
Abstract
We examined the expression and localization of the prohormone convertases, PC1 and PC2, in the anterior pituitary cells of developing rats by a double staining procedure using in situ RT-PCR and an immunofluorescence technique. In the adult, both PC1 mRNA and PC2 mRNA were expressed in corticotrophs, gonadotrophs, thyrotrophs, and mammotrophs. These cells, except for corticotrophs, had previously been considered to be ones in which proprotein processing does not take place, but both PC1 and PC2 may be necessary to process other proteins, such as granin family proteins, having proteolytic cleavage sites and located in secretory granules of the above trophs. In addition, no PC1 or PC2 mRNA was expressed in somatotrophs, which is consistent with the fact that somatotrophs do not contain these granins. In addition, 7B2 mRNA was expressed in these PC2-positive trophs, suggesting that there is a functional relationship between PC2 and 7B2 proteins. We found that α-MSH was expressed in the corticotrophs of the postnatal rat and that the number of α-MSH-immunopositive corticotrophs decreased as development proceeded. Because the changes in the pattern of POMC processing are considered to depend on the relative expression levels of PC1 and PC2, PC1 and PC2 mRNAs were examined in corticotrophs during postnatal development. We found a decrease in the number of PC2 mRNA-positive cells, which coincided with one in the number of α-MSH-immunopositive corticotrophs, as postnatal development proceeded. Our present data demonstrate that the α-MSH production varies directly in accordance with the expression of PC2. We also discuss the possible significance of α-MSH production during the postnatal period.
Adrenocorticotropin (ACTH)-related peptides in corticotrophs in the pars distalis, and α-melanocyte-stimulating hormone (α-MSH)-related peptides and β-endorphin in melanotrophs in the pars intermedia, are produced posttranslationally by intracellular proteolytic cleavage of the large precursor molecule proopiomelanocortin (POMC). Nevertheless, the processing of POMC differs between the two lobes. In the pars distalis corticotrophs, ACTH, β-lipotropic hormone (β-LPH), and a 16-K peptide are the major end products, whereas in the pars intermedia melanotrophs ACTH is processed further into α-MSH and corticotropin-like intermediate peptide (CLIP), and β-LPH is processed almost completely into β-endorphin (Eipper and Mains 1980; Rosa et al. 1980; Chretien et al. 1989). In situ hybridization (ISH) studies showed that both PC1 and PC2 are present in the pars intermedia and that PC1 is expressed in the pars distalis (Seidah et al. 1991; Day et al. 1992). Consequently, tissue-specific processing of POMC is considered to proceed by the differential distribution of PC1 and PC2 in these tissues. This hypothesis was confirmed by the results of cell transfection studies. Introduction of antisense PC1 cRNA into AtT-20 cells, a mouse corticotroph tumor cell line, inhibits the production of ACTH. On the other hand, α-MSH was cleaved from ACTH when the cells were transfected with the PC2 gene (Zhou et al. 1993).
7B2 is a member of the granin family of acidic, neuroendocrine-specific secretory granule proteins that include chromogranin A and secretogranin II (Huttner et al. 1991; Mbikay et al. 2001). The 7B2 protein participates as a chaperone in the regulation of PC2 activation and to control the timing for activating the convertase (Martens et al. 1994). Consequently, identification of 7B2 in the PC2-expressing cells is important to elucidate the functional relationship between PC2 and 7B2 proteins.
Light micrographs showing localization of PC1 mRNA (
It is well known that the pattern of proteolytic processing of POMC in corticotrophs of the pars distalis shows a characteristic change during the postnatal period. In newborn rats, corticotrophs display a processing pattern similar to that of the melanotrophs, but as development proceeds the processing pattern in the corticotrophs changes to the adult type, with no ability to cleave POMC into α-MSH (Schafer et al. 1983; Allen et al. 1984; Sato and Mains 1985, 1986, 1988; Noel and Mains 1991). A few studies using ISH showed the expression of PC1 and PC2 in the pars distalis and pars intermedia, but these studies were unable to fully identify the cell types expressing these endoproteases because the resolution power of the ISH with radioisotopes was low (Day et al. 1992; Dong and Day 2002).
In the present study we identified cells expressing PC1, PC2, and 7B2 in adult rats by a double staining procedure using in situ RT-PCR and an immunofluorescence technique. Then we examined the expression patterns of PC1 and PC2 mRNAs during the postnatal period. We found that a decrease occurred in the expression level of PC2 mRNA, which was concomitant with a decrease in the number of α-MSH-positive corticotrophs as postnatal development proceeded.
In situ amplification of products by the second PCR. In the plasmid DNA (
Light micrographs showing localization of PC1 and PC2 mRNAs in corticotrophs of 8-week-old rats. PC1 mRNA (
Materials and Methods
Animals
Normal adult rats of the Wistar strain were housed in a temperature-controlled room (22 ± 2C) with automatically controlled lighting (lights on 0600–1800 hr daily) and were fed food and water ad libitum. The day after birth of the pups was denoted day 1 of postnatal life. The male postnatal rats (0.5 days, 1, 2, 3, 4, and 8 weeks) were decapitated after ether anesthesia and their pituitary glands were quickly removed and used for histological examination. All animal experiments were conducted in compliance with the Guide for Care and Use of Laboratory Animals established by Shizuoka University.
Light micrographs showing localization of PC1 and PC2 mRNAs in gonadotrophs of 8-week-old rats. PC1 mRNA (
In Situ RT-PCR
Animals were perfused through the heart with 4% paraformaldehyde (PFA) in 0.1 M phosphate buffer, pH 7.4. Their pituitary glands were excised and further fixed by immersion in the same fixative for 2 days. After fixation the tissues were dehydrated through a graded ethanol series, cleaned in methyl benzoate-celloidin, and embedded in Paraplast. Sections were cut at 4-μm thickness and mounted on silane-coated slides. The sections were pretreated with 1 U/ml DNase I (Takara; Kyoto, Japan) in 1 × DNase buffer containing 2 U/ml RNase inhibitor (Toyobo; Osaka, Japan) at 37C for 2 hr. After the DNase treatment they were washed with RNase-free PBS (0.01 M sodium phosphate buffer and 0.14 M NaCl, pH 7.5) and Mill-Q twice for 1 min each. One-step in situ RT-PCR was performed with the rTth enzyme, which combines cDNA synthesis and PCR amplification in a single reaction mixture on a thermal cycler (PCR Express, Thermo Hybaid; ASTEC, Fukuoka, Japan). The RT-PCR kit containing rTth DNA polymerase (Toyobo) was used and the reaction mixture (50 μl total per section) was prepared as the following solution: 10 μl of 5 × reaction buffer, 6 μl of 2.5 mM dNTPs, 1 μl of 10 U/μl RNase inhibitor, 23 μl of Milli-Q, 5 μl of 25 mM Mn(OAc)2, 1.2 μl of 25 pmol/ml each primer, 0.6 μl of 1 mM digoxigenin-11-dUTP (Roche Molecular Biochemicals; Meylan, France), and 2 μl of 2.5 U/μl rTth DNA polymerase. The sequences of sense (primer 1) and antisense (primer 2) primers were as follows: PC1 primer 1, 5′-AATCCTGTAGGCACCTGGAC-3′; PC1 primer 2, 5′-GGAGTTTTTGGGTACCAGGA-3′; PC2 primer 1, 5′-GAGAGGAGACCTGAACATCA-3′, PC2 primer 2, 5′-GCAAGCCCTTCTGTGGTGCA-3′. The cDNA was synthesized at 60C for 30 min. The procedure of PCR amplification was an initial denaturation step of 94C for 3 min followed by 25 reaction cycles of extension (50C, 60 sec) and denaturation (94C, 30 sec). Finally, an additional extension reaction was performed at 50C for 5 min. The specimens were fixed with a mixture of 4% PFA and 0.1% glutaraldehyde in Milli-Q for 20 min at 4C. After fixation, they were washed twice in 0.1 × SSC (standard saline citrate) for 20 min at 45C and once in washing buffer for 5 min. After a blocking step, the sections were incubated with alkaline phosphatase-conjugated sheep anti-DIG Fab antibody (Roche) for 15 hr. The label was detected with nitroblue tetrazolium chloride and 5-bromo-4-chloro-3-indolylphosphate (Roche) by incubating for the proper time. To check the specificity of the staining, we carried out in situ RT-PCR by using a reaction solution lacking the rTth polymerase.
As a control, the plasmid DNAs for rat PC1 and PC2 were used. PCR amplification for 40 cycles with specific primers was performed for pBK-CMV vectors containing a 1639-bp fragment (1020–2659 bp) of Wistar rat PC1 cDNA or a 1743-bp fragment (703–2446 bp) of the rat PC2 cDNA (gifts from Prof. T. Yamamoto). The PCR was also carried out using the cDNA that had been extracted from the tissue sections after in situ PCR, and the resultant PCR products were electrophoresed through a 2% agarose gel along with those from the PC1 or PC2 cDNA.
Similarly, to identify 7B2 mRNA-positive cells in the pituitary gland, we performed in situ RT-PCR using degenerated oligonucleotides designed based on the specific region of rat 7B2 (Waldbieser et al. 1991) according to the method described above. The sequences of sense (primer 1) and antisense (primer 2) primers were as follows: 7B2 primer 1, 5′-CCTGCACATCAAGCCATGAA-3′; 7B2 primer 2, 5′-GAAAGGTTGTCCCGTACCTA-3′.
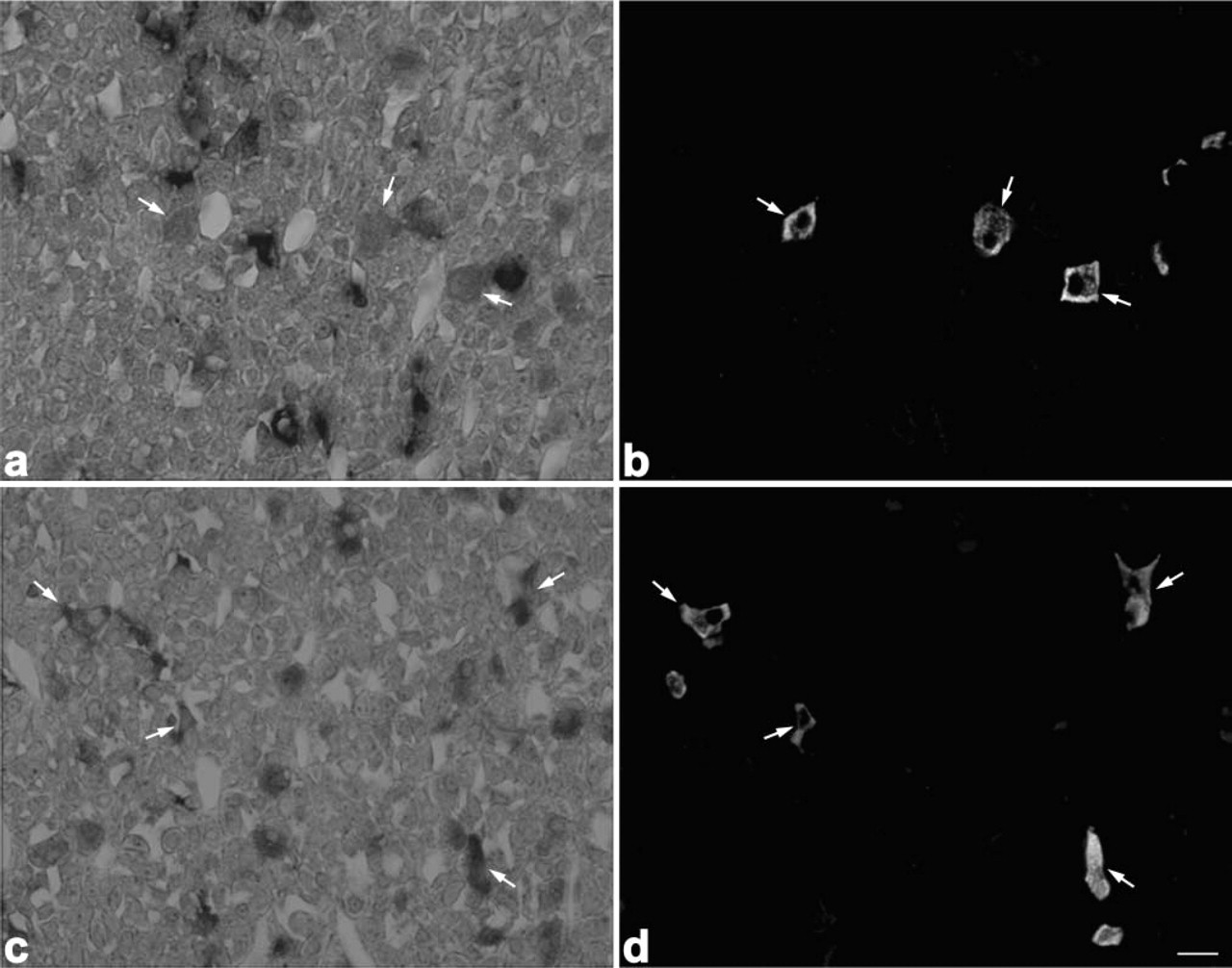
Light micrographs showing localization of PC1 and PC2 mRNAs in thyrotrophs of 8-week-old rats. PC1 mRNA (
Dual mRNA and Protein Staining
After the mRNA had been stained as described above, the sections were washed with 10 mM Tris-HCl containing 1 mM EDTA and fixed with 4% PFA in Milli-Q for 10 min. They were next twice washed with Milli-Q for 5 min each, three times with PBS for 5 min each, and immunolabeled by the immunofluoresence method. Before immunolabeling, for antigen retrieval, the sections were heated in a retrieval solution (10 mM EDTA in Milli-Q) at 120C for 3 min in an autoclave (Sanyo; Osaka, Japan). After the antigen retrieval, the sections were washed three times with PBS and blocked with 1% BSA-PBS for 1 hr. They were then incubated with guinea pig anti-amidated joining peptide (JP) (ST-3: 1:2000; Tanaka and Kurosumi 1992), rabbit anti-α-MSH (1:2000; Tanaka and Kurosumi 1986), rabbit anti-rat prolactin (PRL) (1:1000; supplied by Prof. K. Wakabayashi), goat anti-human thyroid-stimulating hormone (TSH)-β (1:2000; Biogenesis, New Fields, UK), guinea pig anti-human growth hormone (GH) (1:2000; prepared by our Department), or mouse monoclonal antibody against ovine luteinizing hormone (LH)-β (Uehara et al. 2001) for 21 hr, followed by FITC (fluorescein isothiocyanate)-labeled donkey anti-guinea pig IgG (1:400; Jackson Immunoresearch, West Grove, PA), Cy3 (indocarbocyanine)-labeled donkey anti-rabbit IgG (1:400; Jackson), or FITC-labeled donkey anti-mouse IgG (1:400; Jackson) for 2 hr. The sections were washed with PBS, mounted in PermaFluor (Immunon; Pittsburgh, PA), and observed with an Olympus BX-50 microscope equipped with a BX-fluorescence attachment (Olympus Optical; Tokyo, Japan).
The number of PC1 mRNA- or PC2 mRNA-positive corticotrophs, PC2 mRNA-positive α-MSH-producing cells, amidated JP-immunopositive cells, and α-MSH-immunopositive cells, regardless of whether the nucleus was visible or not, was counted: the ratio of PC1 mRNA- or PC2 mRNA-positive corticotrophs per amidated JP-producing cells, or PC2 mRNA-positive cells per α-MSH-producing cells was used as an index of each cell population in the mid-frontal plane of the gland. Results were from five sections obtained from each of four animals.
Light micrographs showing localization of PC1 and PC2 mRNAs in mammotrophs of 8-week-old rats. PC1 mRNA (
Immunofluorescence Labeling
Pituitary glands were fixed by immersion in Bouin–Hollande for 2 days. After dehydration and embedding in Paraplast, frontal sections (4 μm) were cut and mounted on gelatin-coated slides. After the deparaffinized sections had been rinsed in Milli-Q and PBS, they were immunostained using the double immunofluorescence method. Sections were incubated sequentially at room temperature with the following reagents: 5% normal goat serum (NGS) for 2 hr, the first antibody for 16 hr, and immunofluorescence-labeled secondary antibody for 2 hr, as described previously (Tanaka et al. 1997).
To estimate the ratio of corticotrophs bearing α-MSH or not, we first incubated sections with a mixture of rabbit anti-α-MSH (1:2000) and guinea pig anti-amidated JP (1:2000) serum, followed by incubation with a mixture of FITC-labeled donkey anti-rabbit IgG (1:400; Jackson) and Cy3-labeled donkey anti-guinea pig IgG (1:400; Jackson). After they had been rinsed three times with PBS and mounted in PermaFluor (Immunon), the specimens were examined under the fluorescence microscope.
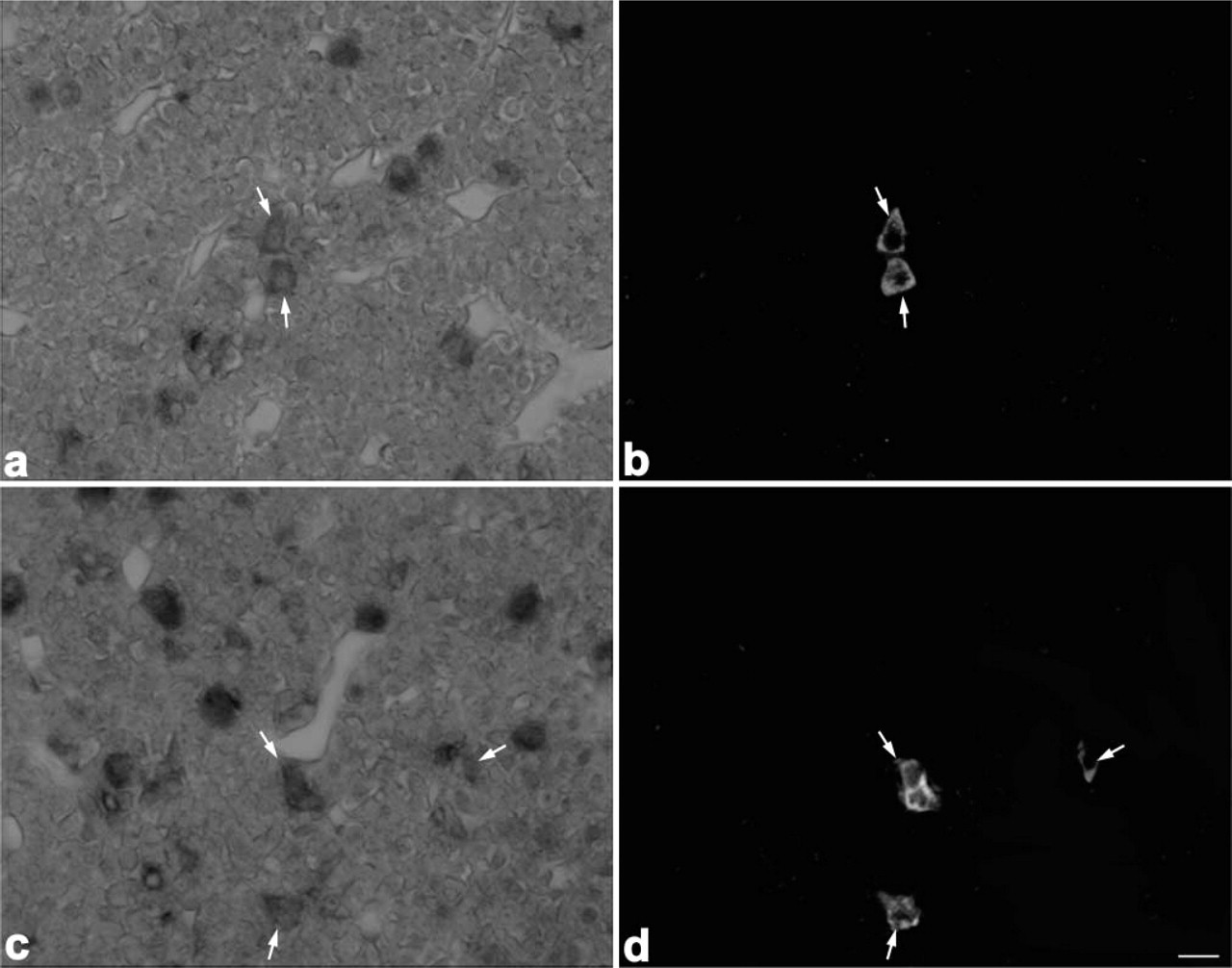
Light micrographs showing localization of PC1 and PC2 mRNAs in somatotrophs. PC1 mRNA (
The number of immunoreactive cells in each section was matched with each other, and the number of cells was counted. Data from five sections obtained from each of four animals were used for quantitative analysis.
As a control for specificity, diluted antisera were preabsorbed with their corresponding peptide immunogens at a final concentration of 1 μg/ml for 16 hr at 4C before immunolabeling.
Statistical Analysis
All data are presented as means ± SE. For statistical analysis, the Duncan multiple rage test was employed.
Results
Expression Sites of PC1 and PC2 mRNAs in Pituitary Gland
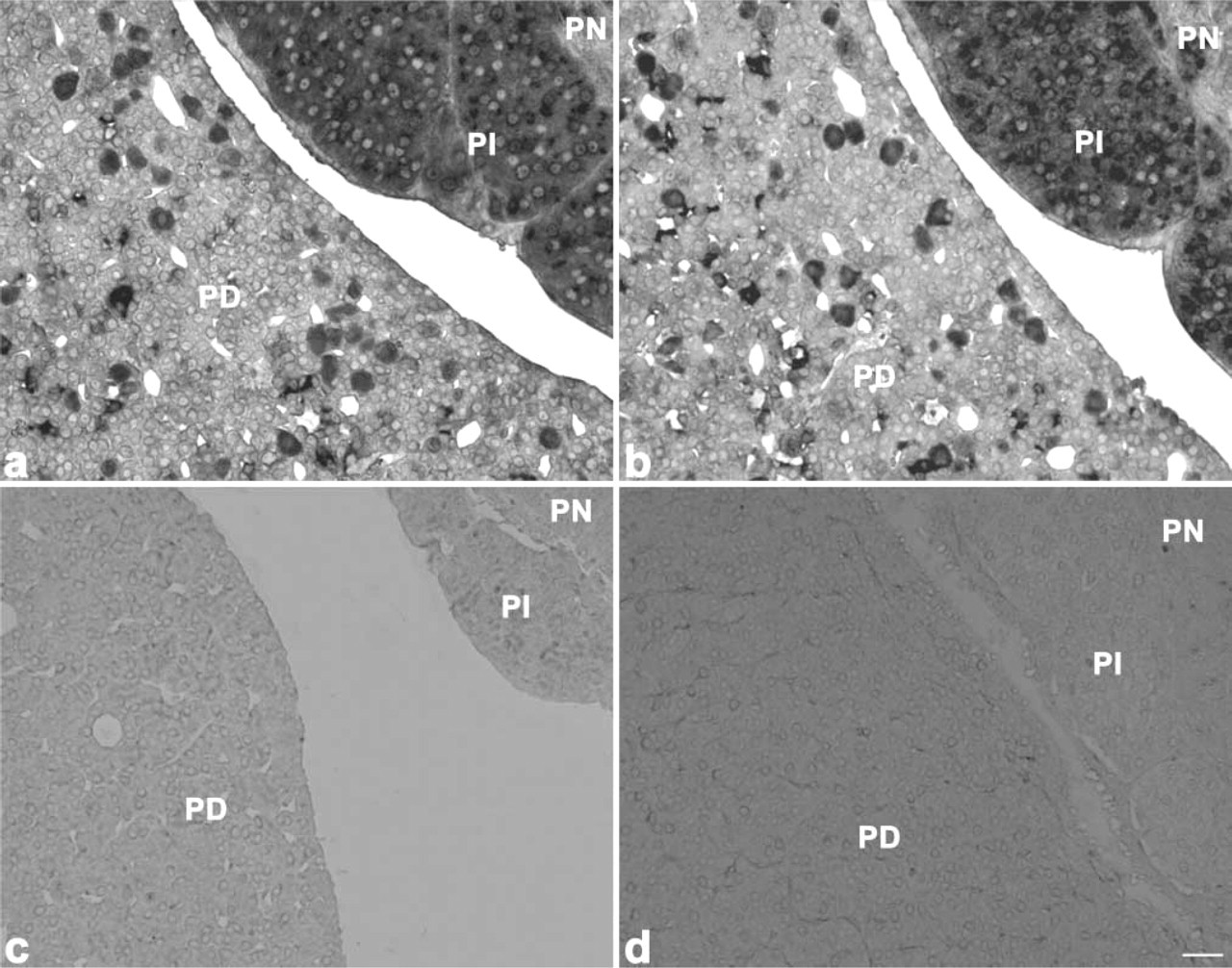
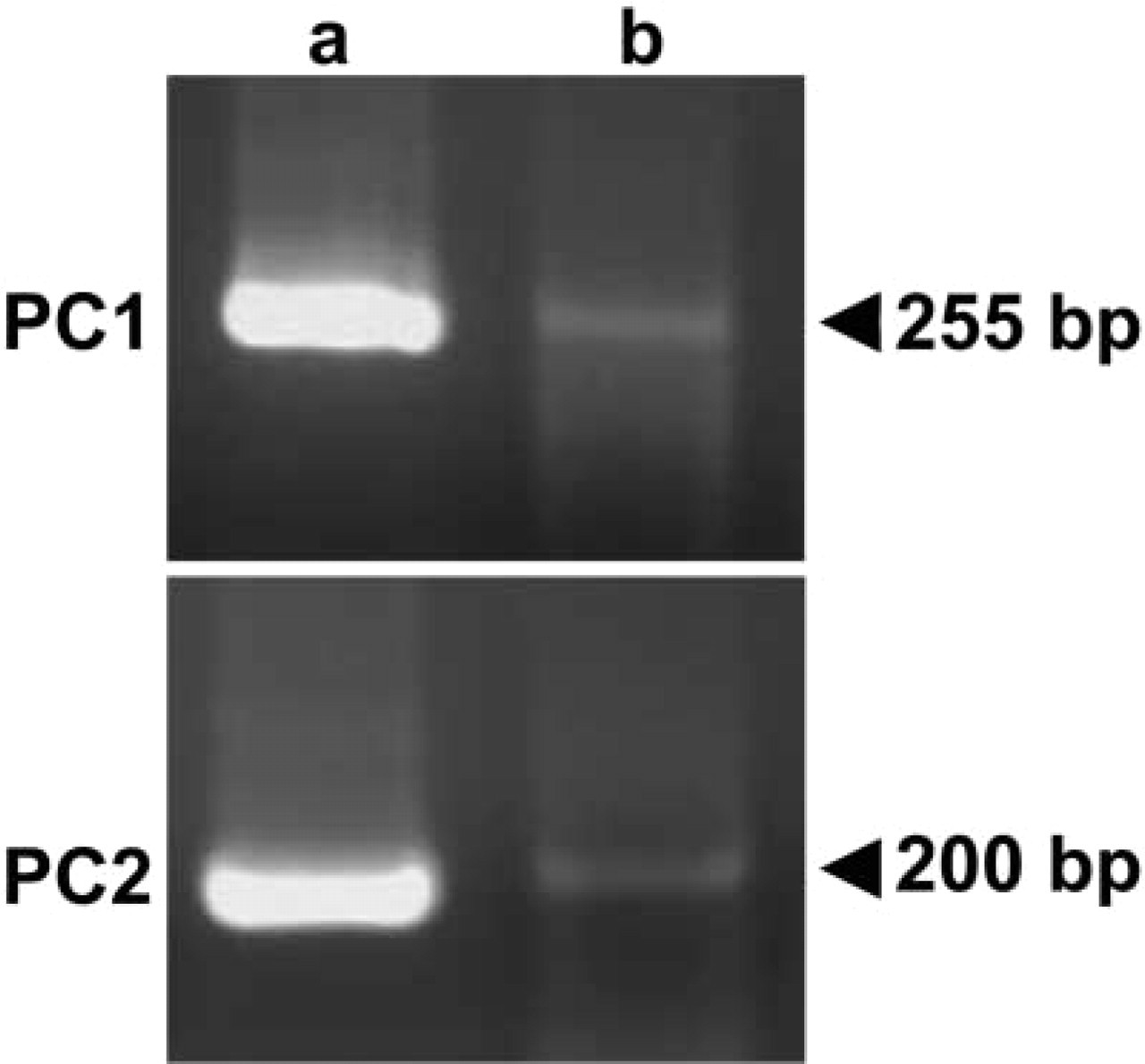
To identify PC1 mRNA- and/or PC2 mRNA-positive cells, we performed a double staining using in situ RT-PCR for PC1 and PC2 and immunofluoresence for pituitary hormones. Reactions indicating both PC1 mRNA and PC2 mRNA expression were observed in both the pars distalis and the pars intermediate (Figures 1a and 1b). However, no positive reaction was detected when the reverse transcriptase was omitted from the reaction mixture, thus indicating the validity of the in situ RT-PCR methodology used (Figures 1c and 1d). The authenticity of the in situ RT-PCR for PC1 or PC2 was confirmed by the second PCR, which revealed the expected bands with the same primer pairs, by using the harvested pituitary sections and plasmid DNA for rat PC1 and PC2 after in situ amplification (Figure 2).
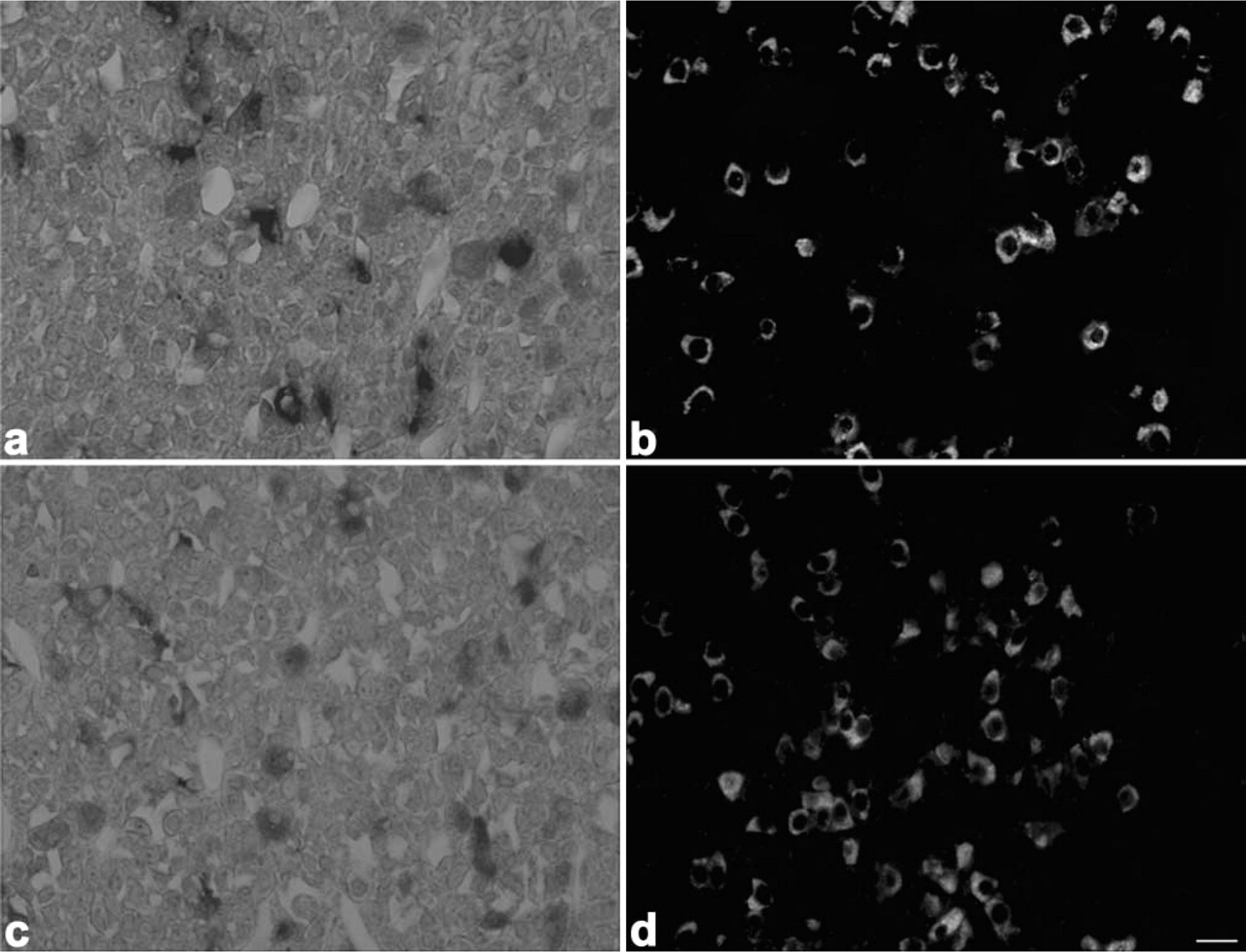
Light micrographs showing localization of 7B2 mRNA in corticotrophs, gonadotrophs, thyrotrophs, mammotrophs, and somatotrophs. 7B2 mRNA-positive cells (
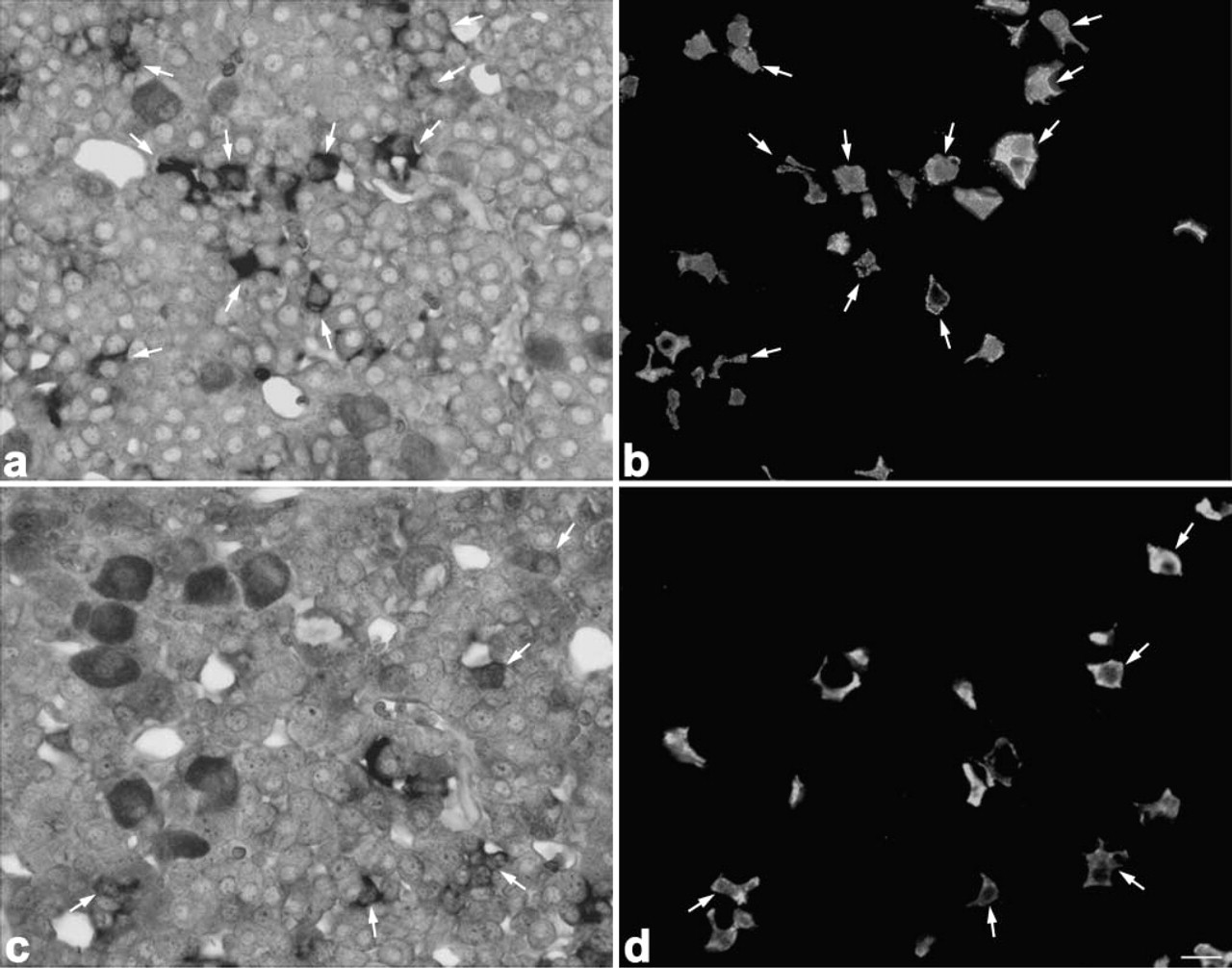
When sections from 8-week-old rats were immunostained with anti-amidated JP, anti-LHβ, anti-TSHβ, anti-PRL, and anti-GH after detection with the PC1 or PC2 probes, both PC1 and PC2 mRNAs were expressed in corticotrophs (Figure 3), gonadotrophs (Figure 4), thyrotrophs (Figure 5), and mammotrophs (Figure 6). However, only a small population of corticotrophs expressed both PC1 and PC2 mRNAs, with most of them expressing only PC1 mRNA (Figure 3). No PC1 and PC2 mRNAs were expressed in somatotrophs (Figure 7). 7B2 mRNA was observed in corticotrophs, gonadotrophs, thyrotrophs, and mammotrophs, but not in somatotrophs (Figure 8).
Changes in the Number of α-MSH-immunopositive Corticotrophs During Postnatal Life
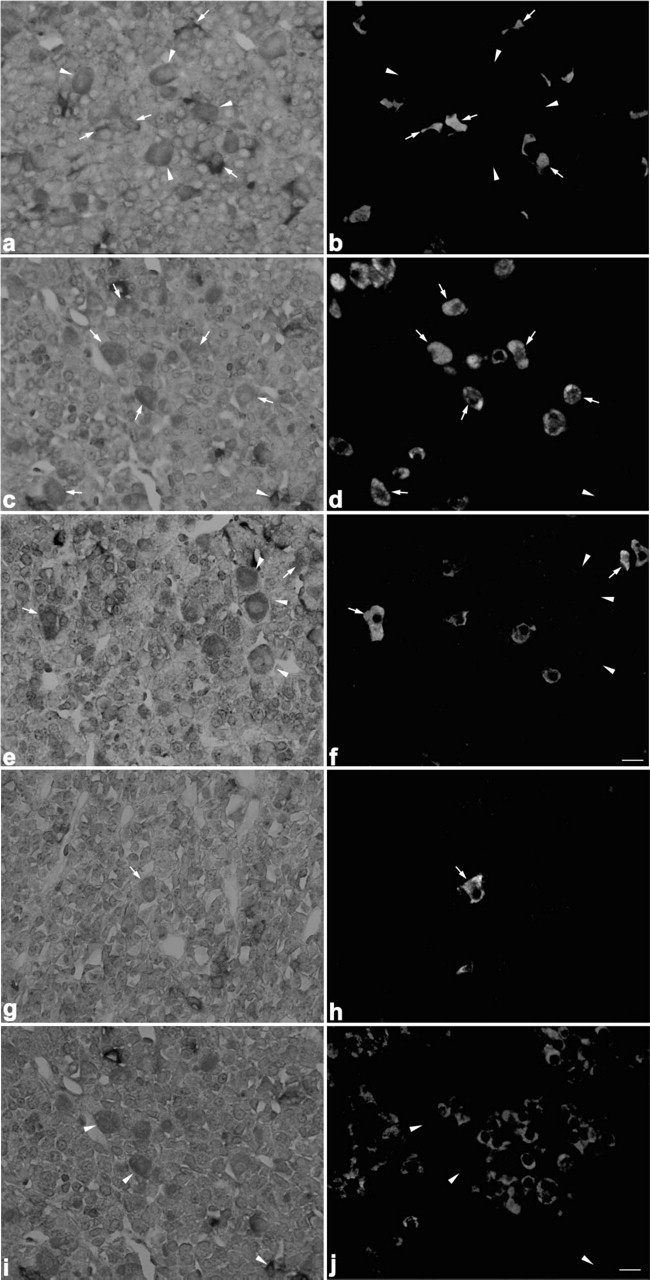
When pituitary sections of 8-week-old rats were stained for amidated JP and α-MSH by the double immunofluorescence technique, α-MSH-positive cells were hardly observed in the pars distalis. In pars distalis of the 0.5-day-old and 1-week-old rats, on the other hand, most of the corticotrophs, identified by reaction with anti-amidated JP, were also immunostained with anti-α-MSH (Figure 9). Then we quantified the ratio of the number of α-MSH-positive corticotrophs per total number of corticotrophs during postnatal life. As shown in Figure 10, most of the corticotrophs expressed α-MSH in 1-week-old rats, but the expression of α-MSH in the corticotrophs decreased gradually with advancing postnatal age, i.e., in the 3-week-old rats the α-MSH-positive corticotrophs decreased to a quarter of their earlier number and then were hardly seen in the 8-week-old animals. These data indicate that the pattern of POMC processing started to change to the adult pattern in the 3-week-old rats, similar to previous findings (Schafer et al. 1983; Sato and Mains 1985; Lugo et al. 1989).
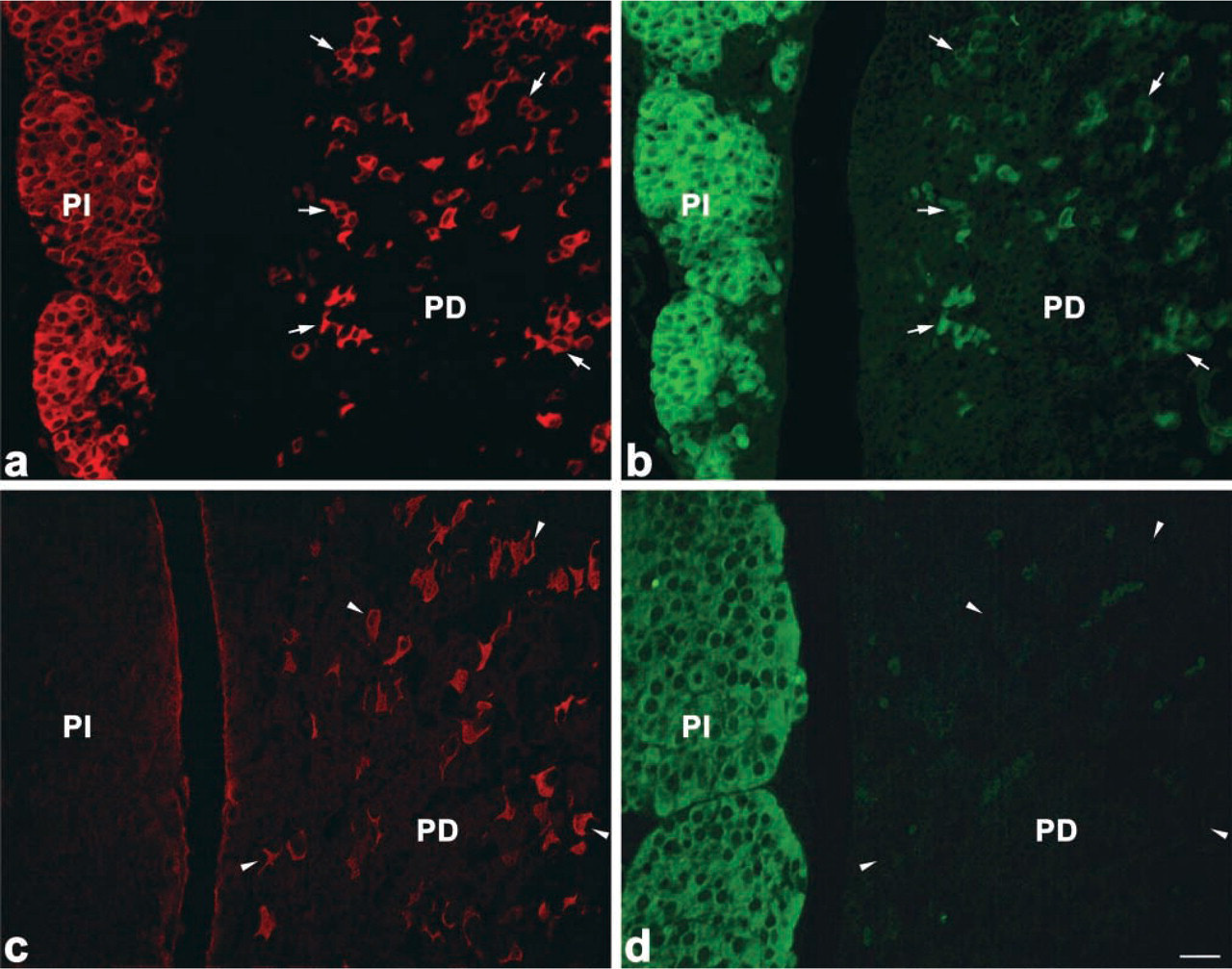
Immunofluorescence images showing differences in the number of α-MSH-immunopositive corticotrophs between the 1-week-old (
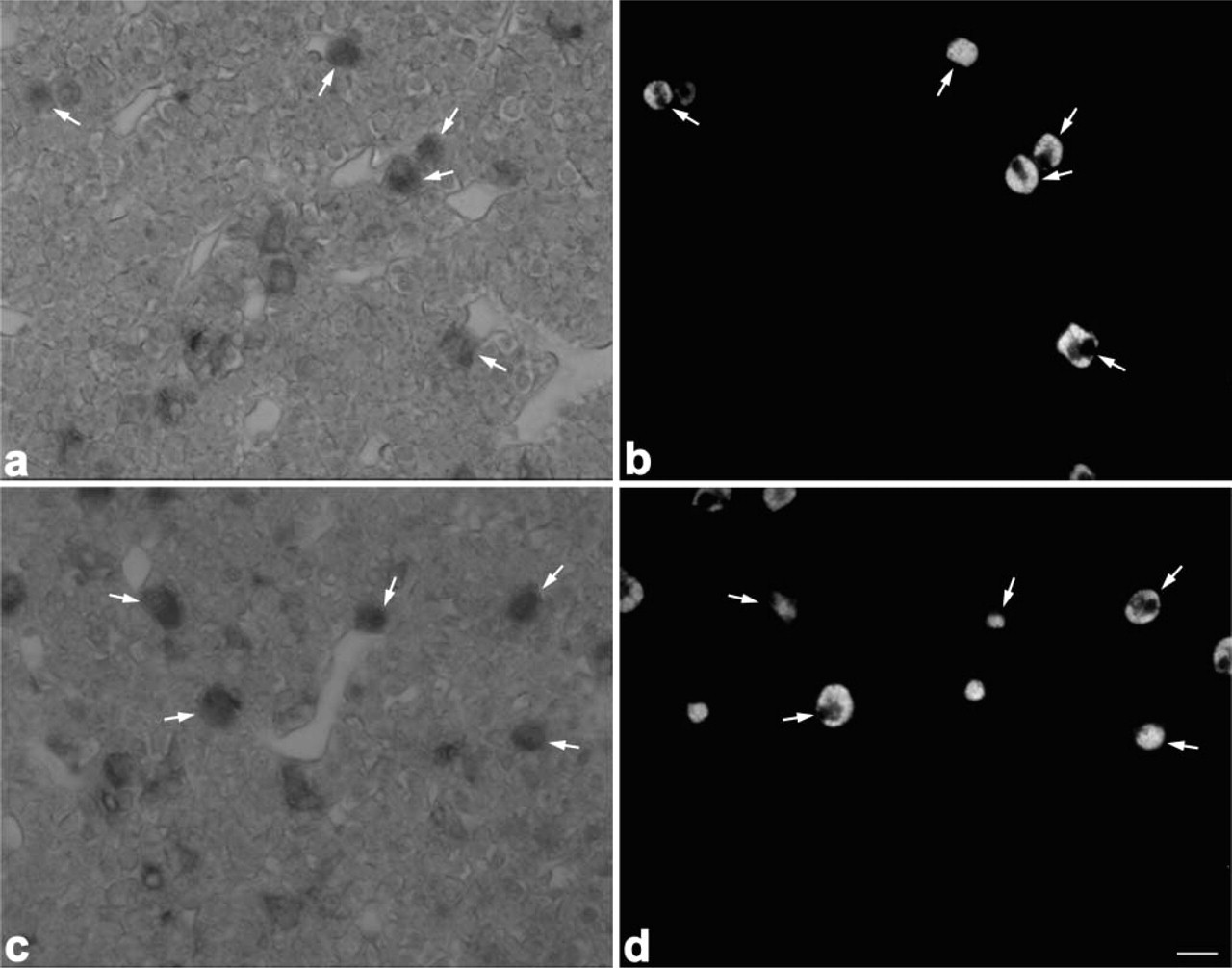
Expression Pattern of PC1 and PC2 mRNAs in Corticotrophs During Postnatal Life
The changes in the pattern of POMC processing are considered to depend on the relative expression levels of PC1 and PC2. Therefore, we compared the expression of PC1 and PC2 mRNAs in corticotrophs among 1-, 3-, and 8-week-old rats by using the dual staining techniques for amidated JP or α-MSH proteins and either PC1 or PC2 mRNA (Figure 11). In the corticotrophs of 1-week-old rats, PC1 mRNA- and PC2 mRNA-positive cells were seen: PC1 mRNA-positive cells represented 60.7 ± 1.2% of the total corticotrophs and PC2 mRNA-positive ones 89.2 ± 1.5%. In the corticotrophs of 3-week-old and 8-week-old rats, the percentages for PC1 mRNA-positive cells were 65.8 ± 6.5% and 62.7 ± 9.9%, and those for PC2 mRNA-positive cells were 39.2% and 27.9% (Figures 12a and 12b). Thus, the number of PC2 mRNA-positive cells significantly decreased in 3-week-old rats compared with that for the 1-week-old rats (p <0.001). When we also quantified the number of PC2 mRNA-positive cells among α-MSH-immunopositive corticotrophs, a graded decrease was observed in the ratio of PC2 mRNA-positive cells per α-MSH-positive corticotrophs: 93.0 ± 1.5% in 1-week-old, 32.5 ± 3.1% in 3-week-old, and 11.9 ± 1.7% in 8-week-old rats (Figure 12c).
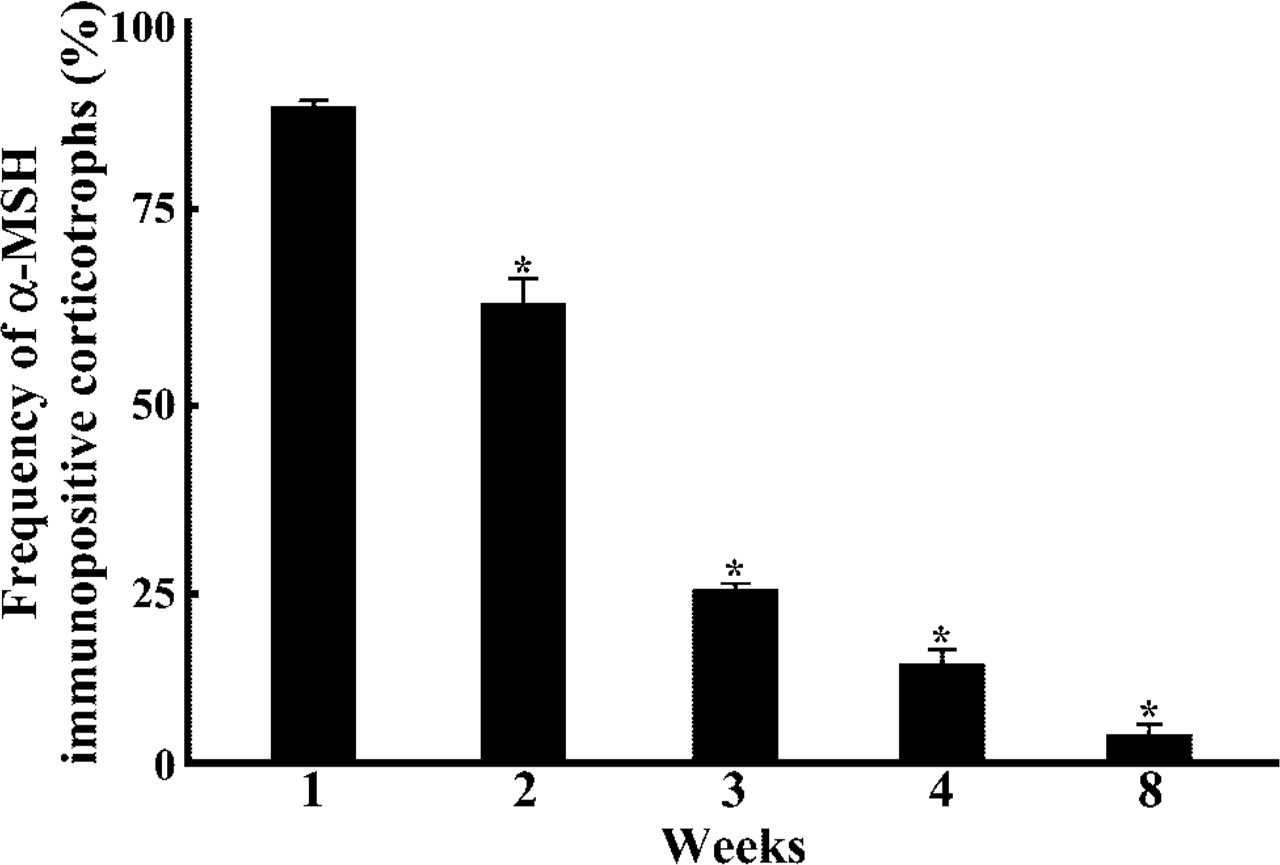
Frequency of α-MSH-immunopositive corticotrophs during postnatal development. ∗p <0.001, 1-week-old rat vs 2-week-old rat, 2-week-old rat vs 3-week-old rat, 3-week-old rat vs 4-week-old rat, 4-week-old rat vs 8-week-old rat.
Discussion
The present study, using in situ RT-PCR, demonstrated that PC1 and PC2 mRNAs were expressed in several cell types of the pars distalis of the rat pituitary. Previously, we examined the localization of PC1 and PC2 proteins in the rat pituitary gland by using anti-PC1 (ST-28) and anti-PC2 (ST-29) against synthetic peptides covering amino acids 442–459 and 613–629 of the respective mouse proteins (Tanaka et al. 1996), the sequences of which are identical to those of the rat (Hakes et al. 1991). However, the PC1- and/ or PC2-immunopositive reaction was observed only in gonadotrophs in the pars distalis and in melanotrophs in the pars intermedia (Uehara et al. 2001). Then we tried to detect them by using the conventional ISH technique but we failed to obtain a sufficient intensity of reaction. Finally, we decided to use an in situ RT-PCR method with higher sensitivity. As a result, we were able to obtain a clear reaction in several cell types of the pars distalis of the rat. The ISH using staining by chromogens also provides higher resolution for recognizing positive cells compared with ISH using radioisotopes. Thus, we demonstrated that this in situ RT-PCR method was useful for detecting low amounts of signal in histological sections. The results were also confirmed by solution-phase RT-PCR on the pituitary sections and the plasmid DNA for rat PC1 and PC2 by using the same primer pairs.
In the present study we showed that PC1 mRNA-positive cells corresponded to corticotrophs in the pars distalis. This finding implies that proteolytic cleavage of POMC by PC1 would produce ACTH (1–39) in this lobe. On the other hand, both PC1 and PC2 mRNAs were expressed in the pars intermedia. Therefore, ACTH (1–39) liberated from POMC would be further cleaved into α-MSH and CLIP in this part of the pituitary. This finding is consistent with previous data obtained from rats (Day et al. 1992). The present study also indicates that PC1 and PC2 mRNAs of the 8-week-old rats were expressed in gonadotrophs, thyrotrophs, and mammotrophs but that corticotrophs expressed mainly PC1 mRNA. It is conceivable that proprotein processing of the hormones does not take place in these cell types. However, it is possible that other proteins, such as granin family proteins, with proteolytic cleavage sites are contained in the secretory granules and that their proteins are cleaved by PC1 and/or PC2 (Arden et al. 1994; Dittie and Tooze 1995; Hoflehner et al. 1995; Eskeland et al. 1996; Laslop et al. 1998). Indeed, we earlier showed that both PC1 and PC2 proteins were expressed in rat pituitary gonadotrophs, suggesting that these convertases may be involved in the processing of secretogranin II and chromogranin A (Uehara et al. 2001). The expression of PC1 and PC2 mRNAs in thyrotrophs and mammotrophs may be involved in the processing of granins because of the existence of chromograin B, secretogranin II, and secretogranin III in these cells (Sakai et al. 2003). Furthermore, the present study detected no expression of PC1 and PC2 mRNAs in somatotrophs. This may be attributed to the fact that somatotrophs do not contain these granins (Sakai et al. 2003).
In the present study we found that 7B2 mRNA-positive cells are expressed in several kinds of trophs bearing PC2 mRNA, suggesting that the expression of PC2 correlates positively with that of 7B2. Because the 7B2 protein is considered to participate in the activation of PC2 (Martens et al. 1994; Mbikay et al. 2001), this system may also function in the processing of pro-proteins in other neuroendocrine cells in addition to POMC processing in corticotrophs.
Interestingly, the present study revealed that both PC1 and PC2 mRNAs were expressed in most corticotrophs of the neonatal rats and that the ratio of PC2 mRNA-positive corticotrophs decreased gradually as the postnatal period proceeded from week 1 to week 8. Similar results were reported for the mouse by Marcinkiewicz et al. (1993), who used the double immunostaining method with rabbit anti-PC1 or PC2 antisera and rabbit anti-α-MSH serum to reveal co-localization of them. In the present study, we showed that the number of PC1 mRNA-positive corticotrophs in the 8-week-old rats was lower than that of the PC2-mRNA-positive ones. We are clearly unable to give the reason for this difference, but it might be attributable to a more rapid turnover of PC1 mRNA than of PC2 mRNA. Furthermore, we could observe more numerous α-MSH-immunopositive cells by treatment for antigen retrieval in adult rats compared with the normal immunolabeling in them (cf. Figures 9 and 11). In our previous immunoelectron microscopic study, we demonstrated that many corticotrophs contained a small number of secretory granules immunolabeled for α-MSH, but these cells are not detected by conventional light microscopic immunohistochemistry (Tanaka and Kurosumi 1986). Consequently, these α-MSH-immunopositive corticotrophs may have become detectable after the antigen retrieval.
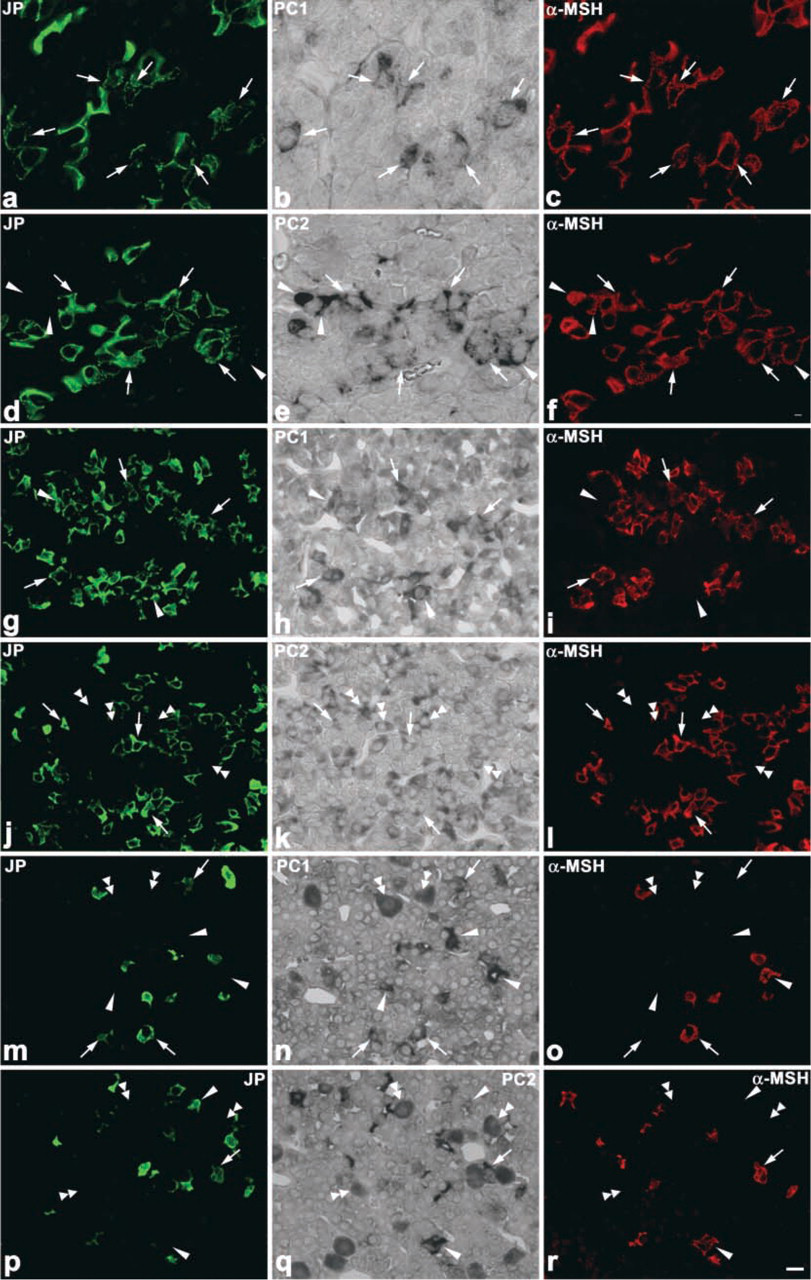
Expression levels of PC1 and PC2 mRNAs in 1-week-old (
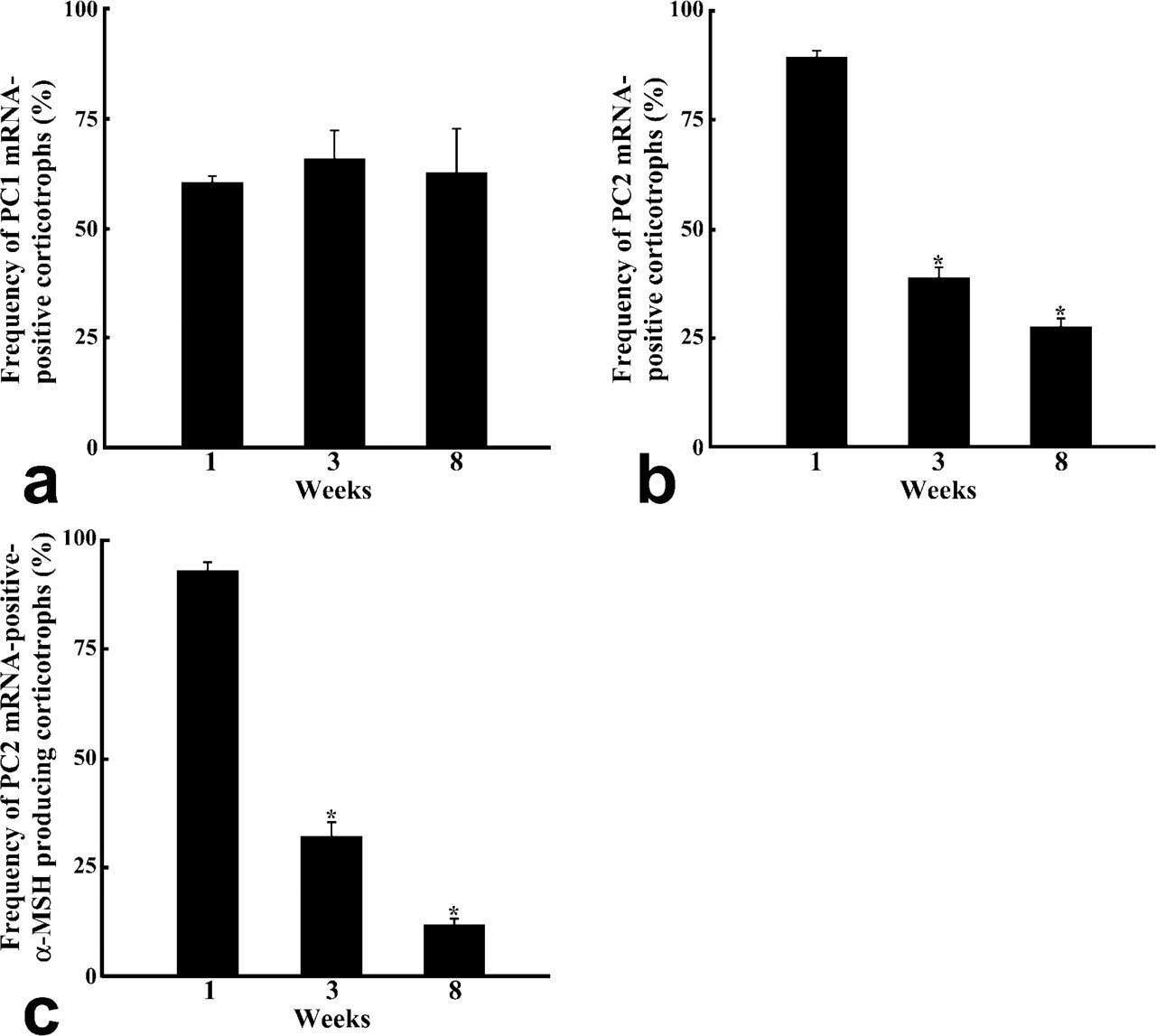
Expression changes in the frequency ratio of PC1 mRNA-expressing (
The first 2 weeks of the postnatal period are known to be a stress-nonresponsive period, and a lack of corticosterone secretion is observed then in response to various stressors (Schapiro 1962; Schoenfeld et al. 1980). Proteolytic processing of ACTH to α-MSH may facilitate the maintenance of a lower level of endogenous corticosterone. An earlier in vitro experiment showed that corticosterone suppressed the cleavage of ACTH to α-MSH in animals older than postnatal 3 weeks, but not in animals during this stress-nonresponsive period (Noel and Mains 1991). In the near future it will be necessary to clarify the molecular mechanism causing the difference in the corticosterone responsiveness between these periods. The cleavage of ACTH into α-MSH may be a security measure to cease the secretion of a large amount of corticosterone. Therefore, the lower levels of corticosterone during the postnatal period may also afford a favorable environment to allow T-cells to escape from apoptosis induced in the thymus by corticosterone (Henning 1978; Compton et al. 1987; Inomata and Nakamura 1989).
α-MSH is also considered to have stimulatory effects on intrauterine growth (Swaab et al. 1976) and growth-stimulating effects on the adrenal zona glomerulosa (Robba et al. 1986). Similarly, in Ambystoma, corticotrophs were shown to produce α-MSH during the larval period (Dores et al., 1989, 1990, 1993). Recently, we found that PC2 mRNA was expressed in the corticotrophs of bullfrog tadpoles, but not in those of the adults (Yaoi et al. 2003). The production of α-MSH during the neonatal period or larval period is considered to be a general phenomenon. Taken together, the present data demonstrated that the α-MSH production varied directly in accordance with the expression of PC2.
Footnotes
Acknowledgements
Supported in part by a grant-in-aid for science research from the Ministry of Education, Culture, Sports, Science, and Technology of Japan to ST.
We thank Prof T. Yamamoto and Dr R. Fukabori (Division of Behavioral Physiology, Department of Behavioral Sciences, Graduate School of Human Sciences, Osaka University) for supplying the plasmid DNA for rat PC1 and PC2, and Prof K. Wakabayashi (Institute for Molecular and Cellular Regulation, Gunma University) for supplying rabbit anti-PRL.