
Abstract
We present a novel procedure for detection of low- and high-abundance messenger RNAs in the brain by in situ hybridization histochemistry, by using fragmented double-stranded cDNA as molecular probes. The procedure involves digesting the cDNA of interest with the restriction endonuclease from Desulfocibrio desulfuricans (Dde I digestion), followed by random primed labeling, which generates a family of high specific activity cDNA fragments. This procedure is a rapid, straightforward, and reproducible method of obtaining sensitive probes for in situ hybridization and is generally applicable to the analysis of the expression of a large number of genes. Here we report the use of this procedure to prepare probes for the detection of synapsin I, p150Glued, neurotensin, c-fos, and c-jun mRNAs in brain, using both isotopic and non-isotopic labeling methods. Because this procedure does not require complex recombinant DNA manipulations or oligonucleotide design, it should prove useful to the non-molecular biologist examining the expression of genes in the central nervous system.
I
Alternatively, to circumvent the time-consuming and tedious nature of cRNA probe production, labeled single-stranded synthetic oligonucleotides can be used as molecular probes (Caruthers 1985). The use of oligonucleotide probes, however, also presents unique technical concerns. For example, the design of effective oligonucleotide probes requires the aid of predictive computer analysis and, even when such software is utilized to optimize probe design, the synthesis or purchase of the many oligonucleotides can be prohibitively expensive. Furthermore, the typical oligonucleotide probe of 30–50 bases contains labeled nucleotides only on the 5' end (Coghlan et al. 1985; Wolfson et al. 1985) or 3' tail (Lewis et al., 1985, 1986a,b), often resulting in hybridization signals that are usually significantly less compared to a family of internally labeled variable-length or single- or double-stranded nucleotide probes.
The use of double-stranded cDNAs as molecular probes for in situ hybridization to chromosomal preparations has been effective for localizing the position of a particular gene (Fostel et al. 1984; Pardue and Dawid 1981). However, their use to detect and localize mRNA expression in brain tissue has been constrained because of sensitivity issues. In general, cDNA probes are labeled by nick-translation (Rigby et al. 1977). In this procedure, the double-stranded phosphate backbone of the cDNA is “nicked” by incubation in dilute concentrations of the nonspecific DNA-cleaving enzyme DNAse I and then is repaired in the presence of radioactive- or chromogen-labeled nucleotide bases using the large fragment of DNA polymerase I (Klenow fragment), yielding a labeled set of probe molecules. Because the intrinsic limitations of in situ hybridization require that cDNA fragments radiolabeled by the nick-translation method have an average length of 100–300 BP, this goal can require considerable effort in optimizing DNAse digestion conditions. Recently, rapid progress in technique development in the field of in situ hybridization has led to reports describing the use of short cDNA probes generated by polymerase chain reaction (PCR) using Thermus aquaticus DNA (Taq DNA) polymerase (Fuhrman and Fayein 1994; Brooks et al. 1993). Although these procedures offer alternative techniques for generating short cDNA probes for use in in situ hybridization histochemistry in tissue sections, they present additional technical barriers that may preclude their use on a broader scale. Some of the issues include uncertainties and expense associated with DNA oligonucleotide primer design and purchase, subcloning of cDNA fragments into suitable vectors, and access to a thermal cycling unit (PCR machine). The overall applicability of such PCR-generated probes is potentially complicated by problems of efficacy and sensitivity when these methods are applied to a broader spectrum of target genes.
We sought a straightforward approach to produce double-stranded cDNA probes for detection of low- and high-abundance cellular RNAs by in situ hybridization histochemistry, with particular emphasis on the study of gene expression by in situ hybridization within the CNS. The procedure relies on the digestion of the cDNA of interest with the high-frequency recognition site restriction endonuclease Dde-I to produce a series of cDNA fragments specific for a particular target mRNA sequence. After random prime labeling (Feinberg and Vogelstein 1983), this method effectively produces a “family” of small cDNA probes well suited for in situ hybridization. This procedure is a reproducible method of obtaining sensitive probes for in situ hybridization studies and is readily applicable to the analysis of the expression of a large number of genes. Furthermore, this procedure does not require complex recombinant DNA manipulations and avoids the design and expense of custom-synthesized oligonucleotides. We report here the detection of five mRNAs in the nervous system using Dde I-digested cDNA probes labeled by random primed labeling. Synapsin I mRNA was detected in neurons in the developing CNS and PNS. p150Glued mRNA was detected in neurons in the developing rat CNS and PNS, and in cells throughout the developing embryo. c-fos and neurotensin/neuromedian N mRNAs were detected in the rat dorsolateral striatum (DLS), and c-jun mRNA was detected in the rat hippocampus.
Materials and Methods
Animals, Experimental Procedures, and Tissue Preparation
For detection of endogenous levels of synapsin I and p150Glued mRNAs in developing rat embryos, Sprague-Dawley rat pups (n = 3–4 per time point) (Charles River Breeding Laboratories; Wilmington, MA) from embryonic (E) Days 16 and 19 were used in this study. For in situ hybridization on embryonic rat tissue, rat pups were frozen by immersion in dry-ice-supercooled 2-methyl butane (Aldrich; Milwaukee, WI), warmed to - 16C in a cryostat, and sectioned whole at 16 μm in the sagittal plane. Sections were thaw-mounted on precooled slides coated with Vectabond reagent (Vector Laboratories; Burlingame, CA) and stored at - 70C.
For detection of mRNAs encoding c-fos and neurotensin/neuromedian N (NT/N) in brain after neuroleptic treatment, several adult male Sprague-Dawley rats (Charles River Breeding Laboratories), 90–120 days old and weighing 250–300 g, were treated with a single intraperitoneal (IP) injection of haloperidol (1 mg/kg; Sigma Chemicals, St Louis, MO). At various times after treatment (i.e., 0.0, 0.5, and 3.0 hr), rats were sacrificed by carbon dioxide asphyxiation followed by decapitation. The brains were rapidly removed, divided sagittally into two halves, and then frozen and sectioned as above in the coronal plane.
For detection of endogenous levels of c-jun mRNA in brain, other adult male Sprague-Dawley rats were sacrificed and the brains removed and processed as described above.
Dde I Restriction Fragment Analysis
The cDNA used to generation synapsin I probe was 1700-BP EcoRI fragment, 5E2 [nucleotides 694–2445 of the full length synapsin I cDNA clone, pSyn 5 from rat (Kilimann and DeGennaro 1985)]. The cDNA used to generate p150Glued probe was a full-length 4.4-KB cDNA from rat (Holzbaur et al. 1991). The cDNA used to generate NT/N probe was a full-length 686 BP cDNA from rat (Alexander et al. 1989). The cDNA used to generate c-fos probe was 2.1-KB EcoRI fragment [nucleotides 1–2140 of the full-length c-fos cDNA clone, pCMV-c-Fos from mouse (Harrison et al. 1995)]. The cDNA used to generate c-jun probe was an 1800-BP EcoR1-Pst-1 fragment of pCMV2c-Jun (Harrison et al. 1995).
The cDNA fragments were generated with the restriction endonuclease Dde I (Boehringer Mannheim; Indianapolis, IN), from Desulfovibrio desulfuricans. Dde I has a four nucleotide base recognition sequence (C/TNAG), and therefore typically recognizes and digests at a high frequency along any given length of DNA. Briefly, 50 μg each of the cDNAs mentioned above were digested with 50 U of Dde I in a high-salt buffer (Buffer H: 50 mM Tris-HCl, 10 mM MgCl2, 100 mM NaCl, 1 mM dithioerythritol, pH 7.5) for 24 hr at 37C. The restriction products were then purified by ethanol precipitation by the addition of 1/10 volumes of 3 M sodium acetate and 2.2 volumes ice-cold 100% ethanol for 2 hr at -20C. Restriction fragments were resuspended to a final concentration of 1 μg/ml in distilled water, and aliquots of the Dde I-digested cDNAs were separated by agarose gel electrophoresis (4% agarose in 40 mM Tris acetate, 10 mM EDTA, pH 8.3) to monitor Dde I digestion (Figure 1).
Preparation of cDNA Probes
For in situ hybridization histochemistry, Dde I-digested cDNAs were labeled in the presence of either [α-35S]-dCTP, [α-32P]-dCTP, [α-33P]-dCTP, or [α-digP]-dUTP (BMB, Indianapolis, IN) by random primed labeling (Boehringer Mannheim). As an example, 500 ng-1 μg cDNA in 10 μl of water was denatured for 5 min at 95C. After denaturation, the cDNA was immediately cooled on ice; 2 μl 10 X reaction mixture [500 mM potassium phosphate, 2.5 mM dithiothreitol, 50% glycerol (v/v), pH 7.0], 1 μl each of the appropriate cold nucleotide-typically 10 mM dATP, dGTP, dTTP, and 5 μl of [α-35S]-dCTP (50 μCi) (3000 Ci/mmol), and 1 μl sequencing grade DNA polymerase I (5U) [Klenow fragment] were added sequentially on ice. The reaction mixture (20 μl total volume) was incubated for 45 min at 37C and was terminated by addition of 2 μl of 0.2M EDTA. Unincorporated labeled nucleotides were removed by chromatography over a Sephadex G25 spin column (Boehringer Mannheim). Efficiency of cDNA synthesis and labeling was determined by incorporation of [α-35S]-dCTP, [α-32P]-dCTP, or [α-33P]-dCTP, in the reaction mixture. For digoxigenin-labeled probes, a slot-blot was performed to assess the quality and specific activity of probe produced and was compared to a standard provided by the manufacturer and to previous probe batches that had performed well under experimental conditions.
In Situ Hybridization
Slide-mounted brain sections were warmed to room temperature (RT), postfixed in 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) for 5 min at 4C, rinsed in PBS, and treated with 0.25% acetic anhydride (in 0.1 M triethanolamine, pH 8.0) for 10 min at RT. After rinsing in 2 X SSC and dehydration through a graded series of alcohols, the sections were delipidated in chloroform for 5 min at RT. The sections were subsequently rehydrated to 95% ethanol in descending concentrations of alcohols and then air-dried.
Isotopic Detection. For hybridization, radiolabeled cDNA probe (2–4 × 106 cpm) was applied in 65 μl of hybridization buffer [50% formamide, 10% dextran sulfate, 2 X Denhardt's solution (1 X Denhardt's solution is 0.02% Ficoll, 0.02% polyvinylpyrrolidone, 0.02% BSA), 5 X SSC, 50 mM DTT, 0.1% SDS, 100 μM dNTPs (i.e., ATP, dTTP, dGTP, and dCTP), and 0.1% Na4P2O7], and the slides were cover-slipped and incubated at various temperatures in a moist chamber for <12 hr. Synapsin I cDNA probes were incubated at 50C, p150Glued cDNA probes at 45C, and NT/N cDNA probes and c-fos at 42C. c-jun cDNA probes were incubated at 55C. After hybridization, coverslips were removed in 2 X SSC, 0.1% Na4P2O7 at RT. Slides were washed for 30 min each in two changes each of 2 X SSC, 0.1% Na4P2O7 at RT, 2 X SSC, 0.1% Na4P2O7 at 42C, 0.5 X SSC, 0.1% Na4P2O7 at RT, 0.1 X SSC, 0.1% Na4P2O7 at RT, and 0.1 X SSC, 0.1% Na4P2O7 at 42C. A final wash in 0.1 X SSC, 0.1% Na4P2O7 for 15 min at RT was done, and the sections were dehydrated through a graded series of alcohols in which water was replaced by 0.6 M ammonium acetate. The slides were then air-dried and exposed to Kodak XAR-5 film (Eastman Kodak; Rochester, NY) for 7–14 days at RT.
Non-isotopic Detection. For prehybridization, sections were incubated in 4 X SSC, 1 X Denhardt's solution at 37C for 1 hr in a moist chamber. For hybridization, digoxigenin-11-dUTP labeled cDNA probe (20 ng) was applied in 150 μl hybridization buffer (50% formamide, 600 mM NaCl, 4 mM EDTA, 0.1% Na4P2O7, 0.2% Sarkosyl, 10 mM DTT, 80 mM Tris-Cl, pH 7.5), and the slides were coverslipped and incubated at 37C in a moist chamber for at least 12 hr. After hybridization, coverslips were removed in 2 X SSC at RT. Slides were washed for 30 min each in two changes of 2 X SSC preheated to 37C and then for 30 min each in three changes of 0.1 X SSC also preheated to 37C. Sections were then immersed in 1 X PBS (0.001 M KH2PO4, 0.01 M Na2HPO4, 0.137 M NaCl, 0.0027 KCl, pH 7.4) for 1–2 hr at 4C. For immunological detection, slides were rinsed in Buffer A [1 M NaCl, 2 mM MgCl2, 2% fetal goat serum (Sigma Chemicals), 0.1% Triton X-100, 0.1 M Tris-Cl, pH 7.5] for 30 min at RT and then incubated with anti-digoxigenin antibody (Boehringer Mannheim) diluted 1:5000 in Buffer A for up to 24 hr at 4C. Slides were then washed for 10 min each in three changes of Buffer A, followed by one wash for 5 min each in Buffer B (1 M NaCl, 5 mM MgCl2, 0.1 M Tris buffer, pH 9.5) and Buffer C (0.1 M NaCl, 5 mM MgCl2, 0.1 M Tris buffer, pH 9.5). The color reaction was then carried out in the dark by incubation in NBT and X-phosphate in Buffer C for 6–12 hr at RT. The color reaction was stopped in 1 X PBS, and the slides were coverslipped with Aqua-Mount (Polysciences; Warrington, PA).
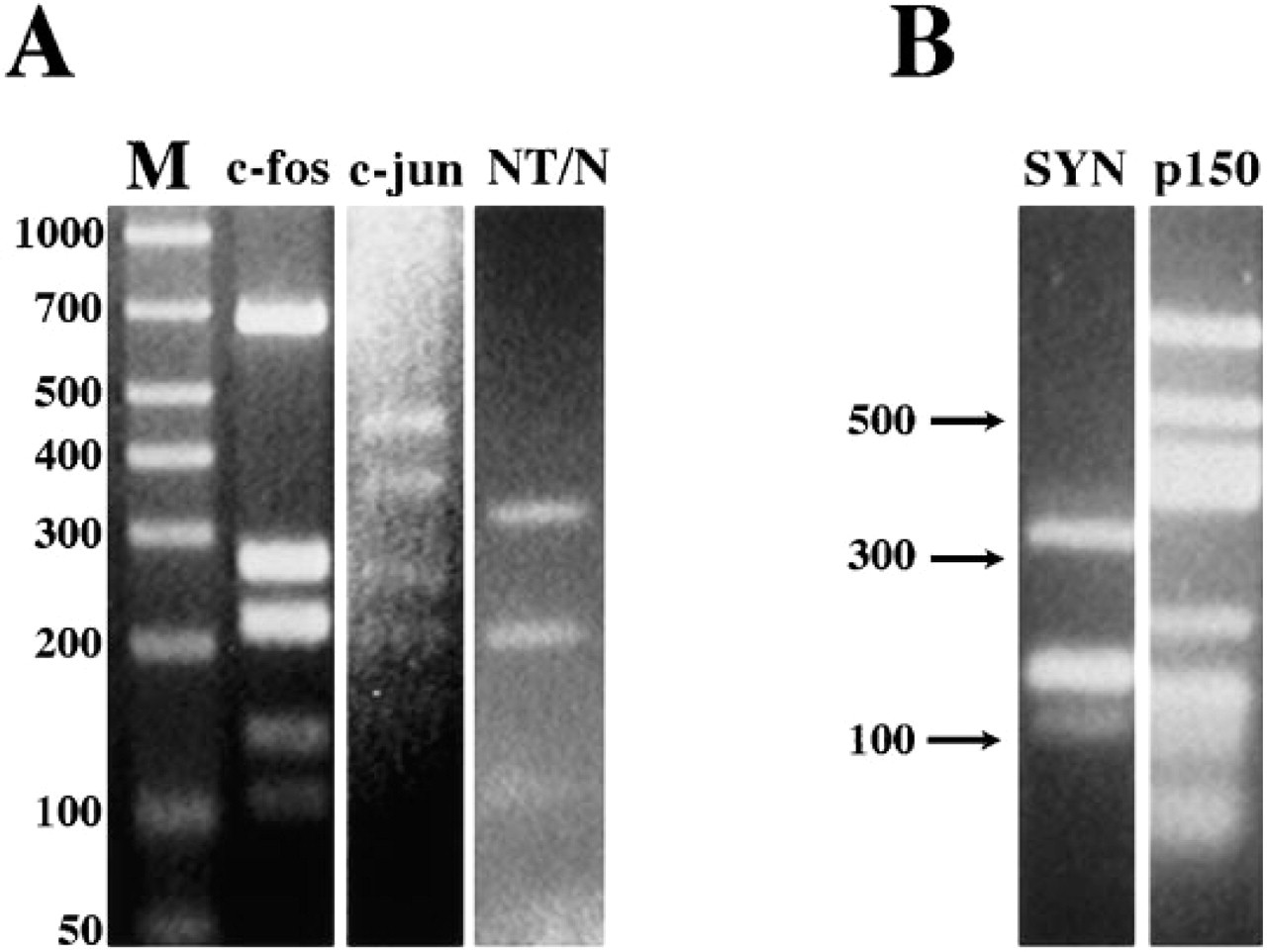
Agarose (4%) gel electrophoresis detailing the pattern of Dde-I digestion of synapsin I, p150Glued, c-fos, c-jun, and neurotensin/neuromedian N cDNAs.
Controls for Specificity
The specificity of the probe for target mRNAs in situ was evidenced by several controls: (a) no hybridization was observed in tissue sections pretreated with RNAse A (10 μg/μl; (b) no hybridization was observed in the absence of labeled cDNA probe in the hybridization reaction; (c) under the conditions described above, no hybridization was observed in the presence of a 100-fold excess of homologous unlabeled Dde I-digested cDNA in the hybridization reaction.
Results
Isotopic Detection of Synapsin I and p150Glued mRNAs in the Rat Embryo Using Dde I-digested cDNA Probes
Synapsin I is the best characterized member of a family of nerve terminal-specific phosphoproteins believed to play an integral role in the process of neural transmission. p150Glued is a component of the dynactin (Glued) complex, which in vitro has been shown to be a required activator of cytoplasmic dynein-mediated transport of vesicles along microtubules, and therefore has been postulated to be an essential component of retrograde transport mechanism. Figure 2 presents a comparison of the use of Dde I-digested cDNA probes to examine the pattern of expression synapsin I and p150Glued mRNAs in developing rat embryo and brain by in situ hybridization histochemistry. Synapsin I mRNAs were localized to neurons within CNS and PNS tissue (Melloni and DeGennaro 1994; Melloni et al. 1993, 1994). At embryonic day 16 (E16), synapsin I mRNAs were detected by Dde I-digested synapsin cDNA probes throughout the CNS and PNS (Figure 2A). Areas high in synapsin I mRNA by E16 were the hypothalamic and thalamic neuroepithelium, the brainstem, the dorsal root ganglion (DRG), and the developing spinal cord (Sp Cord). By comparison, at E16 the hybridization of Dde I-digested p150Glued cDNA probes displays a more ubiquitous expression (Figure 2B; and Melloni et al., 1995). From the earliest developmental time points examined, a low level of hybridization was observed in cells throughout the developing embryo (Figure 2B and D). In agreement with data published on the expression of the polypeptide (Holzbaur et al., 1989, 1991), p150Glued mRNA expression is apparently enriched in neuronal tissue, because the most intense levels of hybridization to the p150Glued probe were observed in neuronal cells. Particularly high expression was observed in the spinal cord, dorsal root ganglion, and the olfactory epithelium of the developing rat embryos at Day E16.
By embryonic Day 19, Dde I-digested synapsin I and p150Glued cDNA probes hybridized to neurons, forming distinct nuclei and recognizable tracts of cells in the rat brain (Figures 2C and D). Labeling of synapsin I mRNA appeared high in the hippocampus, the piriform cortex, and the thalamus (Figure 2C). In addition, synapsin I mRNA was detected at low levels by Dde I-digested cDNA probes in the developing retina (Figure 2C). Areas notably high in hybridization to Dde I-digested p150Glued cDNA probes included the olfactory epithelium, neuroepithelial layer of the retina, the hippocampus, the piriform cortex, and the thalamus (Figure 2D).
Isotopic Detection of Haloperidol-induced c-fos and Neurotensin/Neuromedian N mRNAs in Adult Rat Brain Using Dde I-digested cDNA Probes
Neurotensin (NT) is a tridecapeptide with a widespread distribution in the CNS and in the gastrointestinal tract, where NT affects gut motility and perhaps serves as growth factor (Alexander et al. 1989; deNadai et al. 1989; Carraway and Mitra 1987; Jennes et al. 1982). In the CNS, NT is likely to function as a neurotransmitter or neuromodulator, because binding of NT to the neurotensin receptor causes changes in cyclic nucleotide levels and inositol phosphate metabolism (Gully et al. 1993; Vita et al. 1993; Tanaka et al. 1990). Neurotensin peptide has been shown to have a close anatomic and biochemical association with the dopaminergic systems in the basal ganglia, supporting the notion that NT may play a role in modulating dopaminergic pathways (Augood et al. 1991; Ervin and Nermeroff 1988). A striking induction of NT/N mRNA in the dorsolateral striatum has been reported in rats after treatment with the neuroleptic drug haloperidol, i.e., a dopamine D2 receptor antagonist (Merchant and Miller 1994; Merchant et al. 1992). In addition to the induction of NT/N mRNA, a parallel yet temporally distinct induction of the cellular immediate early gene c-fos was also observed (Merchant and Dorsa 1993). c-Fos induction has been widely utilized as a marker for neuronal activity and recently has been shown to be important for transcriptional response of the NT/N gene to environmental stimuli in vitro (Harrison et al. 1995).
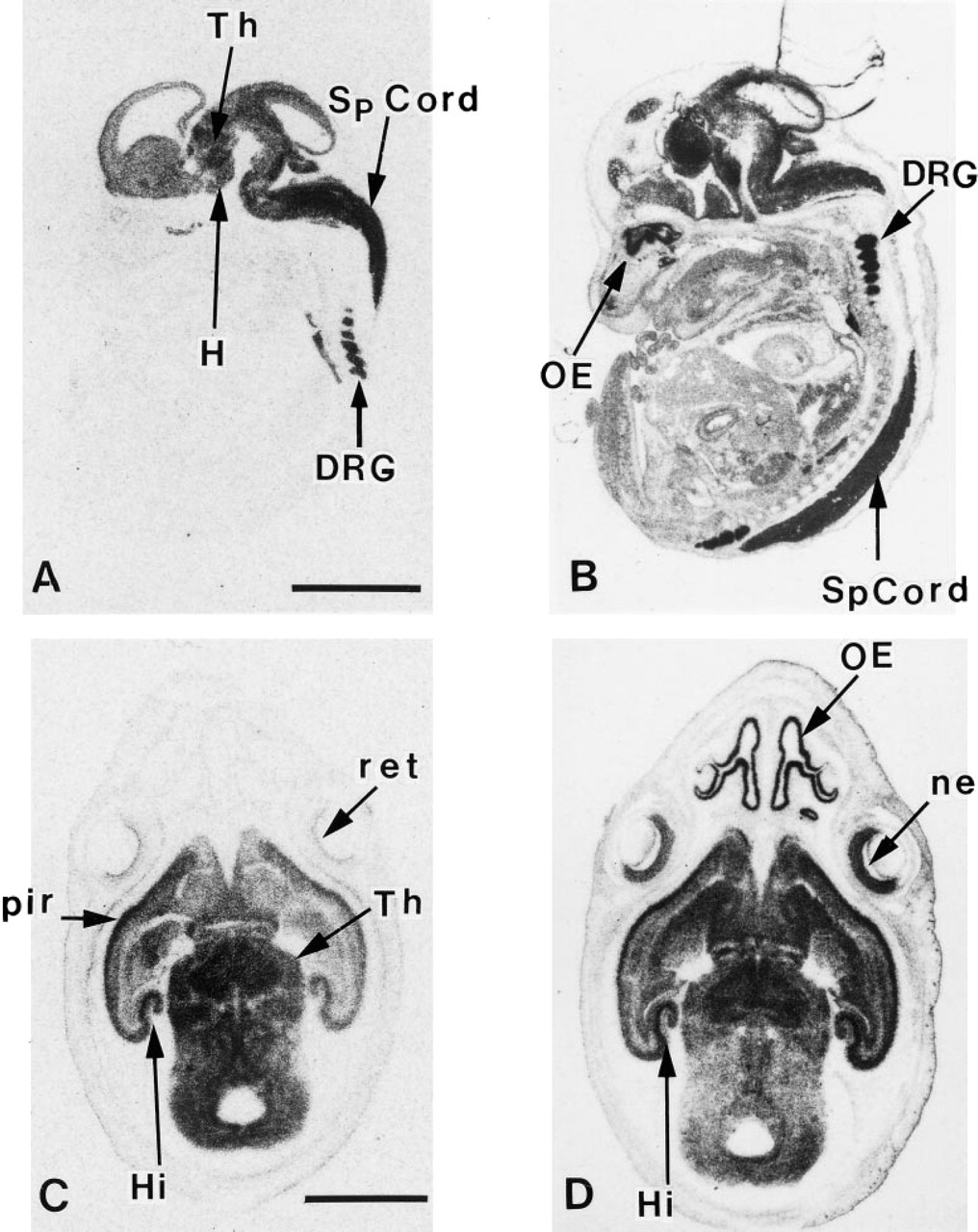
In situ hybridization detection of synapsin I and p150Glued mRNAs in developing rat embryo using 35S-labeled Dde I-digested cDNA probes. Typical localization of synapsin I
In the present set of experiments we have used in situ hybridization to assess the ability of Dde I-digested cDNA probes to detect the expression of c-fos and NT/N mRNA in the brain after acute administration of the antipsychotic drug, haloperidol. Figure 3 depicts the effects of acute haloperidol treatment on c-fos mRNA expression in the dorsolateral striatum (DLS). In accord with all previous analyses, the temporal and spatial expression of c-fos mRNAs followed the same pattern as that previously described (Merchant and Dorsa 1993), i.e., c-fos mRNA expression was observed at high levels in the DLS at 30 min after haloperidol treatment (Figure 3A). Competition control experiments were devoid of hybridization in the DLS (Figure 3B).
Figure 4 depicts a time-course analysis of the effects of acute haloperidol treatment on NT/N mRNA expression in the DLS. Similar to that seen for c-fos, the temporal and spatial expression of NT/N mRNA generally followed the same pattern as that previously described (Merchant and Dorsa 1993). Immediately after haloperidol treatment, no detectable NT/N gene expression was observed in the DLS (Figure 4A). However, 30 min after acute haloperidol administration, NT/N mRNA was detectable at moderate amounts in the DLS. By 3 hr after haloperidol treatment, NT/N mRNA expression was increasing in the DLS, as NT/N mRNA expression has been shown to peak in this region of the brain 7 hr after drug administration (Merchant and Dorsa 1993).
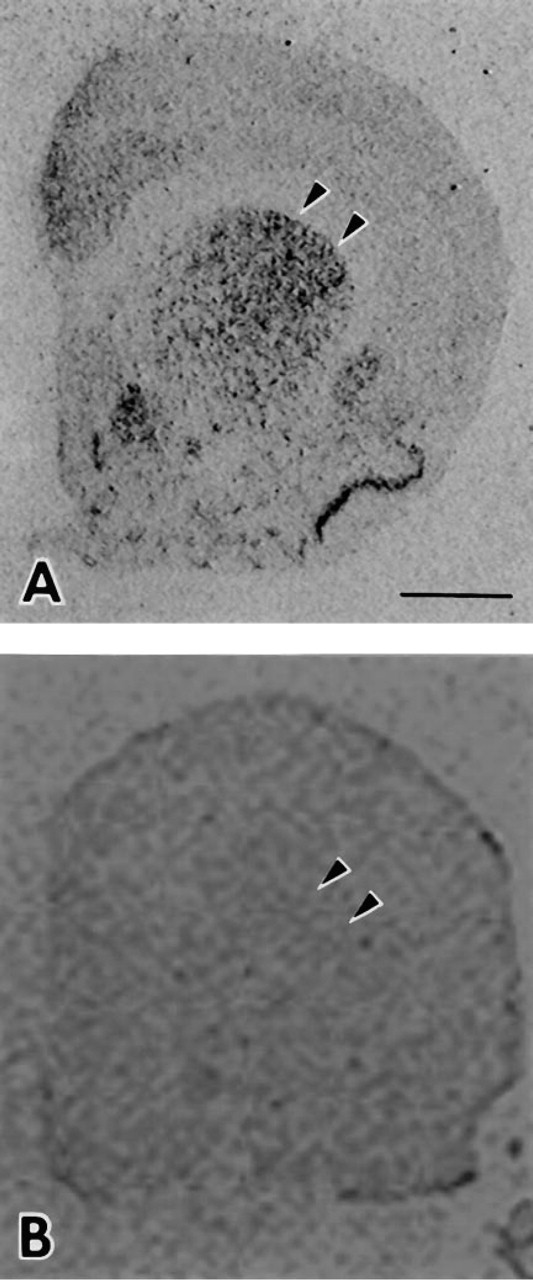
In situ hybridization detection of c-fos mRNA in rat brain at 30 min after haloperidol treatment, using 33P-labeled, Dde I-digested c-fos cDNA probes.
Non-isotopic Detection of c-jun mRNA in Adult Rat Hippocampus Using Dde I-digested cDNA Probes
To assess the ability of Dde I-digested cDNA probes to detect mRNAs in brain by non-isotopic methods, Dde-I-digested c-jun cDNA was labeled with the non-isotopic chromogen 11-digoxygenin dCTP and was used for in situ hybridization. Figure 5 presents low- and high-magnification photomicrographs of the basal expression of c-jun mRNA in the hippocampus of rat brain. In accord with previous data, hybridization of Dde I-digested c-jun cDNA to c-jun mRNA was observed in neurons located in hippocampal subfield CA3 in the hippocampus proper and in the dentate gyrus (Mellstrom et al. 1991). In addition, hybridization to c-jun mRNA in large polymorphic hilar neurons of the CA4 subfield of the hippocampus proper was markedly higher than that previously reported (Mellstrom et al. 1991).
Discussion
We present here a novel procedure for the detection of low to moderate and high-abundance mRNAs by in situ hybridization histochemistry, with particular emphasis on the study of gene expression within the CNS. This procedure employs the use of a series of small double-stranded cDNA fragments as molecular probes obtained via digestion of the cDNA clone of interest with the high-frequency recognition site restriction endonuclease Dde I. The resulting digestion products comprise a series of cDNA fragments (typically between 50 and 1000 BP) specific for a particular target mRNA sequence.
The specificity of the hybridization obtained with our Dde-I-digested cDNA probes is revealed in several ways. First, the distinct patterns of mRNA expression seen using the different cDNAs as probes testifies to the specificity for Dde-I-generated cDNA fragments to specific mRNA targets in situ. Second, control hybridization using homologous or heterologous unlabeled cDNA probes to compete with labeled probes for specific hybridization targets in situ demonstrate the disappearance of specific staining only when the homologous unlabeled cDNA is added. Together, these data demonstrate that the labeling of Dde-I-digested cDNA restriction fragments to high specific activity by random primed oligonucleotide labeling effectively produces a “family” of small cDNA probes well suited for hybridization to their specific target mRNAs in situ. The efficacy of preparing probes by Dde-I digestion also implies that the use of other high-frequency recognition site restriction enzymes, such as Hae III, Rsa I, and others, could be utilized to generate probes from cDNAs, particularly when the cDNA clone of interest has few or no Dde I recognition sites. When these enzymes are used, the target size for cDNA digestion products for use in this procedure should be 100–650 BP. Fragments of smaller size may possess significant homologies to cDNAs encoding other proteins/peptides and lead to increased background as a result of nonspecific hybridization.
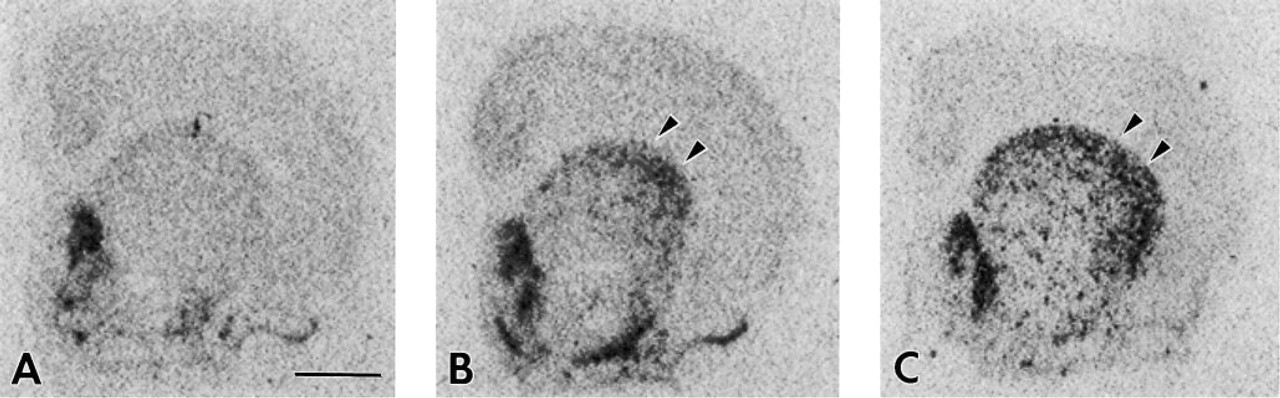
In situ hybridization detection of neurotensin/neuromedian N mRNA in rat brain at 0 min
This procedure is sensitive, as evidenced by the fact that Dde-I-digested cDNA probes detect target mRNAs at time points and locations not previously reported using other methods. For example, previous studies using cRNA probes prepared from synapsin I cDNA clone pSyn 5 (Kilimann and DeGennaro 1985) used to examine synapsin I mRNA in brain detected synapsin I transcripts only at its peak level of expression in the developing cerebellum (Haas and DeGennaro 1988). However, subsequent studies using Dde I-digested pSyn 5 cDNAs proved successful in detecting the expression of synapsin I mRNA in neurons of the CNS and PNS during development and into the adult period (Figures 2A and 2C; Melloni and DeGennaro 1994; Melloni et al. 1993, 1994). Similarly, preliminary studies using cRNA probes to detect p150Glued mRNA in the CNS were unsuccessful (Melloni and Holzbaur, unpublished observations). However, Dde I-digested p150Glued cDNA proved suitable for the detection of this mRNA (Figures 2B and 2D; Melloni et al. 1995). Further, Dde-I-digested c-jun cDNA probes detected c-jun mRNA in moderate to high levels in the dentate gyrus and at lower although detectable levels in the CA4 region of the hippocampus, where previously no such hybridization was reported (Mellstrom et al. 1991). One possible explanation for these results is that because Dde-I-digested cDNAs comprise a “family” of cDNA probes, there may be more labeled nucleotide species available for hybridization to available target sequences within one mRNA molecule. This may be especially true because comparable cRNA probes typically used are composed of a 100–400-BP linear strand of radiolabeled RNA by design (Singer and Lawrence 1986). In contrast, Dde-I cDNAs are composed of fragments along almost the entire length of the cDNA. This may effectively increase the number of hybridizing nucleic acid species within a given concentration of probe.
The general applicability of the cDNA Dde I fragmentation procedure for the study of gene expression in situ within tissues other than brain has been tested. For example, mRNA encoding p150Glued, a component of the dynactin complex implicated in the general cell processes of mitosis and nuclear migration, is detected at high levels in the olfactory epithelium and at lower yet clearly more detectable levels in all tissues of the developing rat embryo (see Figure 2). A more detailed analysis of the suitability of this procedure for the study of gene expression in tissues other than brain is now underway in the laboratory.
As expected with the use of small fragments as molecular probes for in situ hybridization, there are several caveats and concerns to consider when this procedure is used. First, because the hybridizing species are double-stranded DNA fragments, a sense nucleotide strand exists in the hybridization reaction, effectively competing with the endogenous mRNA for the anti-sense DNA probe strand. Therefore, higher concentrations of probe may be required to sufficiently detect the mRNA present in tissue sections. Although the possibility exists that this may lead to increased nonspecific background hybridization signal, we have not observed such results in our laboratory. We attribute this primarily to the physical nature of DNA: RNA interactions, which have a higher melting temperature relative to that of DNA:DNA hybrids. Hence, under the high-stringency conditions employed in this protocol, the presence of anti-sense cDNA strands probably results in negligible diminution of the effective probe concentration. Second, by nature of their design, Dde-I-digested cDNAs may not be suitable for use as molecular probes when the cDNA clone utilized possesses significant homology (>75%) to other cloned cDNAs. For example, Dde-I-digested cDNAs may not work well in detecting mRNAs encoding specific seven-transmembrane (TM) receptor mRNAs, as significant homologies exist in regions encoding TM and second messenger-linked domains. However, if necessary, small regions of the cDNA clone of interest can be isolated, digested with Dde-I, and used for in situ hybridization by this method. We caution that such an approach should include a complete review of the fragment sizes and sequence homologies generated by Dde I digestion.
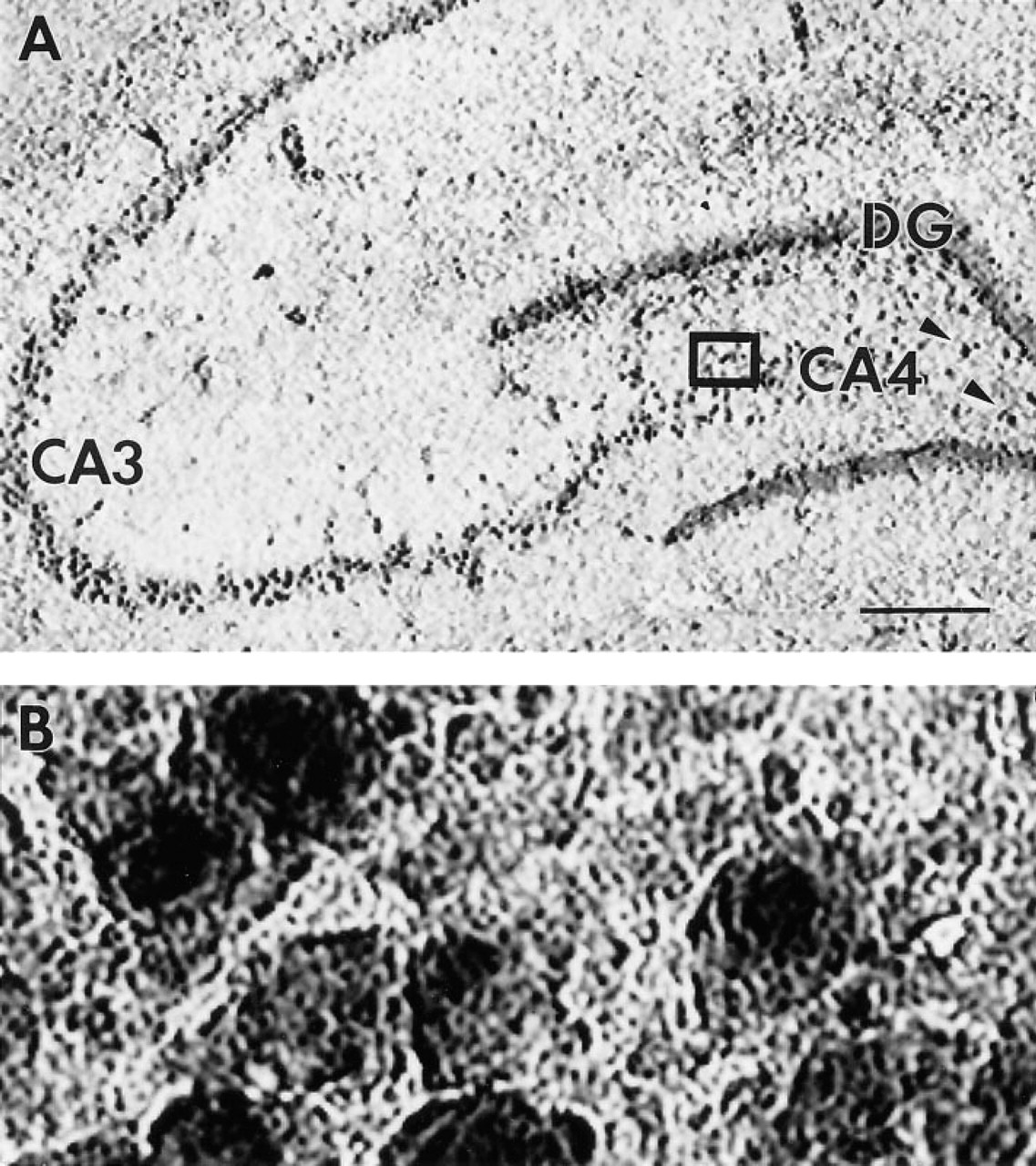
In situ hybridization detection of c-jun mRNA in the rat hippocampus using 32P-labeled, Dde I-digested cDNA probes.
In situ hybridization has proved to be an invaluable histochemical tool for study of gene expression in neurobiology. Our objective was to develop a straightforward and reliable procedure for direct analysis of mRNA in situ in frozen mammalian brain. The principal advantages of Dde-I cDNA fragmentation procedure of in situ hybridization over conventional methods are several. First, the procedure is straightforward and rapid, circumventing the tedious and often difficult nature of the preparation and use of conventional cRNA methods and minimizing time constraints inherent in the design and/or purchase of oligonucleotides. Moreover, because the Dde-I fragmentation procedure is rapid and simple, it is especially useful for the simultaneous study of multiple genes. Second, the procedure is a reproducible method of obtaining sensitive probes for in situ hybridization studies and is generally applicable to analysis of the expression of a large number of genes. Third, the procedure is sensitive and results in excellent detection of high- and moderate to low-abundance mRNA. Finally, because this procedure does not require complex recombinant DNA manipulations or the design and/or purchase of custom-synthesized oligonucleotides, it should prove useful to the non-molecular biologist studying the regulation of gene expression in nervous system tissue.
Acknowledgments
We thank Dr Erika Holzbaur for the p150Glued probe, and Paul R. Dobner for the neurotensin/neuromedian N cDNA clone and partial support of RJH. In addition, we would like to acknowledge Marcel Roy for expert technical assistance. RHM Jr. would like to extend special thanks to Dr Kimberly A. Melloni for her support and motivation.