thereby forming calcium carbonate crystals in the endoplymphatic sac.
Footnotes
Acknowledgements
Supported in part by a Grant-in-Aid for Scientific Research from the Ministry of Education, Science, Sports, and Culture of Japan (ST).