
Abstract
The prohormone convertases PC1 and PC2 are subtilisin-related endopepti-dases that process prohormone and neuropeptide precursors. Using different ultrastructural immunocytochemical approaches, we have investigated their intracellular distribution in a neuroendocrine cell type that has not been examined thus far, the rat anterior pituitary lactotrope. These cells secrete mainly prolactin and also express the neuroendocrine-specific protein secretogranin II, which is considered a peptide precursor. Our study provides evidence for the expression of PC1 and PC2 in rat lactotropes and provides new information on their subcellular localization. Apart from their presence in the secretory granules, PC1 and PC2 displayed different major localization along the secretory pathway. PC1 immunoreactivity was concentrated in the Golgi apparatus, whereas PC2 immunoreactivity was prominent in the rough endoplasmic reticulum (RER). These observations provide morphological support for previous biochemical analysis of proPC1 and proPC2 post-translational processing, which has demonstrated that PC1 exits very rapidly from the RER, whereas PC2 is retained much longer in this compartment.
Keywords
T
The cloning of these enzymes has provided specific tools that allow the study of their expression in neuroendocrine tissues. In situ hybridization experiments have shown that PC1 is the major PC expressed in the anterior pituitary and that PC2 is also present in substantial amounts in this tissue (Day et al. 1992). The analysis of the anterior pituitary cell types expressing these enzymes was, however, restricted to the corticotropes because they are the only pituitary cells that express a major precursor that is endoproteolytically processed into active peptides, i.e., pro-opiomelanocortin (POMC) (Day et al. 1992). Immunocytochemistry was not performed to identify other pituitary cells that express PC1 and PC2. This questioned the pattern of expression of the PCs in this heterogeneous tissue as well as the substrate of these enzymes. The other anterior pituitary cell types, i.e., lactotropes, somatotropes, thyrotropes, and gonadotropes, are not specialized in the secretion of a proteolytic maturation product. However, all of them express proteins of the granin family, which contains three major members: chromogranin A (CgA) and B (CgB) and secretogranin II (SgII) (reviewed in Huttner et al. 1991a,b; Winkler and Fisher–Colbrie 1992; Fisher–Colbrie et al. 1995). The exact function of these proteins is not definitely established, but they are considered as peptide precursors (reviewed in Natori and Huttner 1994). CgA is the precursor of several peptides whose activities have been well documented (reviewed in Iacangelo and Eiden 1995), and proteolytic fragments of CgB and SgII have also been purified from tissue extracts (reviewed in Winkler and Fisher-Colbrie 1992; Fisher-Colbrie et al. 1995). More recently, a direct precursor-product relationship between SgII and several derived peptides has been demonstrated by pulse-chase experiments (Dittie and Tooze 1995; Muller and Tougard 1995), and the activity of secretoneurin, a peptide derived from SgII, has been proposed (Kirchmair et al. 1993; Saria et al. 1993). We have already reported the expression of SgII in prolactin (PRL) cells (Tougard et al. 1989; Ozawa et al. 1994) and demonstrated its processing in the TGN and secretory granules (Muller and Tougard 1995; Muller et al. 1997). Because PC1 and PC2 are the best candidates for SgII processing, we have studied the expression of PC1 and PC2 in PRL cells by electron microscopic immunocytochemistry. Our results provide the first evidence for the expression of both enzymes in this cell type. Our ultrastructural study also shows that PC1 and PC2 display different subcellular distributions along the secretory pathway that correlate with the steps of their posttranslational processing.
Materials and Methods
Antibodies
We used rabbit polyclonal antibodies directed against the carboxy terminal segment of mPC1 (AbC-mPC1; segment 629–726) (Seidah et al. 1991) and the C-terminal segment of mPC2 (AbC-mPC2; segment 529–637) (Seidah et al. 1990). The specificity of these antibodies was previously well documented by immunohistochemical (Marcinkiewicz et al. 1993) and immunoprecipitation techniques (Benjannet et al. 1993). The specificity of rabbit antiserum raised against purified rat PRL RP-3 (a gift from NIDDK) has been described in detail elsewhere (Duhau et al. 1991).
Pre-embedding Immunoperoxidase Staining
This labeling was performed directly on rat anterior pituitary cells in primary culture as previously described (Tougard et al. 1980; Tougard and Picart 1986). Briefly, cells were obtained by enzymatic dispersion of male Wistar rat (weight 200 g) anterior pituitaries, as described elsewhere (Tixier-Vidal et al. 1973). For experiments, 4 × 105 cells were plated per 35-mm tissue culture dish in 2 ml of Ham's F 12 medium supplemented with 10% horse serum, 2.5% fetal calf serum, and antibiotics. Cells were grown for 7 days at 37C in 5% CO2. They were fixed in periodate-lysine-paraformaldehyde (MacLean and Nakane 1974) for 2 hr at room temperature (RT), washed, preincubated in 50 mM NH4Cl, and permeabilized with 0.005% saponin. After incubation with specific rabbit antibodies, cells were incubated with goat Fab anti-rabbit IgG conjugated with peroxidase (Biosys; Compiègne, France). After washing, cells were postfixed in 1% glutaraldehyde and the peroxidase activity was visualized using 3,3′-diaminobenzidine tetrahydrochloride. Cells were postfixed in 1% osmium tetroxide, dehydrated, and embedded in situ in Epon according to Brinkley et al. (1967). Selected areas of immunoreactive cells were sectioned as previously described (Picart and Tixier-Vidal 1974). Ultrathin sections were examined at the electron microscope without further staining.
Postembedding Immunogold Staining
Small pieces of male Wistar rat anterior pituitaries were fixed with a mixture of 4% paraformaldehyde-0.05% glutaraldehyde in 0.1M cacodylate buffer (pH 7.4) for 2 hr at 4C. After washing, the tissues were treated with 2% uranyl acetate in 50 mM maleate buffer for 2 hr at 4C, partially dehydrated, and embedded in LR White according to Ozawa et al. (1994).
Ultrathin sections were immunostained as previously reported (Ozawa et al. 1994). Briefly, they were incubated with the first specific antibodies for 2 hr and stained with 10-nm protein A-gold particles (Biocell; Cardiff, UK) for 1.5 hr. After rinsing, the sections were air-dried, stained for 15 min with osmium tetroxide vapors, and for 15 sec with 2% uranyl acetate in 50% alcohol. The grids were finally rinsed in alcohol, dried, and examined under the electron microscope. Control sections were obtained by omitting incubation with the specific antibodies or by replacing it with normal rabbit serum.
Double immunostaining
Double Immunogold Staining
The double staining was performed on the same face of the grid. After the first complete immunostaining (i.e., incubation with the first specific antibodies and then with 10-nm protein A-gold particles), the grids were incubated for 1 hr with unconjugated protein A (0.1 mg/ml in TBS) (Sigma; St Louis, MO), as previously described (Ozawa et al. 1994). After rinsing, they were incubated with the second specific antibodies and then with 5-nm protein A-gold particles. After washes, they were finally stained and dried as described above for single immunola-beling. Control sections of the double immunostaining were performed by omitting incubation with the second specific antibody or by replacing it with normal rabbit serum.
Double Pre- and Postembedding Immunostaining
After the pre-embedding immunoperoxidase staining of PC1 or PC2 in rat anterior pituitary cells in primary culture (as described above), ultrathin sections of immunoreactive cells were then immunostained using the postembedding immunogold approach, with antiserum against rat PRL and then with 10-nm protein A-gold particles.
Western Blotting
Rat anterior pituitaries were homogenized at 4C in a lysis buffer (0.01 M PBS, pH 7.4, containing 0.5% sodium deoxy-cholate and 0.5% Nonidet P40) in the presence of 200 μM PMSF, 1 μM leupeptin, and 1 μM pepstatin. GH3B6 rat tumor PRL cells, cultured for 6 days, were scraped on ice in the same buffer. Lysates were then centrifuged for 10 min at 15,000 × g at 4C and the supernatants were further solubilized with SDS-PAGE sample buffer containing 50 mM DTT. After separation by SDS-PAGE, proteins were electrotransferred to nitrocellulose sheets (Schleicher & Schuell; Keene, NH). After incubation with the specific antibodies, immunoreactive proteins were revealed with alkaline phosphatase-conjugated sheep anti-rabbit IgG antibodies (Promega; Madison, WI).
Results
Using both immunoperoxidase and immunogold labeling. PC1 and PC2 were detected in most of the glandular cells of the rat anterior pituitary in situ and in primary culture. The immunoreactive sites corresponded to the intracellular compartments involved in the secretory pathway. Differences in the intensity of the immunolabeling and in the respective subcellular distribution of the two PCs were conspicuous, even at low magnification, using the pre-embedding approach and immunoperoxidase staining. With this approach and whatever the cell type, the intensity of PC1 labeling was always higher than that of PC2 (compare Figures 1 and 2). PC1-specific immunoperoxidase staining concerned mainly the Golgi stacks, which were conspicuously labeled in all glandular cells. The rough endoplasmic reticulum (RER) cisternae were very slightly labeled (Figure 1). On the contrary, PC2 was detected only in the RER cisternae (Figure 2). The Golgi saccules were not immunostained. With this approach, as has been previously described for secretory products (Tougard and Picart 1986), the secretory granules were only weakly labeled. To investigate the presence of the PCs within these organelles, we performed an immunogold labeling using a postembedding approach. Some gold particles specific for PC1 (Figure 3a) or for PC2 (Figure 3b) were observed within the matrix of several secretory granules in all glandular cells (Figure 3).
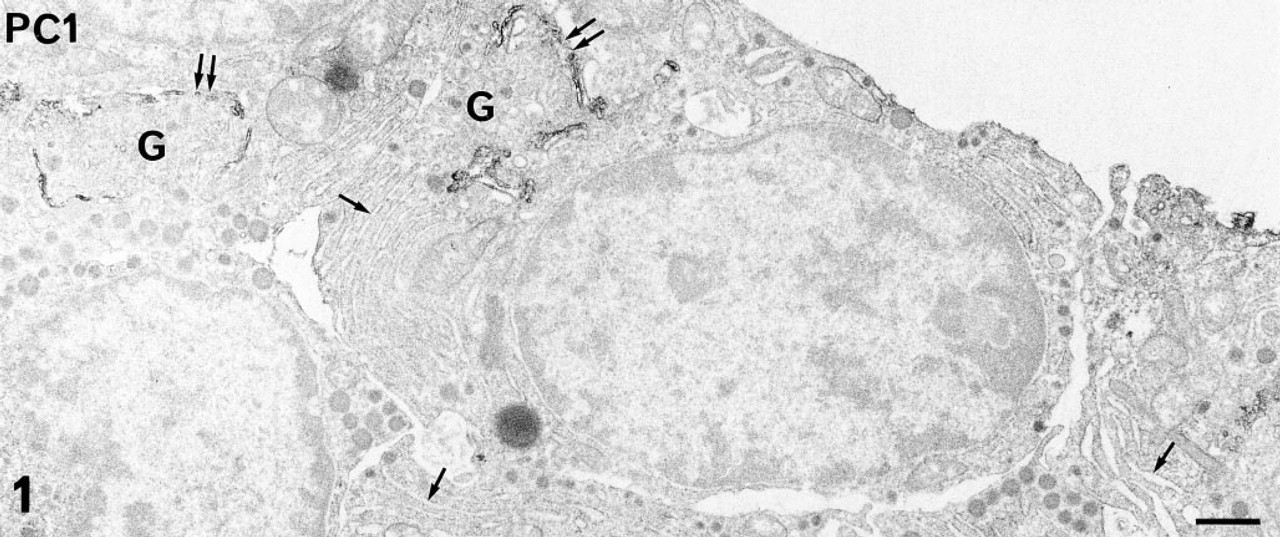
PC1 immunolabeling in anterior pituitary cells in primary culture revealed using the pre-embedding immunoperoxidase staining. At this low magnification, PC1 is detected mainly in the Golgi (G) stacks (double arrows) in all glandular cells. The RER cisternae (arrows) are very slightly stained. Bar = 1 μm.
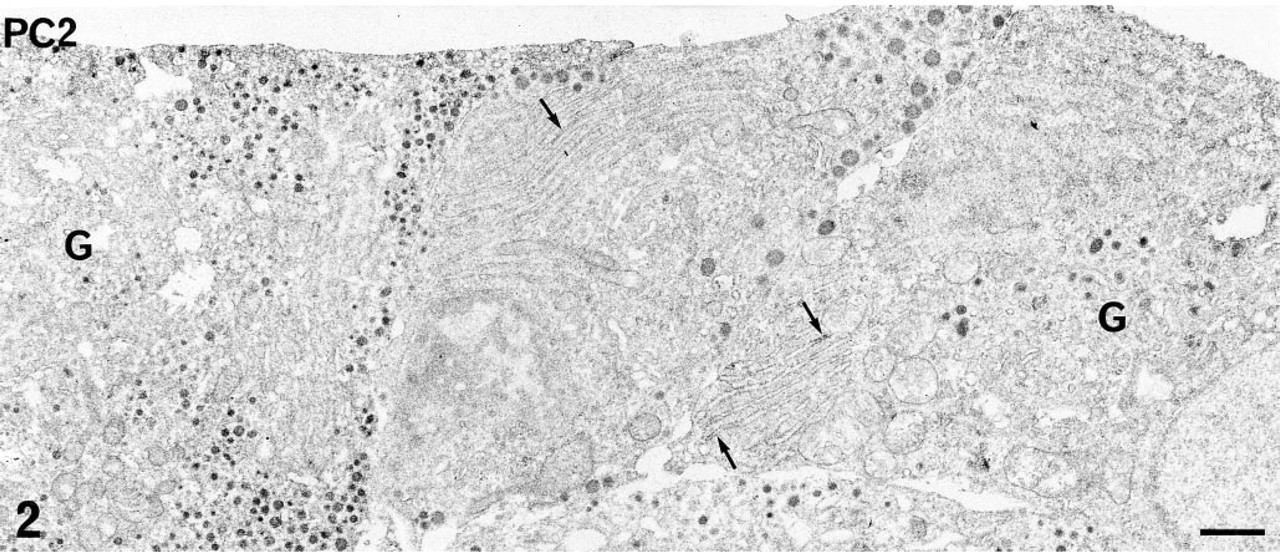
PC2 immunolabeling in anterior pituitary cells in primary culture revealed using the pre-embedding immunoperoxidase staining. At this low magnification, PC2 is detected associated with the membrane of several RER cisternae (arrows) in some glandular cells. The smooth saccules in the Golgi zone (G) are not immunostained. Bar = 1 μm.
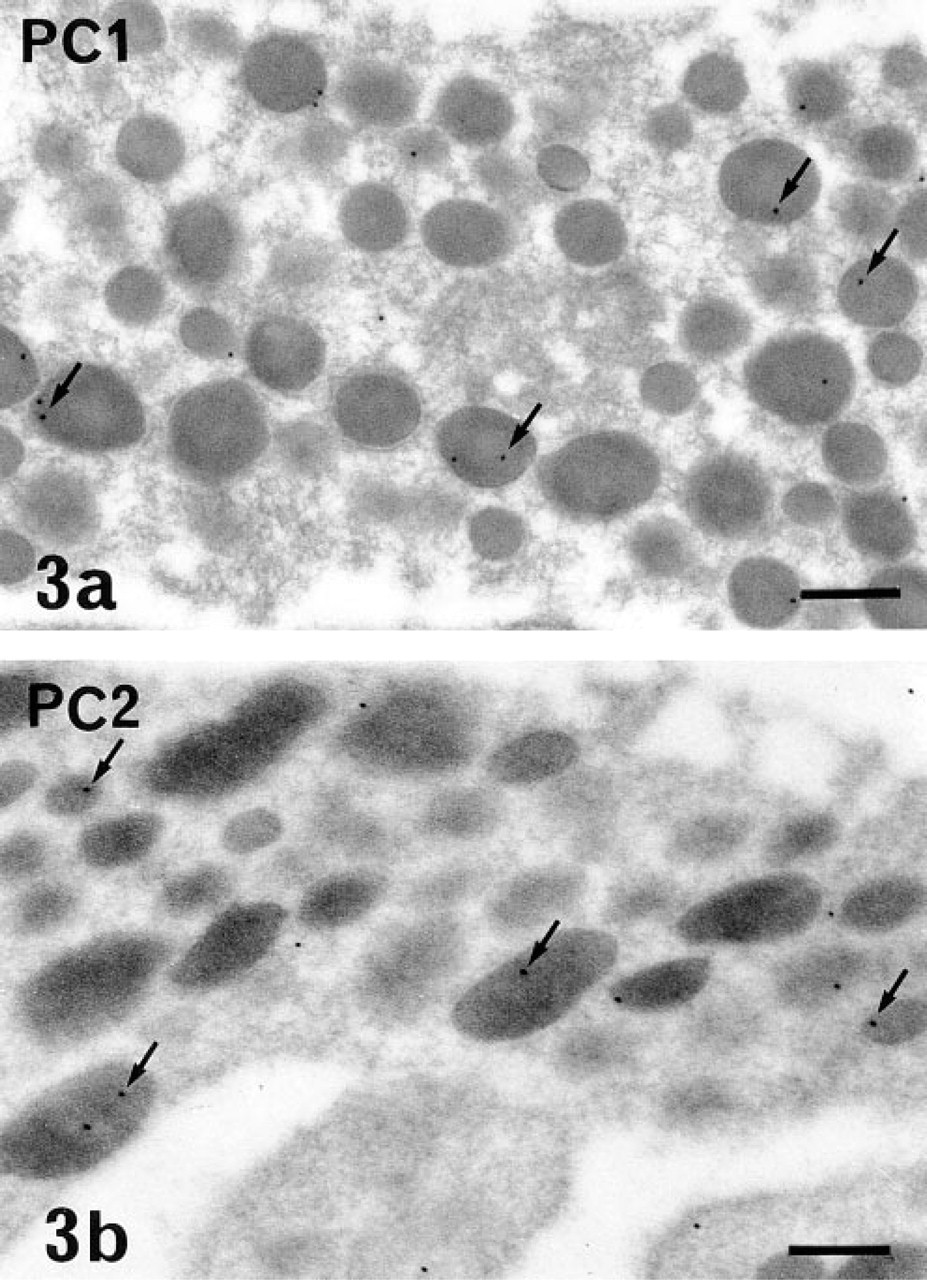
PC1 (
Because the present study was focused on the immunolocalization of the PCs in PRL cells, we performed double immunostaining of either PC1 or PC2 and of PRL to specifically identify the PRL cells. These double immunostainings confirmed that PRL cells expressed both PC1 and PC2. Using a combination of pre- and postembedding approaches and immunoperoxidase to localize the PCs and immunogold to identify PRL cells, intense and preferential distribution of PC1 within the Golgi saccules (Figure 4) and preferential localization of PC2 on the membrane of the flattened RER cisternae (Figure 5), characteristic of PRL cells, were confirmed in PRL cells. The specific hormonal product, visualized with immunogold, was detected on the matrix of immature and mature secretory granules (Figures 4 and 5). The double immunogold staining with anti-PC1 or anti-PC2 and anti-PRL also confirmed that the two PCs are present in at least some PRL secretory granules (Figure 6). Because the gold particles corresponding to immunoreactive PC1 or PC2 were less numerous than those corresponding to immunoreactive PRL, a portion of the PRL secretory granules appeared devoid of detectable PCs.
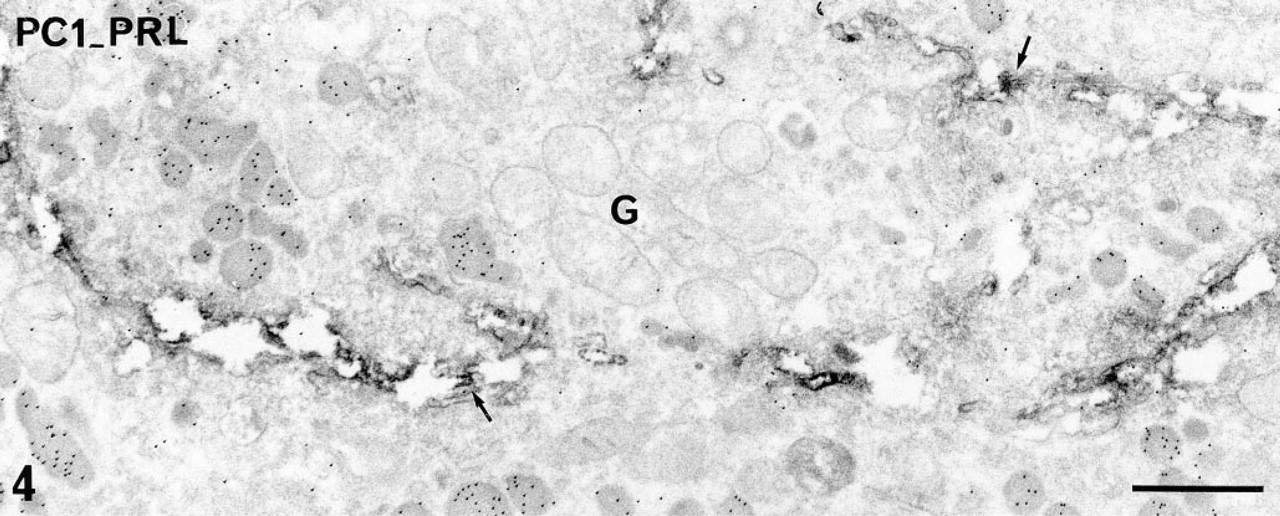
Double pre- and postembedding immunolabeling of PC1 (pre-embedding immunoperoxidase) and PRL (postembedding immunogold) in a PRL cell in primary culture. Immunoreactive PC1 is detected in all the Golgi saccules (arrows). Ten-nm gold particles revealing immunoreactive PRL are observed on the matrix of immature secretory granules in the Golgi zone (G) and on the matrix of mature secretory granules. Bar = 1 μm.
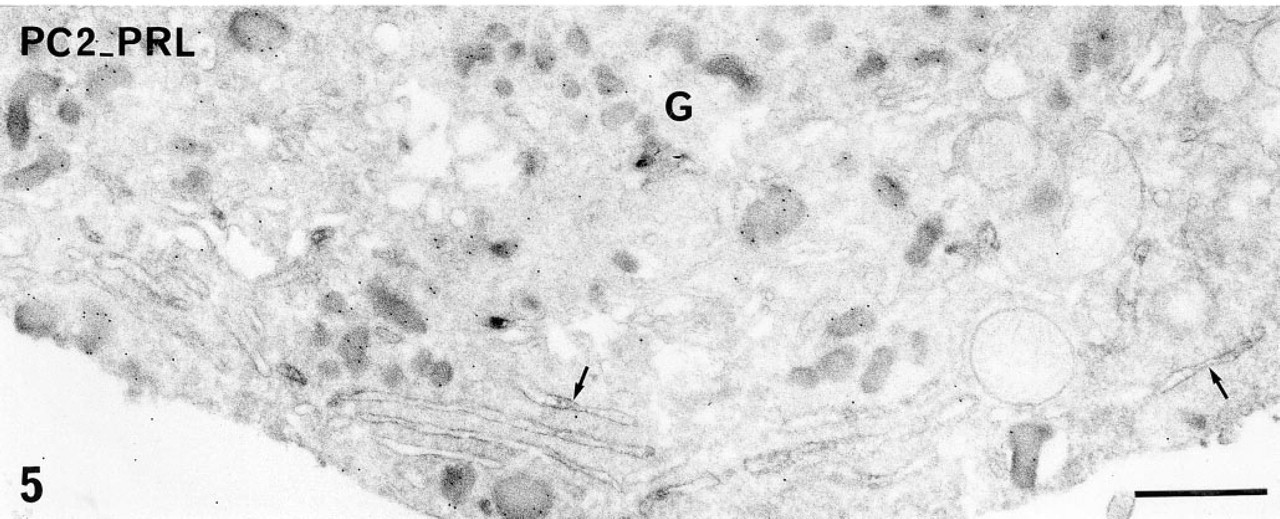
Double pre- and postembedding immunolabeling of PC2 (pre-embedding immunoperoxidase) and PRL (postembedding immunogold) in a PRL cell in primary culture. Immunoreactive PC2 is detected on the membrane of the flattened RER cisternae (arrows). The smooth saccules of the Golgi zone (G) are devoid of peroxidase reaction product. Ten-nm gold particles revealing immunoreactive PRL are observed on the matrix of immature and mature secretory granules. Bar = 1 μm.
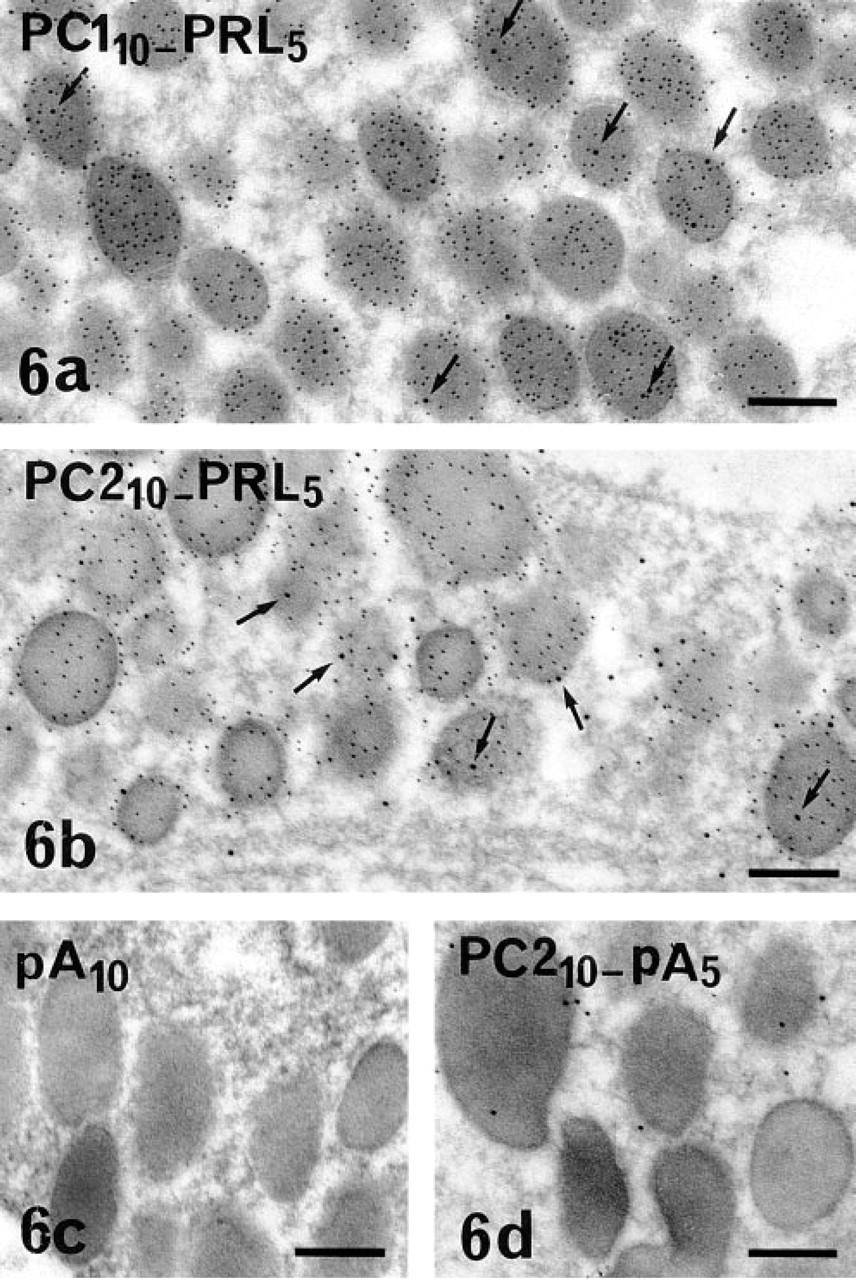
Double immunogold labeling of PC1 or PC2 (10-nm gold) and PRL (5-nm gold) in PRL cells in anterior pituitary. A few 10-nm gold particles revealing (
The presence of PC1 and PC2, in the rat anterior pituitary and in the GH3B6 rat PRL cell line was confirmed by the immunoblotting analysis. The antibodies specific for the C-terminal segment of either PC1 or PC2 recognized two large bands in both cell extracts, as already described in the literature. The anti-PC1 antibodies detected a 91-kD band that corresponds to the precursor form and an 84-kD band that corresponds to the mature enzyme (Figure 7). The anti-PC2 antibodies also detected two bands of 75 kD and 65 kD that correspond to the precursor form and to the mature enzyme, respectively (Figure 7). However, the intensity of the immunolabeled bands varied in the two cell extracts. Whereas the dominant form of PC2 was the mature enzyme in the anterior pituitary extracts, pro-PC2 and mature PC2 were as intensely labeled in the GH3B6 cell extracts. This difference was not observed for PC1. These discrepancies could be related to the difference in post-translational processing of the two enzymes (see Discussion) and to the difference in the storage compartment of our two cell models. Most of the glandular cells of the anterior pituitary contained many large secretory granules, whereas GH3B6 cells were characterized by very few small secretory granules.
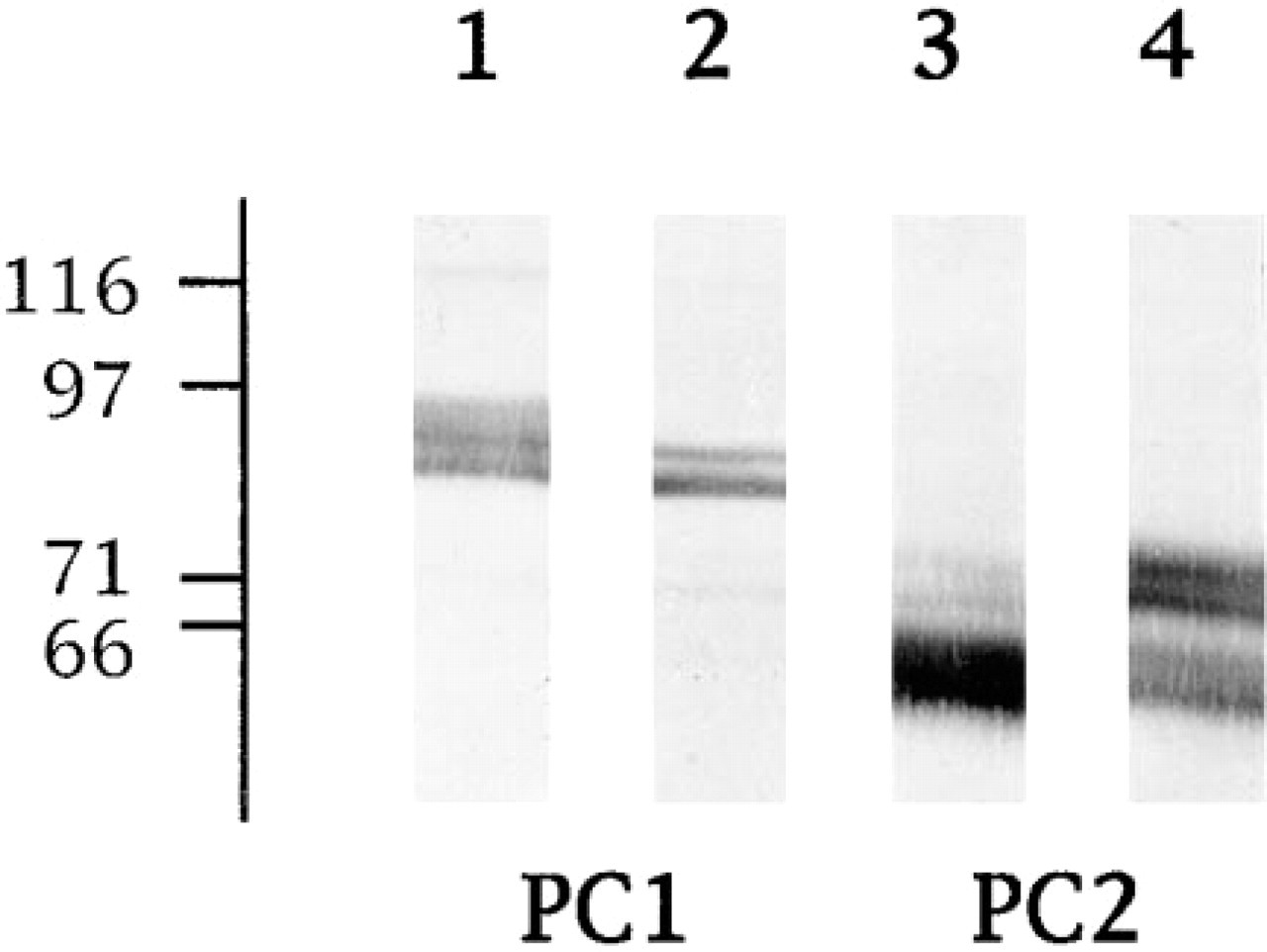
Immunoblotting of PC1 and PC2 in protein extracts of rat anterior pituitaries (Lanes 1 and 3) and GH3B6 cells (Lanes 2 and 4). The proteins (25 μg per lane) were separated by SDS-PAGE on 7.5% gels in the presence of 50 mM DTT, electrotransferred, and immunostained using either anti-PC1 (Lanes 1 and 2) or anti-PC2 (Lanes 3 and 4) antibodies. The position of the molecular weight standards is indicated at left (kD).
Discussion
In the present work, we provide immunocytochemical evidence for the expression of PC1 and PC2 in rat lactotropes.
Previous works have demonstrated the cell-specific distribution of PC1 and PC2 in several neuroendocrine and endocrine cells. These studies were related to the expression of these enzymes in pancreatic β-cells (Smeekens et al. 1992; Malide et al. 1995) and in pituitary cells that express POMC (Seidah et al. 1990, 1991; Day et al. 1992), i.e., in melanotropes of the intermediate lobe and in corticotropes of the anterior lobe. The only data reported thus far for other cell types of the anterior pituitary suggested the expression of PC1 in gonadotropes (Marcinkiewicz et al. 1993). This was, however, not restricted to this cell type, and our study indeed identifies, for the first time, the lacto-tropes as one of the cell types of the anterior pituitary that express both PC1 and PC2. The presence of these two PCs in PRL cells was confirmed by their immunochemical detection in GH3B6 cells, a homogeneous model of rat PRL cells.
The present work also provides original information about the subcellular distribution of these pro-teolytic enzymes. Both are present in the secretory granules of these cells, but they display other different major localization sites along the secretory pathway. PC1 is mainly observed in the Golgi apparatus and PC2 in the RER. As has been previously pointed out for the immunolabeling of PRL (Tougard et al. 1980), the two immunocytochemical procedures performed in this study provide complementary information. The pre-embedding approach, using a moderate permeabilization step and immunoperoxidase, is the best approach to detect secretory products within the RER and the Golgi stacks, whereas the postembedding approach, using immunogold, permits the best detection within the matrix of secretory granules.
Several groups had already shown the presence of PC2 (Guest et al. 1992; Smeekens et al. 1992; Malide et al. 1995) and PC1 (Malide et al. 1995) in the secretory granules of β-cells. The most recent of these studies has co-localized PC1 and PC2 in the same secretory granules (Malide et al. 1995). The concentration of PC1 immunoreactivity in the Golgi apparatus that we observed in the present work is in agreement with the distribution of PC1 in AtT-20 cells already described (Hornby et al. 1993). PC2 immunoreactivity was more prominent in the RER. These observations are consistent with biochemical data concerning proPC1 and proPC2 post-translational processing. PC1 exits from the RER after cleavage of its propeptide within 10 min after synthesis (Benjannet et al. 1993; Zhou and Lindberg 1993; Lindberg 1994; Zhou and Mains 1994). PC2 processing is much slower. The cleavage of the propeptide occurs hours after synthesis, and analysis of PC2 glycosylation as well as transport blockade experiments have demonstrated that proPC2 is retained in the RER (Guest et al. 1992; Benjannet et al. 1993; Shen et al. 1993; Zhou and Mains 1994). ProPC2's unique post-translational processing is related to its interaction with the neuroendocrine protein 7B2 (Braks and Martens 1994; Benjannet et al. 1995; Zhu and Lindberg 1995). Whereas PC2 is not further processed after the cleavage of the propeptide, PC1 is truncated at the C-terminus (Vindrola and Lindberg 1992) in the TGN or in the secretory granules (Lindberg 1994; Zhou and Mains 1994). We cannot visualize this truncated form because the antibodies used in our study are specific for the C-terminal segment of PC1. All these data are in agreement with the differential distribution of PC1 and PC2 in the RER and the Golgi stacks that we observed using the pre-embedding immunoperoxidase staining.
The presence of both PCs in PRL cells raises the question of the precursors that are proteolytically processed within these cells. The major secretory product of lactotropes is PRL, which is released into the medium without further major proteolytic processing other than the removal of the signal peptide in the RER. Some minor proteolytic products have been identified but do not correspond to potential cleavage sites by a PC (reviewed in Sinha 1995). Other secretory proteins expressed in PRL cells may be the substrates proteolytically processed by PC1 and PC2. We have already demonstrated the presence of Sg II and Cg B in PRL cells (Tougard et al. 1989; Ozawa et al. 1994) as well as the proteolytic processing of Sg II in the anterior pituitary cells in primary culture and in GH3B6 cells and stimulation of release of SgII-derived peptides by thyroliberin (TRH) (Muller and Tougard 1995; Muller at al. 1997). This result had suggested that the lactotropes and possibly the thyrotropes could process Sg II. Moreover, stable overexpression of PC2 in PC12 had suggested the role of this convertase in the processing of Sg II (Dittie and Tooze 1995), whereas infection of GH4C1 with PC1 and PC2 suggested the involvement of PC1 in the cleavage of Sg II (Hoflehner et al. 1995). Our study provides evidence for the presence of both enzymes within PRL secretory granules and hence cannot distinguish which PC is involved in the processing of SgII in these cells. PC1 and PC2 could also be involved in the proteolytic processing of neuropeptide precursors, such as pro-TRH, that have been found in the rat anterior pituitary (cf. rev. Tougard and Tixier–Vidal 1994) and which can be cleaved by these PCs (Friedman et al. 1995; Pu et al. 1996).
In conclusion, the observations reported here demonstrate the expression of PC1 and PC2 in lactotropes of the rat anterior pituitary and strengthen the role of these two prohormone convertases in the proteolytic maturation of precursors of secretory proteins within this cell type.
Footnotes
Acknowledgements
Supported by grants from INSERM (U 36).
We gratefully acknowledge Dr Iris Lindberg (New Orleans) for helpful discussions. We also thank Eric Etienne for the photographic work. We acknowledge the National Institute of Diabetes and Digestive, and Kidney Diseases (NIDDK) for the gift of reference rat PRL (rat PRL-RP3).