
Abstract
Nerve growth factor (NGF) in mouse submandibular glands (SGs) is generated from a 35-kD precursor by proteolytic enzymes that have yet to be identified. Prohormone convertases (PCs) cleave the NGF precursor in vitro, and in this study we questioned whether PCs could process salivary NGF in vivo. mRNA coding for PC2 (but not PC1) was detected on Northern blots of SG mRNA and also by in situ hybridization within parasympathetic neurons of intralobular ganglia. Northern blot and in situ hybridization analyses also detect mRNA coding for furin. In SGs of male mice, furin mRNA levels are high at birth and remain high throughout development. In glands from female mice, levels decline during postnatal development and are lower in adults than in newborns. Immunocytochemistry detects furin immunoreactivity in pro-acinar and ductal cells of glands from newborn and pubescent mice. In glands of adults, furin immunoreactivity is detectable in acinar cells but highest levels are present in NGF-containing granular convoluted tubule cells. These data, taken together with those from previous studies, suggest that furin is a candidate processing enzyme for NGF in mouse submandibular glands.
Keywords
NGF is a homodimeric polypeptide made up of two 118 amino acid subunits. The monomer arises from the COOH terminus of a 390-amino-acid precursor (pre-pro-NGF) that is cleaved following an Arg-Ser-Lys-Arg sequence immediately preceding the NH2 terminus of the mature protein (Scott et al. 1983). In searching for the enzymes that process the NGF precursor, we and others have examined the mammalian subtilisin/kexin-like family of serine proteases called prohormone convertases (Seidah et al. 1996a; Bresnahan et al. 1990,1992). These calcium-dependent enzymes are structurally similar to the Kex2 endoprotease of yeast (for reviews see Seidah et al. 1994; van de Ven et al. 1993). In mammalian tissues, seven members of the family have been identified: furin, PACE4, and prohormone convertase (PC) 1, 2, 4, 5/6, and 7. PACE 4 and PC5 are present in some endocrine and non-endocrine cells, PC1 and PC2 have been identified in neurons and endocrine cells (Seidah et al. 1994; Schafer et al. 1993; Day et al. 1992), and PC4 is expressed exclusively in testicular germ cells (Seidah et al. 1992, 1994). Furin is ubiquitously expressed and is the primary convertase in cells that release proteins by constitutive secretion (Seidah et al. 1994,1996b; Day et al. 1993; van de Ven et al. 1993).
PC1 and PC2 usually cleave precursor molecules at solvent-accessible single or paired basic amino acids. Furin, which cleaves at the consensus site Arg-X-(Lys/Arg)-Arg, processes a number of precursors, including the pro-von Willebrand factor (Wise et al. 1990), complement pro-C3 (Misumi et al. 1991), and prosomatostatin (Galanopoulou et al. 1995). Bresnahan et al. (1990, 1992) were the first to show that furin can process mouse pro-NGF to NGF using an in vitro Vaccinia virus infection system, a result we have confirmed (Seidah et al. 1996a). PACE 4 and PC 5/6 also show NGF processing activity.
Furin has been localized to the constitutive secretory pathway through a membrane-spanning C-terminal domain associated with the trans-Golgi network (Misumi et al. 1991; Bresnahan et al. 1990). However, furin may also act at the cell surface because it contains a transmembrane-spanning segment within its C-terminus (Molloy et al. 1992,1994; Klimpel et al. 1992). A soluble form of furin can be released from cells by a proteolytic cleavage that liberates fragments of the protein (Vey et al. 1994; Vidricaire et al. 1993; Rehemtulla et al. 1992).
In this study we compared the developmental expression and cellular localization of NGF, PC1, PC2, and furin in mouse SGs. The purpose was to determine whether these convertases are candidates for processing pro-NGF in vivo. The data we obtained are consistent with the idea that furin, but not PC1 or PC2, could generate NGF from its precursor in mouse SGs.
Materials and Methods
Animals
Male and female Balb/c mice obtained from Charles River Breeding Laboratories (St-Constant, Quebec, Canada) were housed in small groups under standard lighting conditions with free access to water and food. Before surgery, animals were anesthetized by IP injections of sodium pentabarbitol (30 mg/kg).
Complementary RNA Probes
Plasmids for the prohormone convertases furin, PC1, and PC2 were prepared and used as described by Meyer et al. (1996). NGF cDNA (Scott et al. 1983) was inserted into the pGEM3Z plasmid, linearized, and used for synthesizing sense and anti-sense cRNA probes with SP6 and T7 polymerase. Fragments from cDNA clones for mNGF (960 BP), mfurin (1.1 KB), mPC1 (521 BP), and mPC2 (508
Northern Blot Analysis
Total RNA was extracted from mouse SGs using the TRIzol total RNA isolation reagent (Gibco; Grand Island, NY) according to manufacturer's instructions. Samples of RNA were processed as described previously (Day et al. 1992). Briefly, RNA (5 μg, heated for 10 min at 65C) was electro-phoresed on 1.2% agarose gels containing 20 mM HEPES (pH 7.8), 1 mM EDTA, and 6% formaldehyde. The gel was submerged in the same HEPES buffer and the samples were transferred from the gel to a nylon filter (Nytran) by capillary action. The filters were completely dried, and the RNA was fixed to the filters by long-wave UV irradiation. The filters were destained in 0.1 × salt sodium citrate (SSC) solution containing 1% SDS for 15 min (1 × SSC contains 0.15 M NaCl and 0.15 M sodium citrate), and prehybridized at 62C for 2 hr in 400 mM sodium phosphate buffer (pH 7.2) containing 5% SDS, 1 mM EDTA, 1 mg/ml BSA, and 50% formamide. Hybridization was performed with addition of the [32P]-UTP-labeled cRNA probe for 12–16 hr at 62C. Filters were washed in 0.1 × SSC, 0.1% SDS, and 1 mM EDTA at 75C for 2–3 hr and exposed to X-ray film with intensifying screens at −80C for appropriate intervals. Loading concentrations were standardized by monitoring ribosomal RNA with the methyl blue color stain and by hybridization with a probe specific for 18S ribosomal RNA.
In Situ Hybridization
In situ hybridization was performed using the protocol described by Schafer and Day (1995). Briefly, mice under terminal anesthesia were fixed by transcardiac perfusion with PBS followed by a buffered (pH 7.4) solution of 4% paraformaldehyde. Cryostat sections (12 μm) of SGs were postfixed for 20 min in 4% phosphate-buffered formaldehyde solution and washed twice in PBS, pH 7.4, for 10 min. Sections were treated with proteinase K (0.1 μg/ml in 0.1 M Tris-HC1, 50 mM EDTA, pH 8.0) for 10 min at 37C, washed in DEPC water, and transferred to 0.1 M triethan-olamine (pH 8.0) for 5 min at room temperature (RT). Sections were incubated in 0.1 M triethanolamine containing acetic anhydride (0.25% v/v) for 10 min at RT, rinsed in 2 × SSC, and dehydrated in graded ethanol (50–100%). Hybridization was carried out with [35S]-UTP-labeled cRNA probes diluted in hybridization buffer [75% formamide, 3 × SSC, 50 mM sodium phosphate (pH 7.4), 1 mM EDTA, 1 × Denhardt's solution, 0.2% (w/v) bovine serum albumin, 0.02% (w/v) Ficoll 400, and 0.02% (w/v) polyvinylpyrrolidone], yeast tRNA (0.1 mg/ml), and sheared salmon sperm DNA (0.1 mg/ml) with 10 mM DTT. Hybridization was performed for 16 hr at 55C. The sections were incubated in 2 × SSC for 10 min, treated with RNAse A [40 μg/ml in 10 mM Tris-HC1 (pH 7.6), 0.5 M NaC1, 1 mM EDTA] for 30 min at 37C, followed by successive washes in 2 X, 1 X, and 0.5 × SSC for 10 min each at RT and 0.1 × SSC for 1 hr at 55C. Sections were dehydrated in graded ethanol, dipped in Kodak NTB2 nuclear emulsion (diluted 1:1 in water), and stored at 4C for 3–4 weeks. Slides were developed in Kodak D19 for 2 min and fixed in 30% sodium thiosulfate for 5 min. After washing in H2O and serial dehydration, sections were coverslip-mounted with Permount. Some sections were counterstained with cresyl violet before mounting. The specificity of the probes was confirmed by substituting anti-sense probes with sense probes or by pretreating tissue sections with RNAse A (0.1 mg/ml in 2 × SSC for 1 hr at 37C) before hybridization.
Immunocytochemistry
Rabbit polyclonal antiserum raised against mouse 2.5S NGF (Murphy et al. 1989), and a furin-specific monoclonal antibody, MON-148 (van Duijnhoven et al. 1992; a gift from Dr. W.J. van de Ven), were used in these studies. SGs were removed from mice perfused as above and incubated overnight in 30% sucrose at 4C. Cryostat sections (12 μm) were initially fixed with 4% paraformaldehyde/2.5% DMSO for 30 min and washed three times with TBS. The sections were permeabilized with TBS/0.2% TX-100 for 10 min, washed with TBS, and blocked with TBS/0.2% TX-100/1% goat serum for 1 hr, followed by washing with TBS and overnight incubation at 4C with NGF (1:1000) or furin (1:100) antibody. After washes with TBS/0.01% TX-100, sections were incubated for 1 hr with biotinylated goat anti-rabbit IgG (for NGF) or mouse IgG (for furin), followed by washes and 1-hr treatment with avidin-alkaline phosphatase (Sigma; St Louis, MO). Sections were washed and treated for 15 min with NBT/X-phosphate/levamisol (2 mM) in TBS (pH 9.5) containing MgCl2 (50 mM). For localizing NGF immunoreactivity, we stopped the reaction with 10 mM Tris/1 mM EDTA (pH 8) after approximately 5 min. Sections treated with furin antibody were allowed to develop for up to 20 min for optimal staining. After dehydration in sequential alcohols, sections were mounted for light microscopy. Furin antibody specificity was verified by blocking staining with excess amounts of recombinant furin fragment BTMD (Before TransMembrane Domain), prepared as described (Jean et al. 1993). The construct coding for this fragment, made as described by Leduc et al. (1992), is contained within the Bluescript (BSKSII[+]) vector, and was generously provided by Dr. Gary Thomas.
Detection of Furin by Western Blotting
Golgi fractions of adult male mouse SGs were isolated using a methodology based on the isolation of rat liver Golgi fractions (Bergeron et al. 1982). SGs were homogenized (10% homogenate) using a Kinematica CH-6010 polytron (setting of 6) with two 10-sec bursts at 4C in 0.25 M sucrose in 4 mM imidazole buffer, pH 7.4. The homogenizing solution and all buffers contained the protease inhibitors PMSF (1 mM), aprotinin (200 KU/ml), leupeptin (10 μg/ml), and pep-statin (10 μM). After centrifugation at 400 × g for 10 min, the supernatant was retained and the pellet was re-homogenized in the same volume of buffer using a single 10-sec polytron burst at a setting of 6. After centrifugation at 400 × g for 10 min, the two supernatants were combined and centrifuged at 200,000 × g for 45 min. The pellet was resuspended with a loose Dounce homogenizer in 0.25 M sucrose and 4 mM imidazole buffer, the sucrose concentration adjusted to 1.15 M, and overlaid with 0.95 M sucrose-imidazole and 0.4 M sucrose-imidazole. After centrifugation (90 min at 200,000 × g in a Beckman SW 40 rotor), Golgi fractions were collected at the 0.4/0.95 M sucrose interface. Galactosyl transferase assays with ovomucoid as exogenous acceptor were performed as described previously (Bergeron et al. 1982). For Western blotting, aliquots of Golgi fractions were separated on 7–15% gradient gels under denaturing and reducing conditions. Proteins were transferred to Trans-Blot Supported Nitrocellulose Membrane (Bio-Rad; Richmond, CA), and probed overnight at 4C with the furin monoclonal antibody MON-148 (1:200). After several washes, the primary antibody was visualized using a horseradish peroxidase-coupled goat anti-mouse antibody (Jackson ImmunoResearch Laboratories; West Grove, PA), followed by peroxidase-catalyzed chemiluminescence (enhanced chemiluminescence; Amersham, Arlington Heights, IL) according to the manufacturer's instructions.
Results
Northern Blot Analyses
Northern blot data confirm that SGs of adult male mice contain higher levels of mRNA coding for NGF than glands of 21-day males or adult females (Figure 1A). The differences are due to circulating testosterone which promotes the production of NGF within GCT cells, as reviewed previously (Gresik and Mac-Rae 1975). Figure 1B shows that mRNA coding for furin and PC2 is also detectable in SGs. A single mRNA transcript coding for furin is evident in RNA from SGs of male and female mice and is of similar size (4.4 KB) to that reported previously (Seidah et al. 1994; Day et al. 1993; Roebroek et al. 1986). A 2.8-KB mRNA transcript coding for PC2 is also present in SGs of male and female mice.
The developmental expression of furin and PC2 mRNA differs (Figure 1B). Levels of furin mRNA in SGs of postnatal Day 1 mice are approximately equal in males and females. Levels decrease in females between postnatal Days 1 and 21, and in the adult are lower than in the newborn. In males, furin mRNA levels remain stable throughout postnatal development and are as high in adult glands as in newborns. Levels of PC2 mRNA are comparable in glands from male and female mice on Day 1 but decline with increasing age in both sexes. mRNA coding for PC1 is not detectable in SGs of mice of either sex at any developmental stage (data not shown).
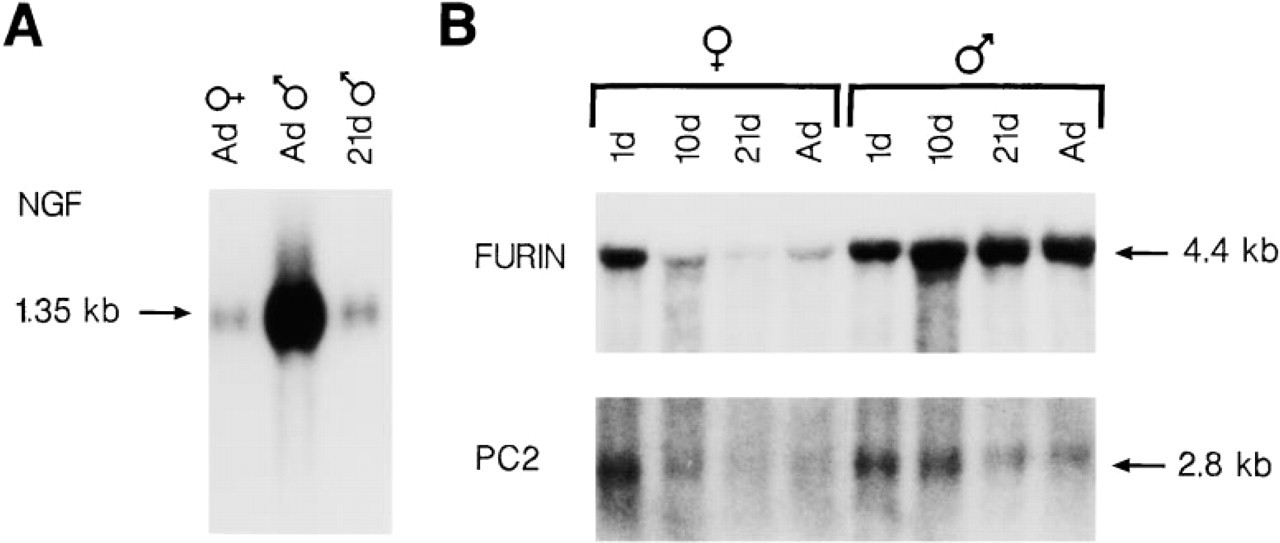
Northern blot analyses of mRNA from male and female mouse SGs.
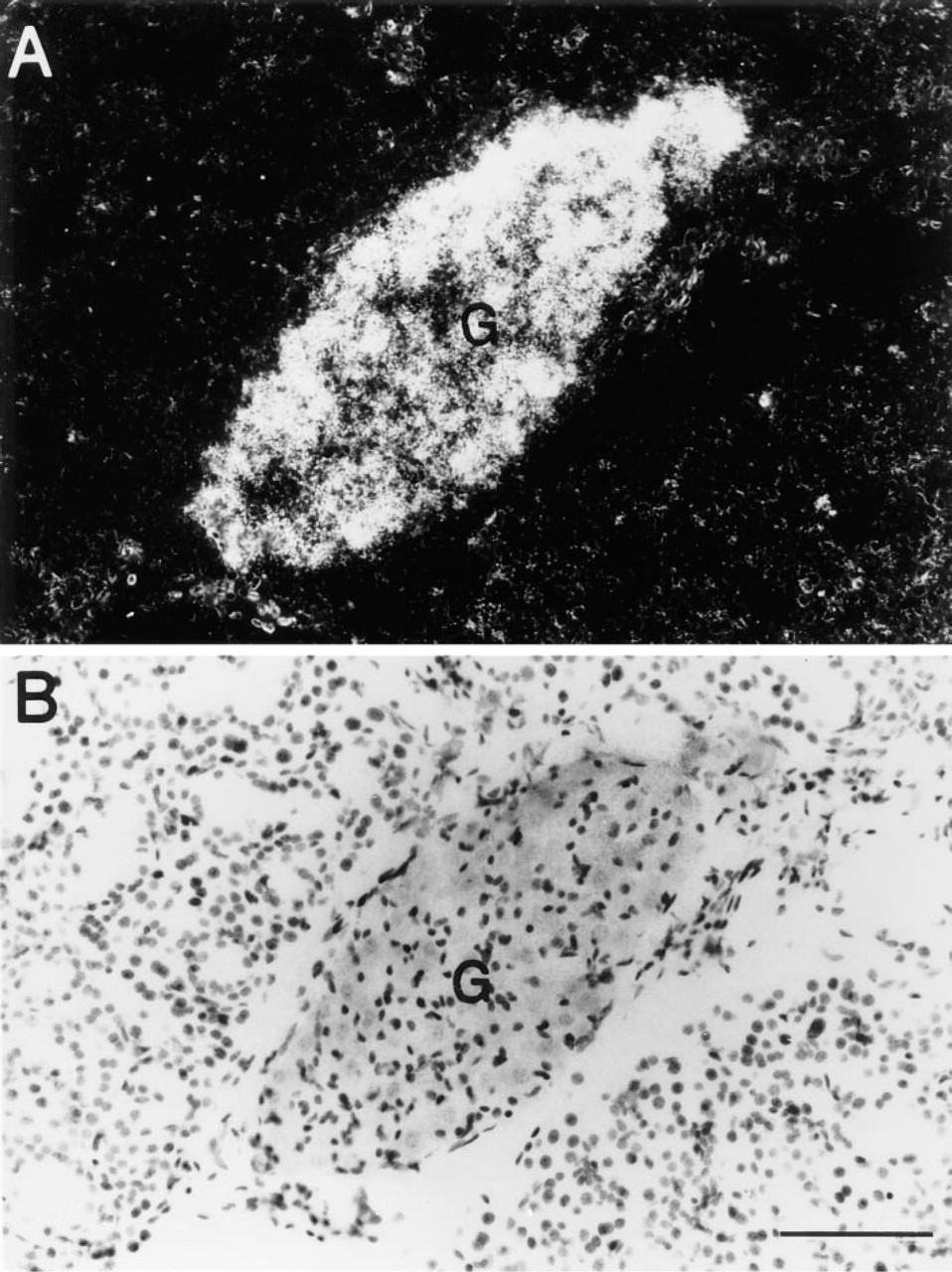
Morphological Studies
The postnatal development of mouse SGs has been well described (Chen et al. 1995; Durban et al. 1994; Denny et al. 1993; Gresik 1980). At birth, the gland is composed of terminal tubule and pro-acinar cells connected to intralobular ducts. Terminal tubule cells, which occupy only a small portion of the gland's volume, are difficult to identify without toluidine blue staining and are not clearly visible in frozen sections. However, tubule cells, together with pro-acinar cells, differentiate into mature acinar cells by postnatal Day 10. Duct cells, which in newborn mice account for approximately 60% of the gland's volume (Chen et al. 1995), undergo a more complicated course of differentiation. Duct cells in newborns give rise to the intercalated and striated duct cells of the adult, both of which are evident by postnatal Day 7 (Gresik and Mac-Rae 1975). Androgen-dependent GCT cells, which begin to develop around Day 15, can be identified by a nucleus located within the basal cytoplasm and, in the adult, by large secretory granules within the apical cytoplasm. GCT cells develop until 6 months of age.
In adult males, GCT cells account for approximately 57% of the SG's volume, acinar cells 23% of volume, and striated and intercalated duct cells together approximately 1% of gland volume. In contrast, GCT cells in glands of adult female mice account for only 19% of SG volume. The remainder of the gland is made up of acinar cells (53% of gland volume) and striated and intercalated duct cells (6% of gland volume) (Murphy et al. 1981).
In Situ Hybridization
Intense hybridization for PC2 mRNA is evident in small clusters of neurons within parasympathetic ganglia located in intralobular regions of SGs from adult mice (Figure 2) (Ng et al. 1992a,b). No hybridization for PC2 is evident in any other cell type.
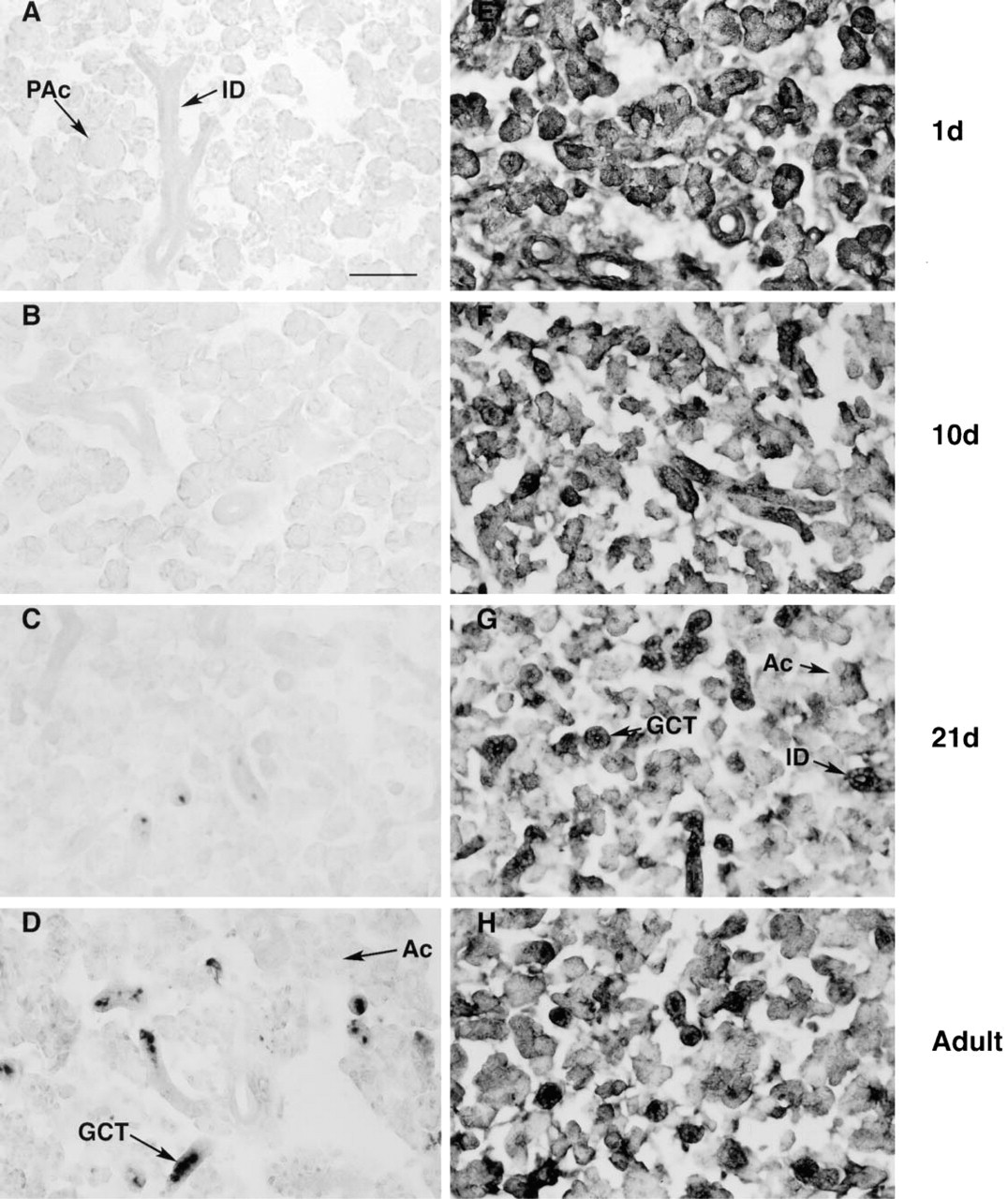
Furin mRNA is widespread. In newborn (Figure 3A) and postnatal Day 10 male mice (Figure 3B), furin mRNA is distributed within the pro-acinar cells and within cells associated with the developing duct system. In 21-day-old and adult animals (Figures 3C and 3D), labeling is evident in duct cells as well as in GCT cells and acinar cells.
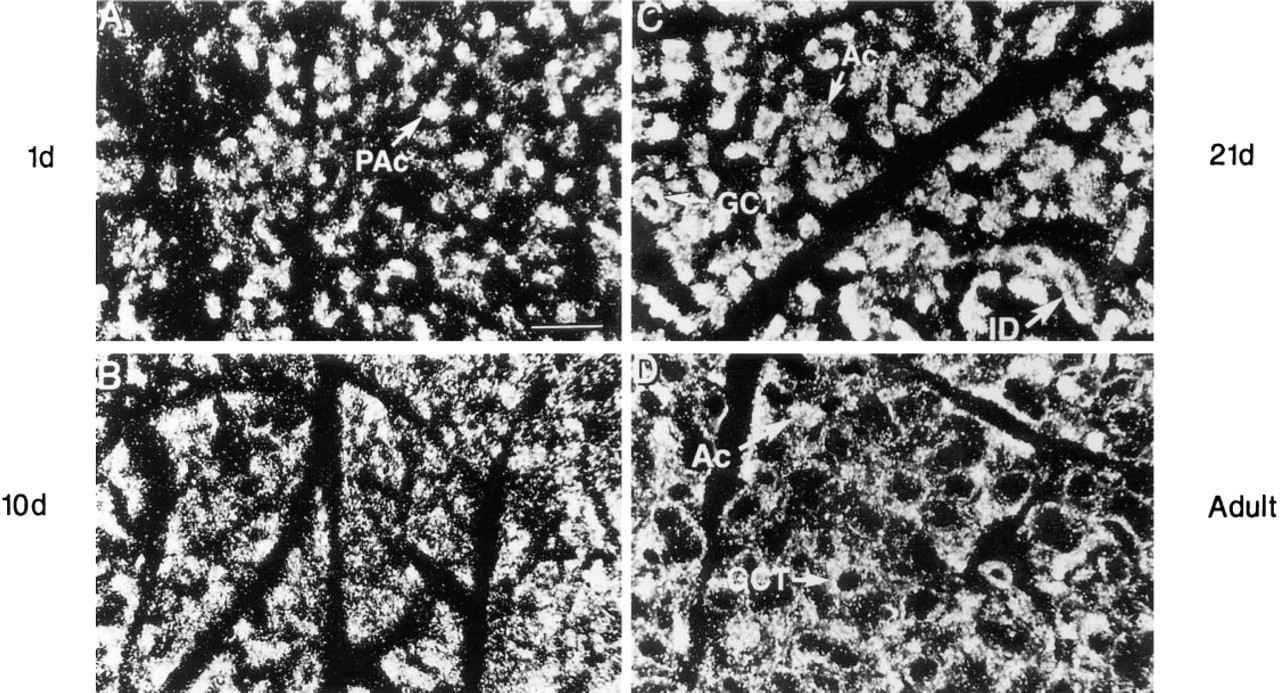
In situ hybridization analyses. Darkfield photomicrographs of SGs from developing male mice showing the expression of furin mRNA. In glands from newborn
Immunocytochemistry
Figures 4 and 5 compare the developmental appearance of NGF and furin in SGs of male (Figure 4) and female (Figure 5) mice. NGF immunoreactivity is absent from glands of newborn and 10-day mice of both sexes (Figures 4A, 4B, 5A, and 5B) but is evident in GCT cells from glands of 21-day and adult males and females (Figures 4C, 4D, 5C, and 5D). Higher levels of NGF immunoreactivity are present in glands of male mice owing to their content of larger and more numerous GCT cells.
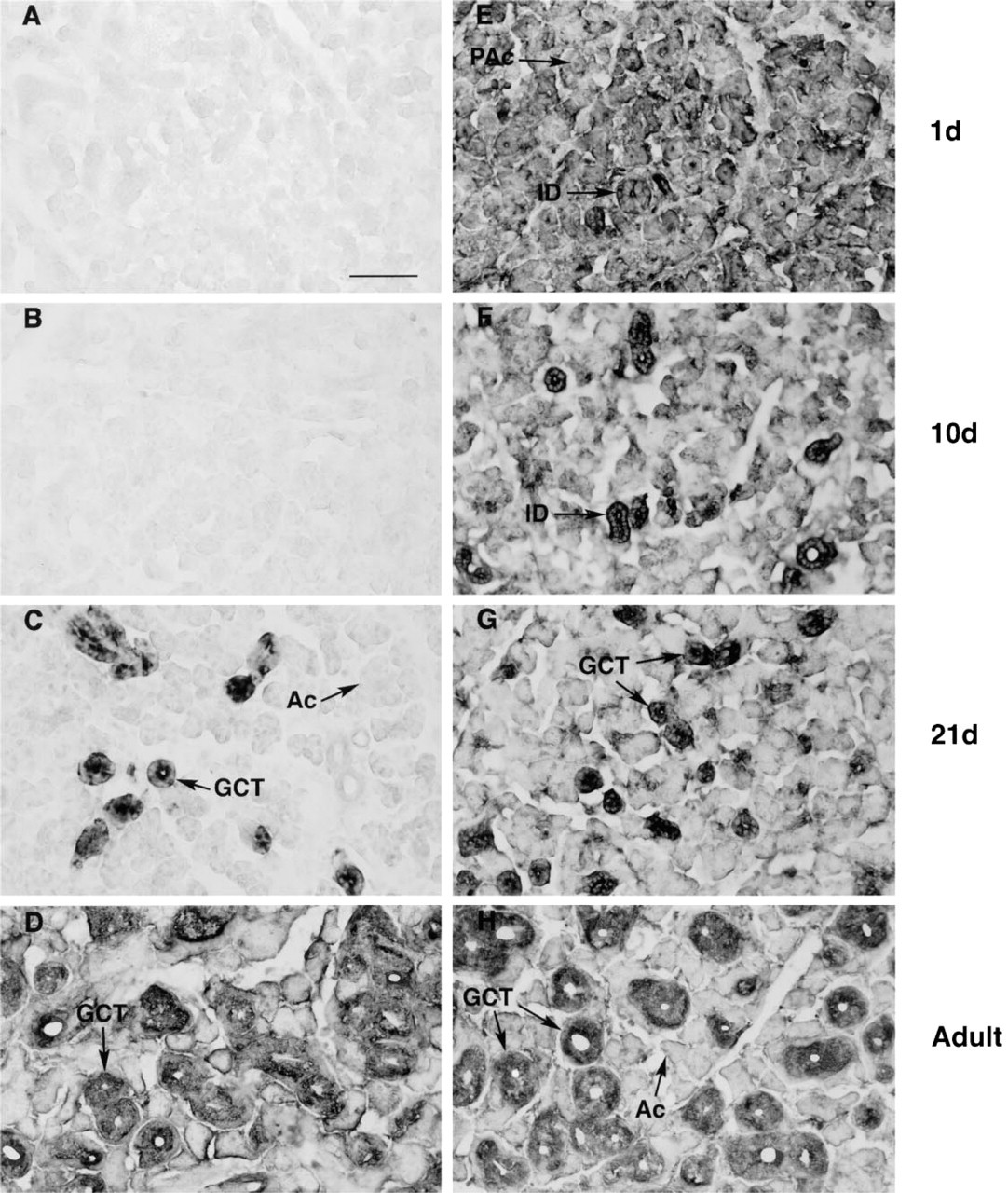
Immunocytochemical detection of NGF
Immunocytochemical localization of NGF
Unlike NGF, furin immunoreactivity is evident in SGs of both male (Figure 4) and female (Figure 5) mice of all ages tested. In postnatal Day 1 animals (Figures 4E and 5E), immunoreactivity is evident in pro-acinar and duct cells. In SGs from Day 10 animals, furin immunoreactivity remains widespread but is especially concentrated within ductule structures (Figures 4F and 5F). In 21-day and adult animals of both sexes, furin staining is visible within acinar cells (Figures 4G, 4H, 5G, and 5H), but highest levels are evident in duct cells, including GCTs. In some GCT cells, furin immunoreactivity is evident in the apical cell cytoplasm, in a position similar to that of NGF (Gresik and MacRae 1975).
Immunoblotting
To confirm that furin protein is present within SGs, we carried out Western blot analysis using the furin-specific monoclonal antibody. Furin immunoreactivity is not detected in crude SG homogenates (Figure 6, Lane 3) but is evident in isolated Golgi fractions, migrating with the expected molecular mass of 100 kD (Figure 6, Lane 4). In these samples, the marker enzyme galactosyl transferase had been concentrated 20-fold compared to the starting homogenate (data not shown). Furin immunoreactivity is not evident in samples blotted with serum not containing the monoclonal antibody (Figure 6, Lanes 1 and 2).
Discussion
The data in this study show that mouse SGs contain PC2 and furin, two members of the convertase family of pro-hormone processing enzymes. In SG from adult mice, furin and NGF co-localize within GCT cells.
mRNA coding for PC2 was detected by Northern blot analyses (Figure 1) in SGs of newborn mice of both sexes; levels were highest in the newborn and declined with increasing age, a result that was explained by in situ hybridization results. PC2 mRNA is present within postmitotic parasympathetic neurons. Therefore, it is not surprising that PC2 mRNA levels in neurons decrease in proportion to total gland mRNA because other cells in the developing SG are dividing, enlarging, and differentiating. Previous studies have localized PC2 in brain neurons (Day et al. 1992) and in endocrine and neuroendocrine cells (Seidah et al. 1990, 1991; Smeekens and Steiner 1990). PC1 mRNA, which has been identified previously in neurons (Seidah et al. 1994), was not detected in SG by either Northern blot or in situ hybridization analyses.
Because PC1 and PC2 are not present in the same cells as NGF, neither enzyme is a candidate for processing salivary pro-NGF. This result is consistent with in vitro data showing that neither PC1 nor PC2 effectively processes pro-NGF (Seidah et al. 1996a). Furthermore, PC2 requires the environment of secretory granules as well as the neuroendocrine peptide 7B2 for maximal activity (Seidah et al. 1994), whereas pro-NGF appears to be activated within the trans-Golgi network (Seidah et al. 1996a).
Furin is widely distributed within SGs of female and male mice throughout development. In females, furin mRNA levels are highest at birth (Figure 1B), and furin immunoreactivity is evident in pro-acinar and ductule cells (Figure 5). Whether terminal tubules also contain furin is unclear from our studies because these cells are difficult to identify in our sections. As the animals age, levels of furin mRNA decline but furin immunoreactivity remains detectable in acinar cells and ductule cells. In the adult, high levels of furin immunoreactivity were present in GCT cells.
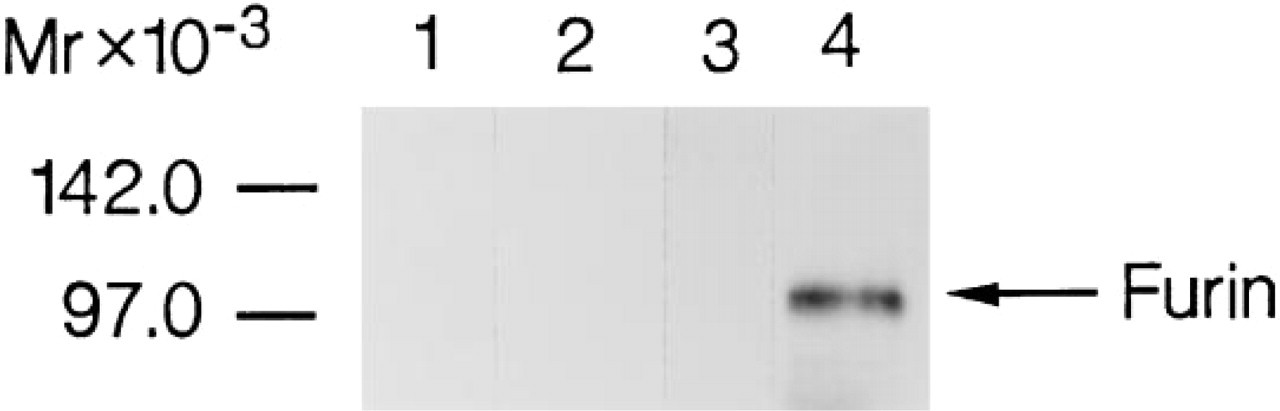
Immunoblot analysis of furin in adult male mouse SGs. Aliquots of SG homogenates and Golgi fractions isolated as described in Materials and Methods were subjected to SDS-PAGE (100 μg total protein in each lane) and immunoblot replicas were developed with a monoclonal antibody to furin. Lanes 1 and 3 contain samples of salivary gland extracts; Lanes 2 and 4 contain Golgi fractions of SG enriched 20-fold in the marker enzyme galactosyl transferase. Samples in Lanes 1 and 2 were treated with a solution containing goat serum with no monoclonal antibody; samples in Lanes 3 and 4 were treated with an identical solution containing the monoclonal antibody (MON-148) to furin.
In SGs from male mice, the cellular distribution of furin is similar to that in females but, unlike females, levels of furin mRNA remain high throughout development. These differences probably arise from furin's production in GCT cells, which are larger and more numerous in glands of males. Co-localization of NGF and furin in GCT cells is consistent with experimental evidence showing that furin effectively processes the NGF precursor (Seidah et al. 1996a; Bresnahan et al. 1990) and is an important requirement for linking the proteins physiologically.
Testosterone does not appear to directly regulate SG production of furin, at least during the early stages of development. Furin mRNA and protein are abundant in SGs of postnatal Day 1 animals, well before pubescent surges in circulating testosterone promote GCT cell development and NGF production. In glands from adult mice, furin mRNA and protein are also detectable in acinar cells and ductule cells, which are not regulated by testosterone. Therefore, furin must carry out additional functions within the SG that are independent of growth factor processing. Testosterone could regulate furin production in GCT cells of pubescent and adult mice through its effects on GCT cells.
Furin has been found in cells that release proteins by constitutive, nonregulated secretory mechanisms. Therefore, it was somewhat surprising to find such high levels of furin mRNA and protein in GCT cells. GCT cells store NGF and other proteins in cellular vesicles that are released by regulated secretory mechanisms (Murphy et al. 1977; Wallace and Partlow 1976). In some GCT cells, furin immunoreactivity was evident in the apical cell cytoplasm, in a position identical to that of NGF, suggesting that furin may be packaged within secretory vesicles, perhaps for release into saliva. Thus far we have not been able to detect furin in SG saliva using immunoblot methods that easily detect salivary gland NGF (data not shown). Furin could be retained within GCT cells through a COOH-terminal hydrophobic domain anchoring the protein to the secretory vesicle membrane. It could also be secreted with its substrate(s) through an autocatalytic mechanism, as suggested previously (Vey et al. 1994; Vidricaire et al. 1993).
Furin immunoreactivity was not detected on Western blots of SG homogenates, but was detected in isolated Golgi preparations containing concentrated levels of processed proteins. In contrast, furin immunoreactivity was readily detected by immunocytochemistry. Differences in detection sensitivity may be due to the furin monoclonal antibody reacting more effectively with native furin than with denatured and reduced forms of the protein. Furthermore, we needed to develop tissue sections approximately five times longer to visualize furin by immunocytochemistry than to visualize NGF, which may reflect differences in the affinities of the two antibodies or relative concentrations of furin and NGF within SGs.
Other proteolytic enzymes that can process NGF have been detected within SGs. The first was identified as a component of 7S NGF, a non-covalently linked protein complex in which mature NGF (called β NGF) is associated with two arginine-specific esterases, the α- and γ-subunits (Varon et al. 1968). The γ- but not the α-subunit can generate mature NGF from its precursor, but only when used at high stoichiometric levels. For that reason, the physiological relevance of the γ-subunit has been questioned (Jongstra-Bilen et al. 1989; Edwards et al. 1988; Saboori and Young 1986). Furthermore, the α- and γ-subunits have not been detected in other NGF-producing cells and tissues (Murphy et al. 1986; Pantazis 1983). A second endopeptidase has been identified that removes the amino terminal octapeptide of mature NGF, but it does not act at the cleavage site necessary for processing mature NGF from its precursor (Fahnestock et al. 1991).
Our observation that furin co-localizes with NGF in GCT cells of SGs, taken together with the studies of Bresnahan et al. (1990) and Seidah et al. (1996a) showing that furin effectively cleaves the NGF precursor, makes a strong case for furin being the physiological protease that activates pro-NGF in SGs. However, that physiological correlation can be made unequivocally only under experimental conditions in which the processing of salivary NGF is monitored under conditions in which furin activity is reduced or ablated, such as in furin knock-out mice.
Footnotes
Acknowledgements
Supported by Medical Research Council of Canada Program Grant MRC-PG11473 and by the Canadian Network of Centers of Excellence (NCE) on Neural Regeneration and Functional Recovery (to RAM). SP was supported by a NCE fellowship and RD is a fellow of the Fonds de la Recherche en Santé du Québec.
We thank Jacynthe Laliberté, Xue-Wen Yuan, and Ali Fazel for excellent technical assistance. We also thank Dr W.J. van de Ven for providing us with the furin antibody and Dr Gary Thomas for the vector coding for the furin fragment.