
Abstract
The enzyme 20α-hydroxysteroid dehydrogenase (20α-HSD) catalyzes the conversion of progesterone into its inactive form, 20α-hydroxyprogesterone. To gain information about the exact sites of 20α-HSD mRNA expression, we performed in situ hybridization using a 35S-labeled cRNA probe in tissues of adult mice of both sexes. 20α-HSD mRNA was expressed in both male and female gonads. In the ovary, high expression was found in luteal cells of corpora lutea, while much lower expression could be detected in granulosa cells of growing follicles. In the testis, a specific hybridization signal was detected only in Leydig cells. In the female reproductive tract, 20α-HSD mRNA was found in the epithelial cells of the uterine cervix. In the adrenal cortex, only the zona reticularis exhibited specific radiolabeling, the expression being very high in the female and very low in the male. In the skin, specific labeling was restricted to sebaceous glands, the hybridization signal being much higher in the female than in the male. In the liver, 20α-HSD mRNA was found in hepatocytes, with a higher degree of expression in the female. In the kidney, specific labeling was observed in the epithelial cells of distal convoluted tubules, the signal being also much more striking in the female than in the male. In non-reproductive tissues, it clearly appears that the expression of 20α-HSD mRNA is higher in the female than in the male, suggesting that 20α-HSD may play an important role in reducing the intracellular concentration of progesterone originating from the circulation at a much higher level in the female.
Keywords
T
Thus far, the precise localization of 20α-HSD at the cellular level has not been reported. We therefore used in situ hybridization (ISH) to localize the 20α-HSD mRNA in several tissues in adult mice of both sexes to gain information about the exact sites of 20α-HSD expression and to determine whether there is a gender-related influence on the expression of the enzyme.
Materials and Methods
Animals
Four adult male (26–30 g) and female (24–27 g) C57BL6 mice were housed under constant temperature (21 ± 1C) and light (lights on from 0600 to 2000 hr) regimen. The animals received Purina Chow (Ralston-Purina; St Louis, MO) and tapwater ad libitum. The experiment was conducted in an animal facility approved by the Canadian Council on Animal Care (CCAC) and by the Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC). The study was performed in accordance with the CCAC Guide for Care and Use of Experimental Animals. The animals were all perfused between 0900 and 1000 hr for histological procedures as described below. The females were on diestrous day 1.
Histological Procedures
All the animals were perfused transcardially with 50 ml 4% (w/v) paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). The different tissues, i.e., liver, kidney, jejunum, pancreas, skin, adrenal, pituitary, testis, prostate, ovary, uterus, vagina, and mammary gland were excised and postfixed in the same fixative for 24 hr at 4C. The tissues were placed in 15% sucrose in 0.1 M phosphate buffer before being quickly frozen in isopentane chilled in liquid nitrogen.
In Situ Hybridization
Frozen sections (10 μm thick) were serially cut at −20C and mounted on gelatin- and poly-
After hybridization, coverslips were removed and slides were rinsed in 2 × SSC at RT for 30 min. Sections were digested by RNase A (20 μg/ml in 2 × SSC) at 37C for 30 min, rinsed in decreasing concentrations of SSC (2 × SSC and 1 × SSC) for 30 min at RT, washed in 0.5 × SSC for 30 min at 37C, followed by 90 min at RT in 0.5 × SSC, at 60C in 0.1 × SSC, and finally for 30 min at RT in 0.1 × SSC.
The sections were then dehydrated and exposed to
Kodak Biomax MR films for 3–8 days before being coated with liquid photographic emulsion (Kodak-NTB2; diluted 1:1 with water). Slides were exposed for 1–45 days, developed in Dektol developer (Kodak; Rochester, NY) for 2 min, and fixed in rapid fixer (Kodak) for 4 min. Thereafter, tissues were rinsed in running water for 30 min, counterstained with hematoxylin, and rapidly dehydrated through graded concentrations of ethanol, cleared in toluene, and cover-slipped with Permount (Fisher Scientific; Montreal, PQ, Canada).
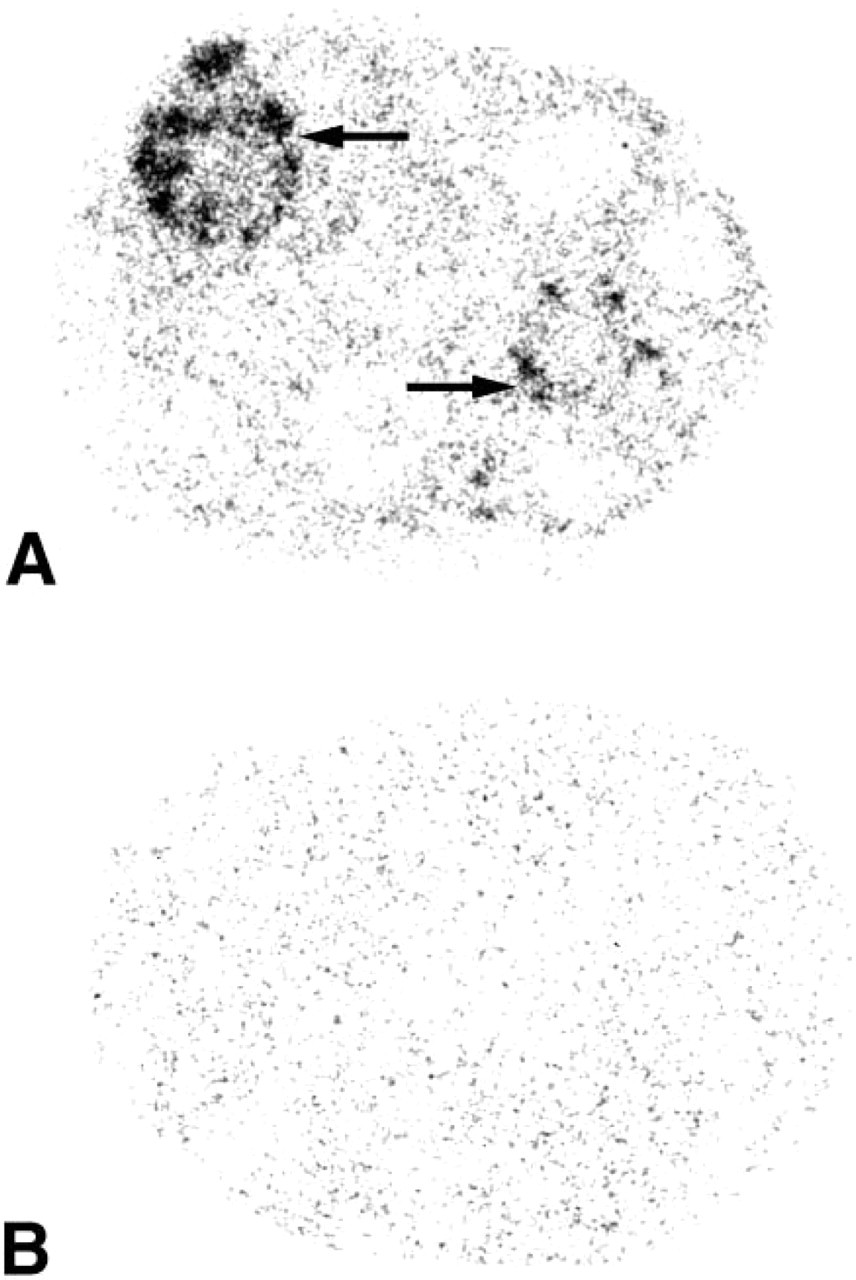
X-ray film autoradiographs illustrating the localization of 20α-HSD mRNA in the ovary. (
Results
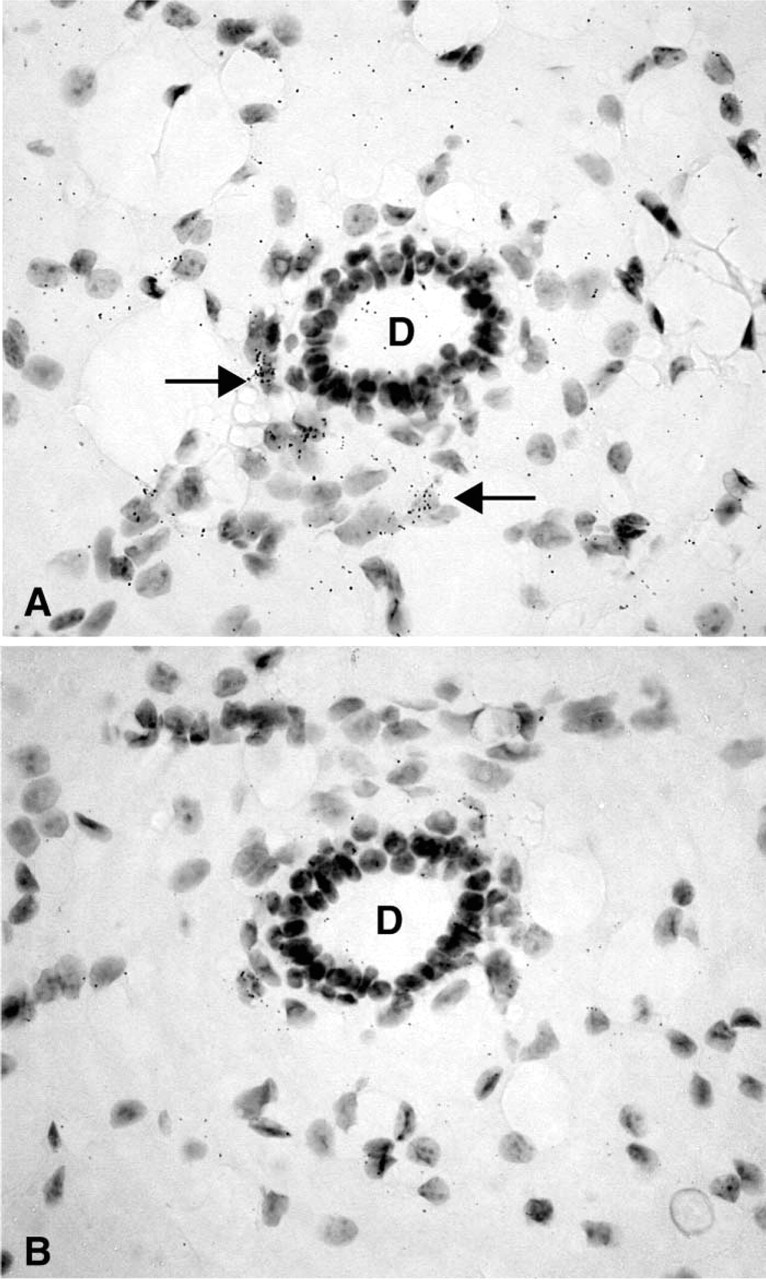
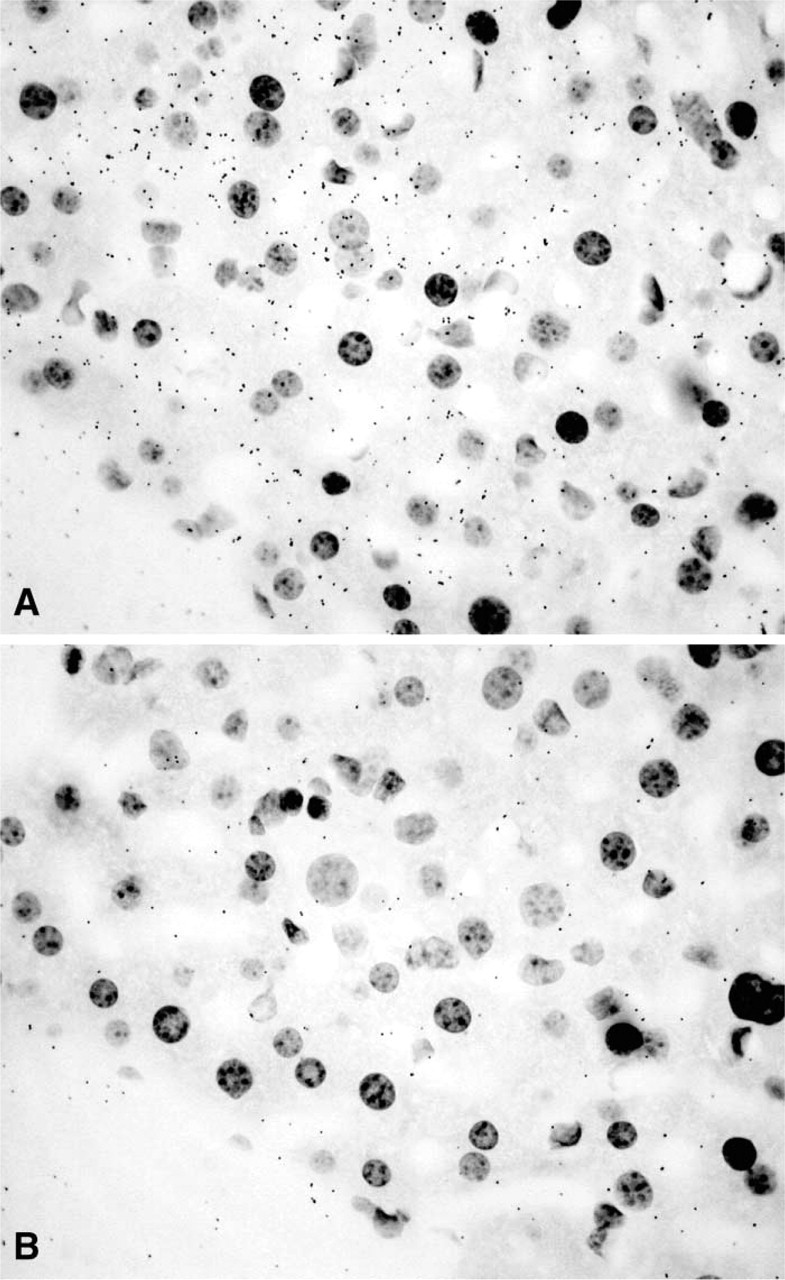
After 3–8-day exposure of the films, specific labeling was found in the ovary, uterine cervix, mammary gland, testis, skin, adrenal cortex, liver, and kidney. No specific hybridization signal could be detected in the vagina, prostate, pituitary, jejunum, and pancreas. From photographic emulsion-coated sections, information could be obtained about the cell types expressing 20α-HSD mRNA in the different tissues. In the ovary, as shown in X-ray films and emulsion-coated slides after a short exposure time (7 days), labeling was restricted to corpora lutea (Figure 1 and 2). Because the mice were in diestrous day 1, only old corpora lutea were present. Cell counting performed on at least two corpora lutea per ovary (four mice) established that approximately 30% of luteal cells were labeled. At longer exposure times (28 days), weak specific labeling was also observed in granulosa cells of growing follicles at all stages of development (Figure 3).
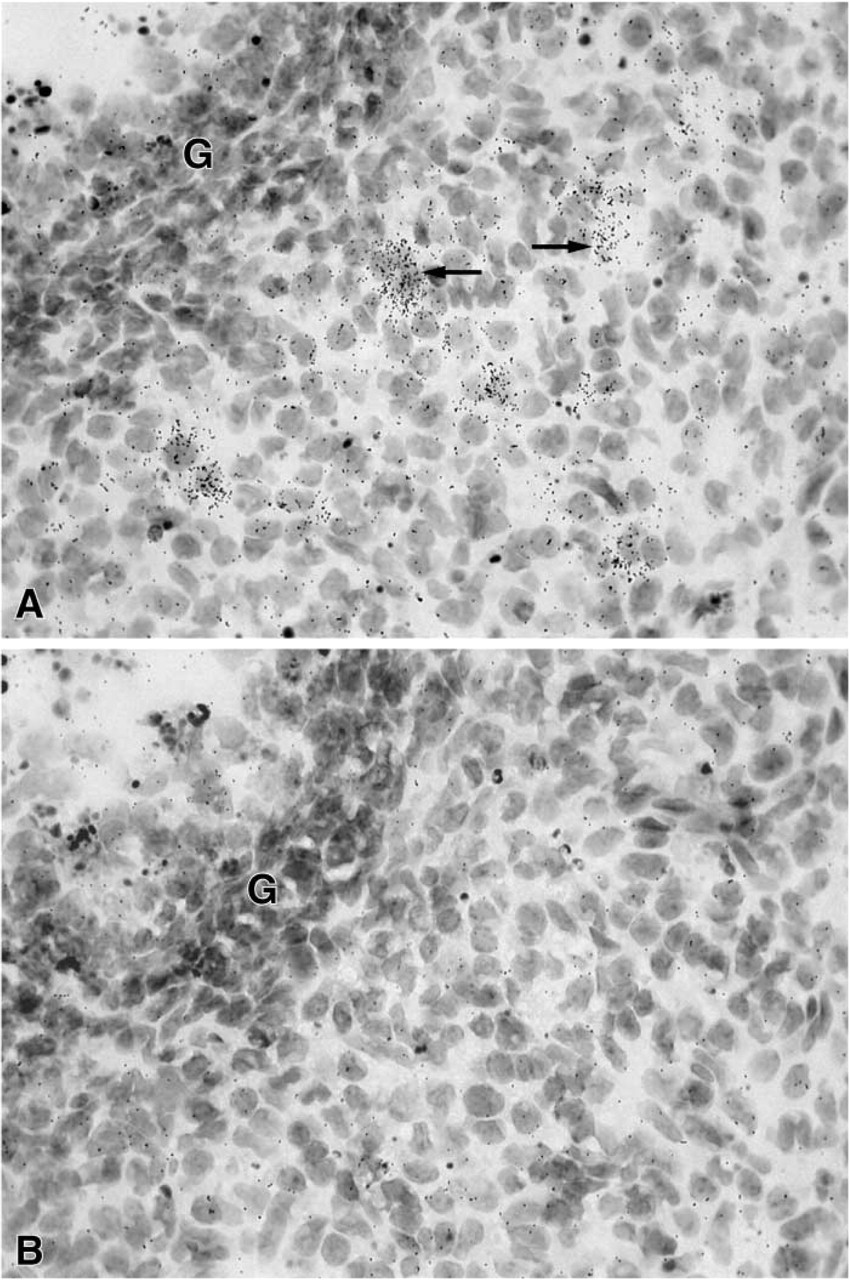
(
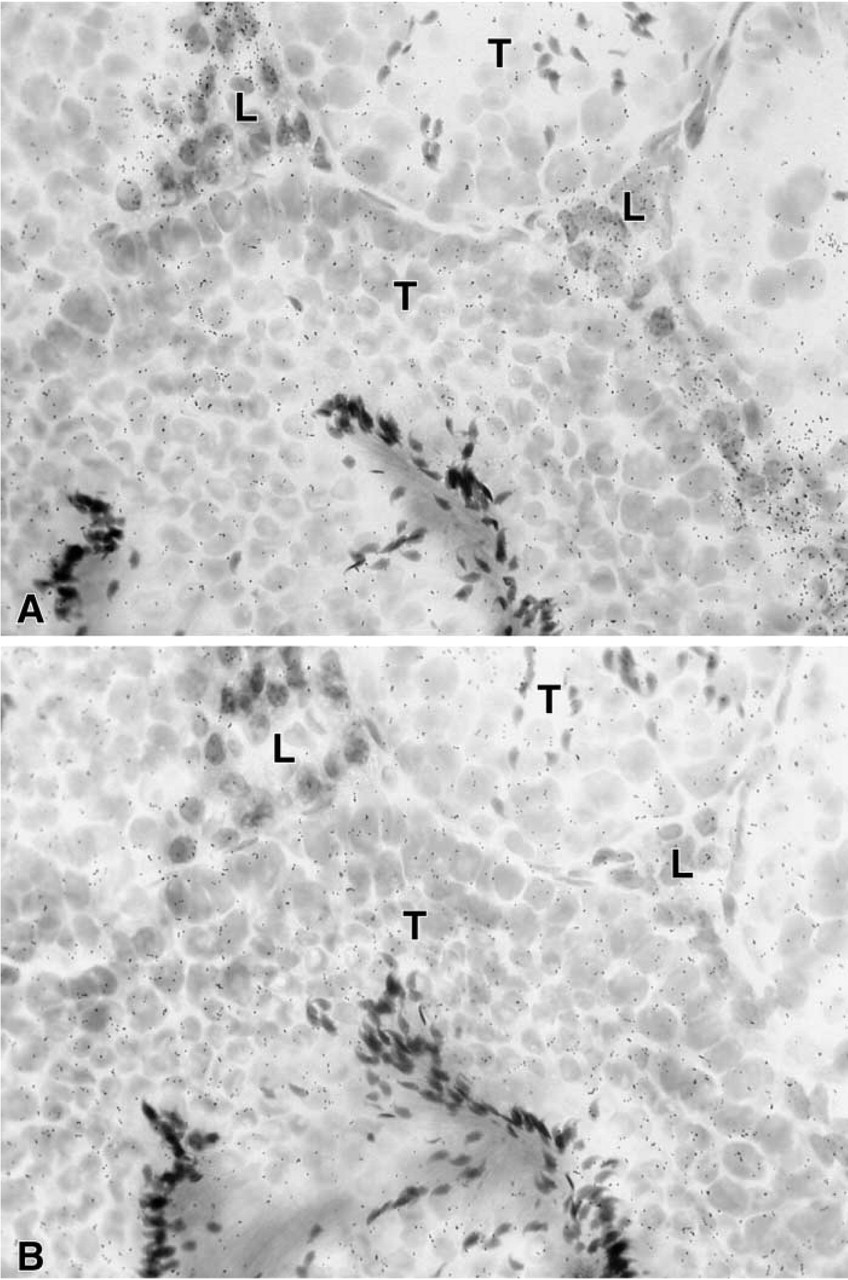
In the uterine cervix, the hybridization signal was found in both endocervix and exocervical epithelial cells, the stroma being devoid of any specific labeling (Figure 4). In the female mouse mammary gland, weak specific labeling was restricted to stromal cells surrounding ducts (Figure 5). Typical acini could not be observed. In the male mouse mammary gland, no specific labeling could be detected in any cell type at the longest time interval studied (45 days). In the testis, a specific hybridization signal was found only over Leydig cells, the tubules being devoid of any specific reaction (Figure 6).
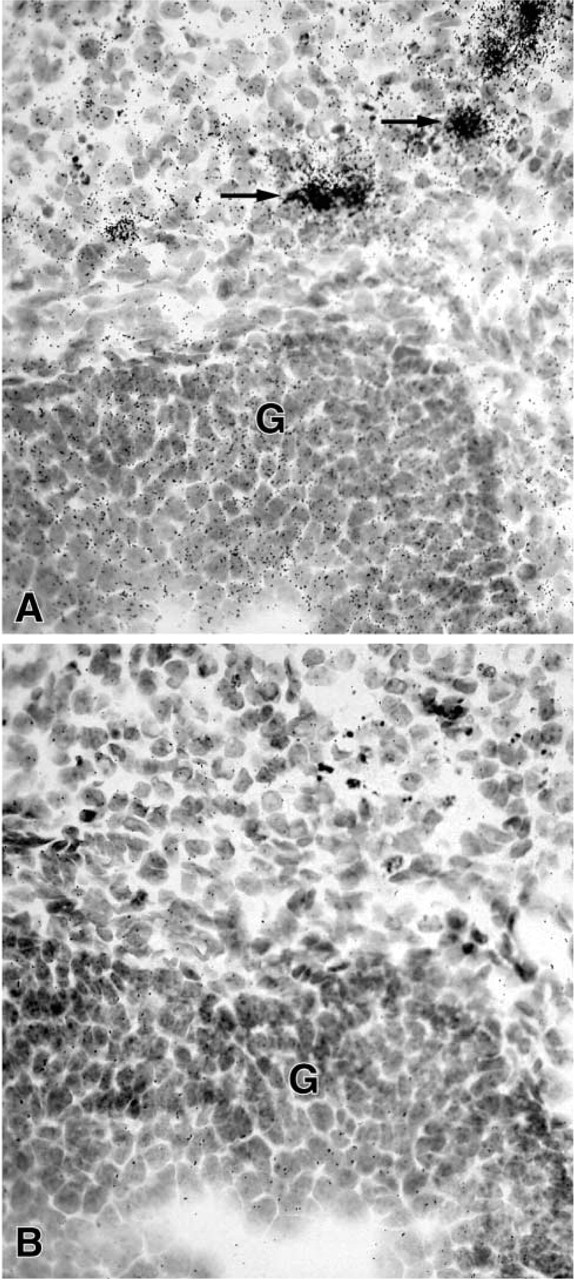
(
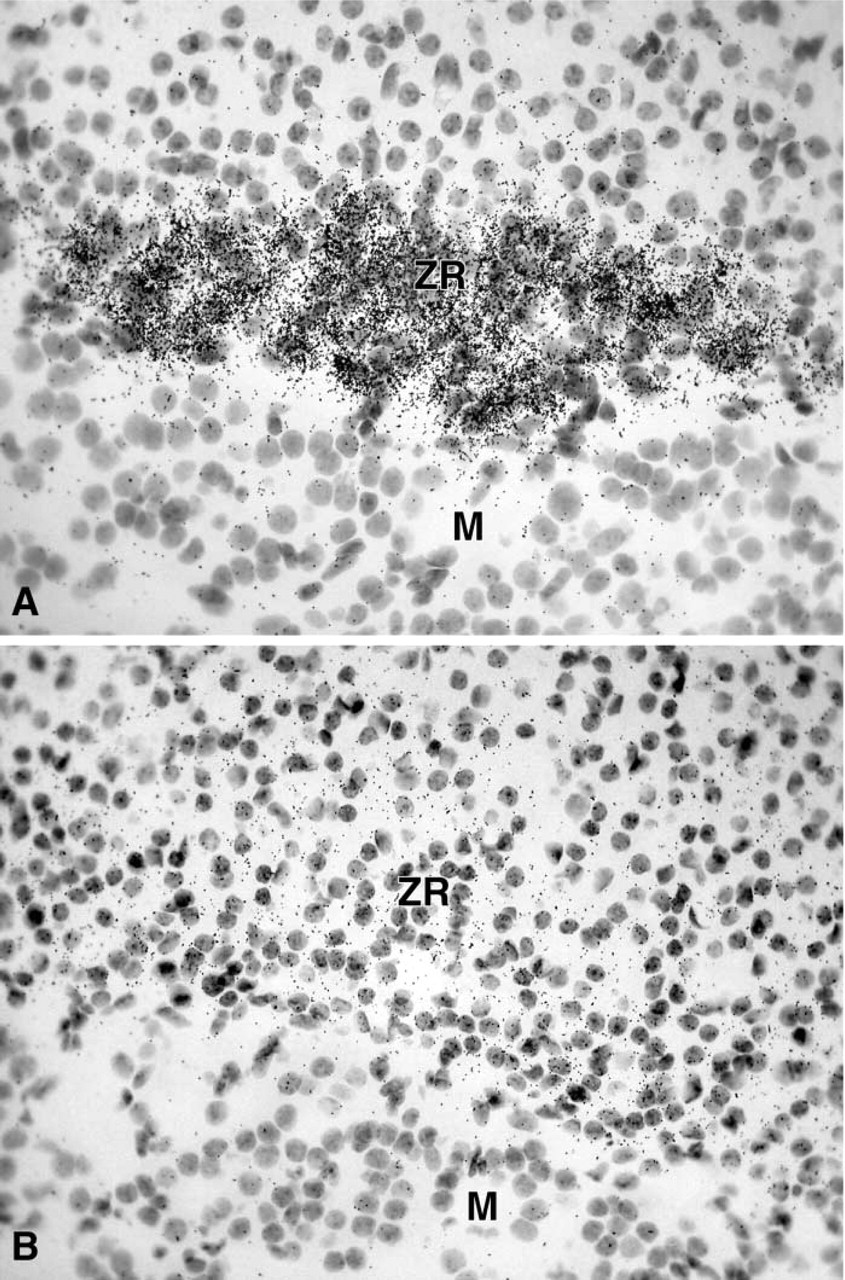
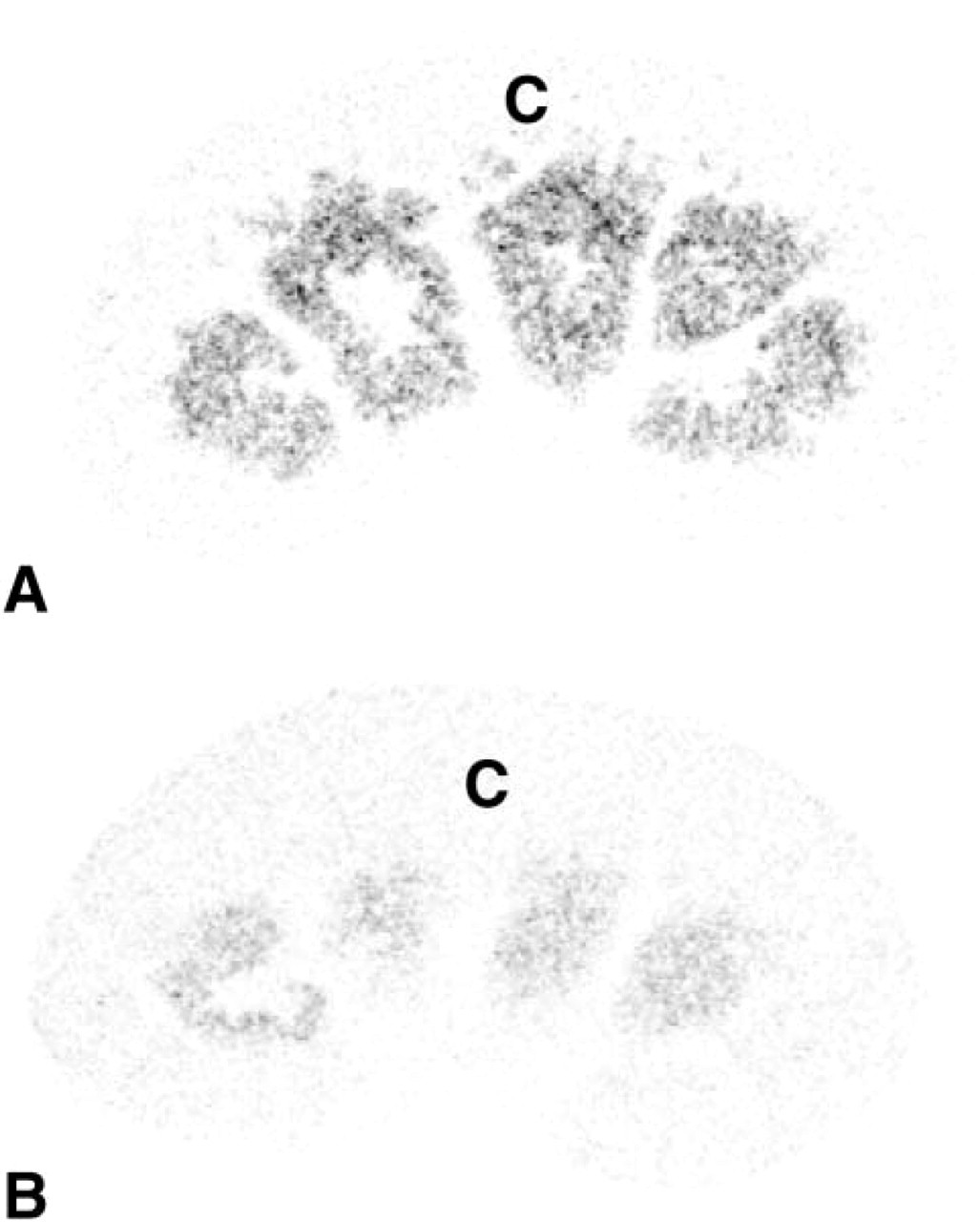
In female adrenal glands, strong labeling was obtained in the zona reticularis of the adrenal cortex as early as after 1 day of exposure time, whereas the other zones of the cortex and the adrenal medulla were totally negative (Figure 7A). In the male, only weak labeling could be detected in the zona reticularis of the adrenal cortex after the longest exposure time (45 days) (Figure 7B).
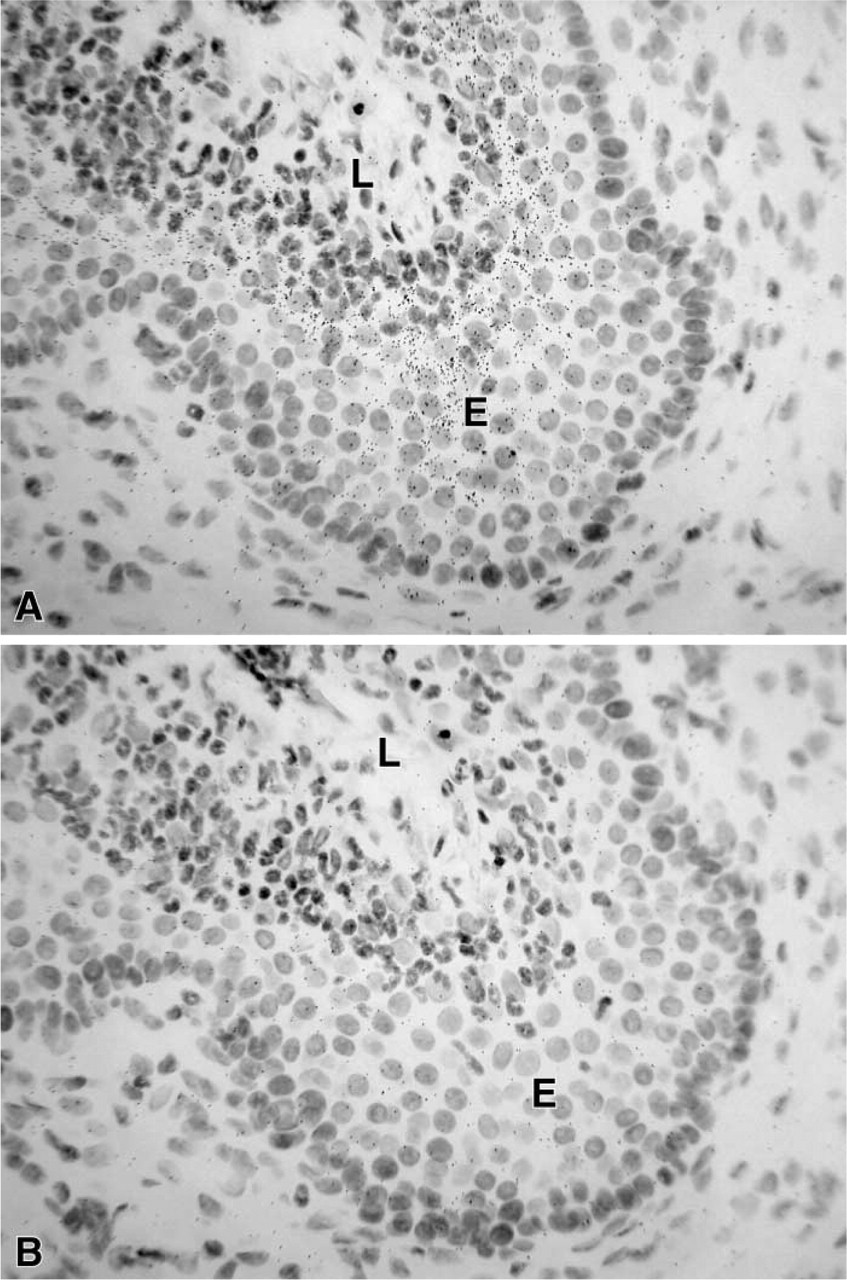
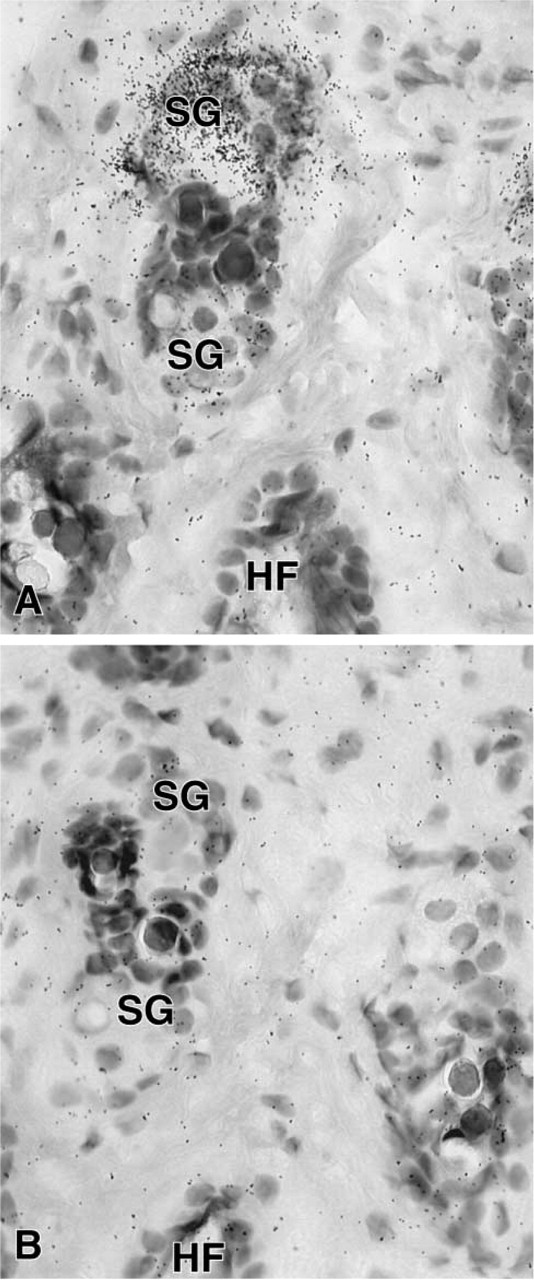
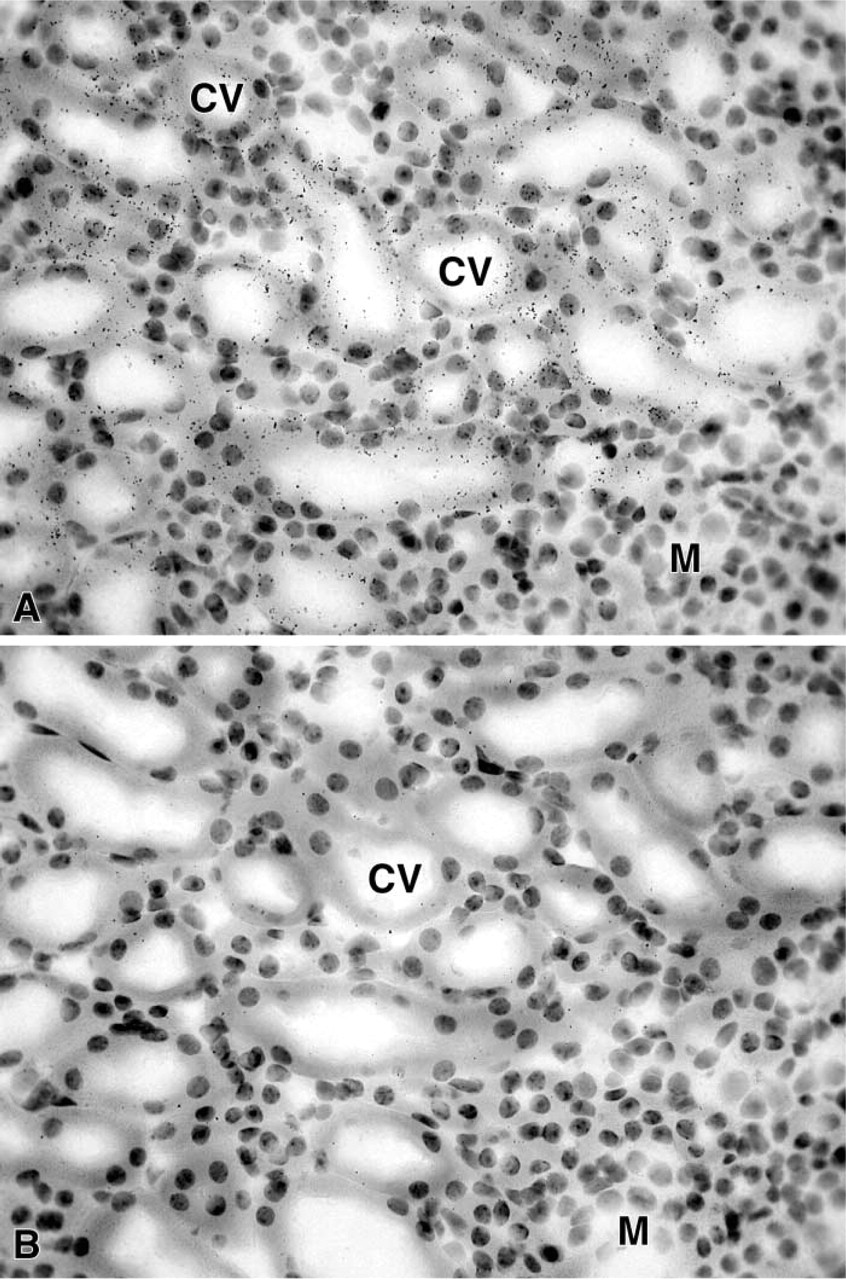
In the female mouse skin, a specific hybridization signal was detected in the sebaceous gland cells after 7 days of exposure (Figure 8), but the epidermis, stromal cells, and hair follicles were totally unlabeled. In the male, only a weak signal could be obtained in the sebaceous glands after a long-term exposure (not shown). In the liver, 20α-HSD mRNA appeared to be expressed in both male and female animals, with a higher signal in the female (Figure 9). As observed by light microscopy, specific labeling was detected in all the hepatocytes (Figure 10). In the kidney, specific labeling was observed in a large area located between the medulla and the outer part of the cortex (Figure 11). As observed for the other tissues, the expression of the enzyme mRNA was higher in the female than in the male (Figure 11). At the light microscopic level, we observed that silver grains were overlying the epithelial cells lining the distal convoluted tubules, whereas the glomeruli, proximal convoluted tubules, collecting tubules, and blood vessels were unlabeled (Figure 12). In all the tissues examined, hybridization with the sense probe generated only a light uniform labeling (for examples, see Figures 1, 2, 4, 5, 6, 8, 10, and 12).
Discussion
The enzyme 20α-HSD plays an important role in the regulation of the intracellular concentration of progesterone. Cloning of the cDNA encoding mouse 20α-HSD has allowed studies on the histological localization of 20α-HSD mRNA in a series of male and female mouse tissues. In the ovary, strong labeling was observed in about 30% of luteal cells in corpora lutea, while a much weaker signal was obtained in granulosa cells of growing follicles. These findings support the data indicating that, in the rat ovary, 20α-HSD activity was much higher in luteal than in non-luteal cells (Seong et al. 1992). 20α-HSD plays a pivotal role in the regulation of progesterone secretion by corpora lutea. It is generally admitted, however, that there are two types of luteal cells: the luteinizing granulosa cells, which have a large cytoplasm with small lipid droplets, and the theca lutein cells, which are smaller but contain large lipid droplets. It was not possible using frozen sections to establish which luteal cell type(s) was involved in 20α-HSD mRNA expression. On the other hand, it has been shown that, in the rat ovary, all the luteal cells express the enzyme 3β-hydroxysteroid dehydrogenase and can therefore be involved in progesterone secretion (Dupont et al. 1990). In the rat, prolactin strongly inhibits the expression of 20α-HSD, thus contributing to maintenance of progesterone at high levels during pregnancy (Albarracin et al. 1994). The role of 20α-HSD weakly expressed in granulosa cells remains to be clarified.
(
In the female reproductive tract, 20α-HSD mRNA expression was detected only in the epithelial cells of the uterine cervix. The role of the enzyme might be related to the delivery process. It has been reported that, in mice lacking 5α-reductase type 1, an enzyme that is also involved in progesterone catabolism and is expressed in the cervix, there was a failure of cervical ripening, leading to abnormal delivery (Mahendroo et al. 1999). Such data suggest that 20α-HSD activity does not play a major role in the catabolism of progesterone in the cervix, at least during parturition.
(
In the mammary gland of the female mouse, weak 20α-HSD expression was detected only in stromal cells surrounding the ducts. Progesterone receptors have been found to be expressed in both epithelial cells of the ducts and stromal cells in the mouse mammary gland (Fenn et al. 1998). It therefore appears that progesterone metabolism is mainly restricted to stromal cells in mammary glands of the cycling mouse.
(
In the mouse testis, the hybridization signal was observed only in Leydig cells. The testicular localization of the enzyme has already been reported in the bovine and human species (Warren et al. 1993; Zhang et al. 2000). The low expression of 20α-HSD mRNA reported in the human testis (Zhang et al. 2000) can be explained by the fact that the expression of the enzyme is restricted to Leydig cells, which represent only a small proportion of the total number of testicular cells. The role of 20α-HSD might be related to some protective effect from progesterone in testicular tissue.
In the adrenal glands, 20α-HSD mRNA expression was restricted to the cells of the zona reticularis. The hybridization signal was very high in the female and very low in the male. From pig adrenal cytosol, Nakajin et al. (1989) have purified two proteins exhibiting 20α-HSD activities, which they named 20α-HSD I and II. We have recently reported low expression of 20α-HSD mRNA in total adrenal glands in whole human adrenal glands (Zhang et al. 2000). This low expression of 20α-HSD mRNA might be explained by the restricted expression of 20α-HSD mRNA to the zona reticularis, which consists of a few layers of cells and represents a very small proportion of the total number of adrenal cells. The physiological role of 20α-HSD in the adrenal cortex remains to be clarified.
(
In the skin, 20α-HSD mRNA was strongly expressed in sebaceous glands, but no specific labeling could be detected in the hair follicle and stromal cells or in the epidermis. The presence of progesterone receptors has been reported in sebaceous glands in the human (Wallace and Smaller 1998). The enzyme might therefore regulate the availability of circulating and locally produced progesterone for progesterone receptors and thus control the influence of progesterone on sebaceous gland cell activity. Because it has been shown that progesterone treatment suppresses estrogen receptors in the monkey sex skin (West et al. 1990), one role of progesterone might be to regulate estrogen receptor levels that are expressed in sebaceous glands (Pelletier 2000). The very low expression of 20α-HSD mRNA in the male mouse skin is indicative that 20α-HSD mRNA might be positively regulated by ovarian hormones. In fact, we have recently observed that ovariectomy induced a striking decrease in 20α-HSD mRNA levels in mouse skin (unpublished data).
(

X-ray autoradiograph showing localization of 20α-HSD mRNA in sections through the liver of a female (
In the kidney, a specific hybridization signal was found in the epithelial cells in the distal convoluted tubules. As in other tissues, the signal was more striking in the female than in the male. In the human kidney, the expression of 20α-HSD has been reported (Rumke-Vogl et al. 2002) but there was no indication about the cell types involved in the expression of the enzyme. Interestingly, progesterone receptors have been found in epithelial cells of distal convoluted tubules (Rumke-Vogl et al. 2002). From the results obtained in the mouse and human kidney, it appears that the same epithelial cells can express both progesterone receptors and the enzyme that inactivates progesterone. 20α-HSD might be involved in the fine regulation of the action of progesterone, which has been shown to be a potent mineralocorticoid receptor antagonist (Myles and Kuner 1996). In the liver, 20α-HSD mRNA expression was observed in the hepatocytes, the expression being also higher in the female. Because of the large number of hepatocytes, such data indicate that liver might be considered as a major site of progesterone catabolism.
(
X-ray autoradiograph showing kidney sections from a female (
The absence of any hybridization signal in the prostate, uterus, vagina, and pituitary might be explained by a low expression of 20α-HSD that could not be detected by ISH in these tissues. We might also consider the possibility that, in these four tissues, a fine regulation of intracellular progesterone concentrations is not critical for hormonal activity. By RT-PCR, Zhang et al. (2000) have shown low expression of 20α-HSD mRNA in human prostate and uterus, whereas in the present studies we were unable to detect any hybridization signal in the same mouse tissues. These apparent species differences might be explained by a lower sensitivity of ISH compared to that obtained with the highly sensitive RT-PCR technique.
In summary, the present results on the localization of 20α-HSD mRNA in mouse tissues indicate that the enzyme is highly expressed in the male and female gonads and in the uterine cervix. In non-reproductive tissue, the expression of 20α-HSD mRNA is always higher in the female than in the male, thus suggesting that the enzyme plays a predominant role in the inactivation of locally produced or circulating progesterone, which is much higher in the female. Other studies involving ovariectomy and hormone replacement therapy are required to clarify the role of ovarian steroids in the regulation of 20α-HSD expression.
(
Footnotes
Acknowledgements
The ATLAS program was supported by Genome Canada and Genome Québec.