
Abstract
The complex interplay between cells and extracellular matrix (ECM) proteins is critical for lung development. Integrins are key modulators of this interaction. The integrin subunit α8 associates with the β1-subunit to form an RGD-binding integrin. We previously showed that, in adult lung, α8 is expressed in contractile interstitial cells and smooth muscle cells and is upregulated in lung injury. To gain insight into the function of α8 during lung development, we examined the spatiotemporal expression of α8 throughout murine lung development. We compared the distribution of α8 with α-smooth muscle actin (αSMA), fibronectin (α8 ligand), and cytokeratin. α8 co-localized with αSMA and fibronectin in the peribronchial and perivascular regions. In all stages, α8 immunoreactivity was detected diffusely in the mesenchyme except for cells surrounding distal, newly forming airways. α8, αSMA, and fibronectin co-localized at tips of secondary septae in the alveolar stage. We conclude that α8 is marker for lung mesenchymal cells starting early in development. α8 is also a marker for smooth muscle cells, expressed as early as αSMA. Co-localization of α8 with fibronectin suggests a role in branching morphogenesis. Furthermore, α8 may participate in secondary septation by modulating signals from the extracellular matrix to alveolar myofibroblasts.
T
The integrin subunit α8 associates with the β1-sub-unit to form an arginine–glycine–aspartic acid (RGD)-binding integrin. Its ligands include fibronectin, vitronectin, tenascin-C, osteopontin, and nephronectin (Muller et al. 1995; Schnapp et al. 1995b; Varnum et al. 1995; Denda et al. 1998; Miner 2001). In adult tissues, α8 is expressed in cells with contractile properties including vascular and visceral smooth muscle cells, mesangial cells, and lung interstitial cells (Schnapp et al. 1995a). We previously showed that, in the adult lung, α8 is expressed in contractile interstitial cells, including alveolar myofibroblasts, lipid-containing fibroblasts, and pericytes (Levine et al. 2000).
α8 expression is increased after injury in models of pulmonary fibrosis, hepatic fibrosis, and glomerulonephritis (Hartner et al. 1999; Levine et al. 2000). The increased expression corresponds to proliferation of alveolar myofibroblasts, hepatic stellate cells, and mesangial cells, respectively. A hallmark of activation of these cells is the de novo expression of αSMA. These events are followed by remodeling of injured tissues and organ fibrosis. Because the molecular pathways of wound healing and development may be similar, we hypothesized that α8 plays an important role in development. The expression of α8 in lung development is unknown.
Our aim was to examine the spatiotemporal distribution of α8 to gain insight into its role in lung development. Because we had previously found co-expression of αSMA with α8 in activated alveolar myofibroblasts in a model of lung injury, we examined the relationship between α8 and αSMA expression in lung development. We also compared the distribution of α8 to one of its ligands, fibronectin, believed to play a role in murine lung development.
Materials and Methods
Tissue Specimens
This study was approved by the University of Washington Institutional Animal Care and Use Committee. C57BL/6 female mice were mated with C57BL/6 males and checked every morning for a vaginal plug to determine day 0.5 of gestation. Pregnant females at 12.5, 13.5, 14.5, 15.5, 16.5, and 17.5 days of gestation (E12.5–E17.5) were sacrificed and embryos were extracted via laparotomy. Gestational age was further confirmed by examination of external features. C57BL/6 pups at postnatal days 2 and 8 (PN2 and PN8) were also examined. Lungs were dissected and immersed at 4C in serial concentrations of sucrose in PBS: 5%, 10%, 15%, and 20%. After sucrose preservation, tissues were placed in OCT embedding medium (Tissue-Tek; Sakura Finetek, Torrance, CA), frozen in methylbutane that had been chilled in liquid nitrogen, and stored at −80C. Five-μm cryostat sections of tissues were obtained and placed on slides. In preparation for immunohistochemistry (IHC), sections were thawed, fixed for 3 min in acetone at −20C, allowed to air-dry at room temperature (RT) for 30 min, and rehydrated in PBS.
IHC for Light Microscopy
Slides from each time point were blocked sequentially with Peroxo-block (Zymed; San Francisco, CA) for 30 sec, 3% normal goat serum (Vector; Burlingame, CA) in PBS for 1 hr at RT, and Endogenous Avidin and Biotin Blocking Reagents (Zymed) for 10 min each. Generation of rabbit anti-α8 peptide polyclonal antiserum was previously described (Schnapp et al. 1995a). Sections were incubated overnight at 4C with α8 antibody (polyclonal antiserum 1:16,000–1:32,000 dilution) followed by 1-hr RT incubation with biotinylated goat anti-rabbit IgG antibody (1.5 μg/ml) (Vector). Immunoperoxidase complexes were formed using a Vectastain Elite ABC Kit (Vector). Color development was performed with DAB (Sigma Chemical; St Louis, MO) in the dark for 10 min, followed by DAB enhancer (Zymed) for 1 min. Sections were counterstained with hematoxylin (Vector), serially dehydrated, and mounted with Permount (Sigma). Control sections were incubated with pre-immune rabbit serum (Vector) in place of rabbit anti-α8 polyclonal antiserum or incubated with α8 antiserum preadsorbed with peptide used to generate the antiserum as previously described (Schnapp et al. 1995a).
IHC for αSMA was performed using the Mouse on Mouse (MOM) Peroxidase kit (Vector). The anti-αSMA monoclonal antibody (clone 1A4; Sigma) concentration used was 0.64 μg/ml. Color development was performed with DAB as described above. Control sections for αSMA were performed with the omission of the primary antibody.
IHC for fibronectin was performed as described for α8 using anti-fibronectin antibody (F3648; Sigma), concentration 0.044 μg/ml. Control sections were performed with the omission of the primary antibody.
Photographs were taken through a Nikon Labophot microscope (Tokyo, Japan) using ×20 and ×40 lenses with a Spot Insight Color digital camera (model 3.2.0; Diagnostic Instruments, Sterling Heights, MI).
Immunofluorescence
Immunofluorescence was performed to examine co-localization between α8 and αSMA. Sections were blocked for endogenous peroxidase for 30 sec with Peroxoblock (Zymed), then incubated with Cy3-conjugated αSMA (clone 1A4; Sigma) (5.5 μg/ml in 3% normal goat serum) for 1 hr at RT. This was followed by avidin and biotin blocking reagents (Zymed) for 10 min each. Sections were incubated overnight at 4C with α8 antiserum (1:300,000 dilution) followed by 1-hr RT incubation with biotinylated goat anti-rabbit IgG antibody (0.3 μg/ml; Vector). The signal was amplified using the TSA Biotin System (Perkin–Elmer Life Sciences; Boston, MA) by incubating the slides with streptavidin-conjugated horseradish peroxidase in PBS (1:100) for 1 hr, followed by TSA–biotin (1:50) in TSA Amplification Diluent for 4 min. Sections were then incubated for 1 hr at RT with streptavidin-conjugated Alexa 488 (Molecular Probes; Eugene, OR) at 5 μg/ml in PBS. The nuclei were counterstained with To-Pro-3 at 0.67 μg/ml (Molecular Probes) for 10 min at RT. Slides were mounted with Vectashield Hardset Medium (Vector).
To differentiate epithelial cells from mesenchymal cells, double immunofluorescence was performed with pan-cytokeratin and α8 antibodies. Immunofluorescent staining for α8 was performed as described above. The sections were then permeabilized with 0.1% Triton X-100 in PBS (PBT) for 15 min at RT. Monoclonal anti-pan cytokeratin antibody (mixture of clones C-11, PCK-26, CY-90, KS-1A3, M20 and A53-B/A2) (Sigma) was dialyzed using the Slide-A-Lyzer Mini-Dialysis Unit (Pierce; Rockford, IL) to remove phenol red to avoid quenching of the Alexa dyes. The pan-cytokeratin antibody was labeled with the Zenon One Mouse IgG1 Alexa 568 Labeling Kit (Molecular Probes) by incubating 1 μl of dialyzed antibody with 7.5 μl of the Zenon Alexa 568-labeled Fab fragments and 10 μl of PBS for 5 min at RT. Excess Fab fragments were adsorbed with 7.5 μl of IgG block for 5 min. The pan-cytokeratin antibody Zenon Fab Alexa 568 complex was diluted in PBT to a final volume of 100 μl and applied to the sections for 1 hr at RT. After washing, tissues were fixed in 4% paraformaldehyde for 10 min at RT to stabilize the Zenon Fab-Fc interaction. The slides were counterstained with To-Pro-3 and mounted as described above.
Immunofluorescent photographs were obtained using a Leica DM-R upright epifluorescent microscope (Carlsbad, IL) using ×20 lenses and ×40 oil-immersion lenses with a Leica TCS/SP Confocal Scanner equipped with argon, krypton, and helium/neon lasers. Images were superimposed with the help of ImagePro plus (version 4.0; Media Cybernetics, Carlsbad, CA) and processed with Adobe Photoshop (version 6.0; San Jose, CA).
Results
In the early pseudoglandular stage, lungs are primarily composed of mesenchymal cells (m) with interspersed airways (a) surrounded by peribronchial cells (Figures 1A–1F). The proportion of epithelial to mesenchymal cells increases dramatically during branching morphogenesis, ending around E16.5 (Figures 1G–1L and 2A–2C). In the canalicular stage, the distal airways (da) can be distinguished from the mesenchyme by delineation of the basement membrane by fibronectin (arrows in 2A and 2D). Postnatally, during the saccular stage, the interstitium has thinned significantly and gas exchange occurs in saccules (s in Figures 2G–2I). True alveoli (al) are formed within saccules during the alveolar stage by secondary septation (arrowheads) (Figures 2J–2L).
Prenatal Peribronchial and Perivascular Expression
α8 immunoreactivity was present in the peribronchial and perivascular regions throughout lung development in a pattern similar to that of αSMA (Figures 1 and 2). To further characterize the localization of α8 and αSMA, we performed co-immunofluorescence and found that α8 co-localized with αSMA in the peribronchial region throughout smooth muscle cell development at all time points examined (Figure 3). Fibronectin, a ligand for α8, was prominently expressed in the basement membrane at the epithelial-mesenchymal interface throughout lung development (arrows in Figures 1A, 1D, 1G, 1J, 2A, and 2D). Fibronectin was also present in the peribronchial regions beyond the epithelial-mesenchymal interface. In the early pseudoglandular stage, the peribronchial expression of fibronectin was more limited compared to that of α8 and αSMA (Figures 1A–1F). However, from the mid-pseudoglandular stage and beyond, fibronectin staining was more similar to that of α8 and αSMA (Figures 1G–1L and 2A–2F).
Prenatal Mesenchymal Expression
Prenatally, α8 was expressed diffusely throughout the lung mesenchyme (Figures 1B, 1E, 1H, 1K, 2B, and 2E). There appeared to be some mesenchymal cells that did not express α8 but it was difficult to define the pattern of mesenchymal α8 expression using light microscopy alone. We used a pan-cytokeratin antibody to identify epithelial cells and performed co-immunofluorescence for α8 and cytokeratin. Cytokeratin staining revealed that many of the α8-negative cells visualized by light microscopy were actually clusters of small airways (Figure 4). Furthermore, we found that all mesenchymal cells were immunoreactive for α8 except for mesenchymal cells surrounding newly forming, distal airways (da) (Figures 3I and 4). Fibronectin was expressed as diffusely as α8 in the mesenchyme during the mid-pseudoglandular stage (Figures 1G and 1J) but less prominently during all other prenatal stages (Figures 1A, 1D, 2A, and 2D). In contrast to α8 and fibronectin, there is minimal mesenchymal expression of αSMA until birth (Figures 1C, 1F, 1I, 1L, 2C, and 2F).
Postnatal Expression
Postnatally, during the later saccular and alveolar stages, the mesenchymal (interstitial) distribution of α8 continued to be diffuse Figures 2H, 2K, 3G, and 3H). Fibronectin was also expressed diffusely in the interstitium during the late saccular stage (Figure 2G), then less diffusely in the alveolar stage (Figure 2J). During the late saccular stage, αSMA was expressed in alveolar myofibroblasts at sites of future secondary septum formation (arrows in Figures 2I and 3G), then at the leading tip of secondary septae in the alveolar stage (arrowheads in Figures 2L and 3H). α8 co-localized with αSMA in alveolar myofibroblasts at these sites (Figures 3G, 3H, and 3J). Fibronectin also localized to the tips of secondary septae in the alveolar stage at PN8 (Figure 2J), suggesting co-localization of fibronectin, α8, and αSMA at these sites.
Although yellow fluorescence indicated co-localization of α8 and αSMA in the peribronchial and vascular regions (Figures 3A–3F and 3I), there was no yellow fluorescence to indicate co-localization in alveolar myofibroblasts (Figures 3G and 3H). However, the intracellular location of α8 and αSMA is different (α8 is a transmembrane protein and αSMA is a cytoskeletal protein). Examination under higher magnification confirmed localization of α8 and αSMA in the same cells (Figure 3J).
α8 immunoreactivity was not present on endothelial or epithelial cells at any of the time points examined.
Discussion
We investigated the spatiotemporal expression pattern of the integrin subunit α8 in the developing murine lung and examined its co-localization with αSMA, fibronectin, and cytokeratin.
Murine lung development begins at approximately 9.5 days of gestation, when the trachea and mainstem bronchi develop from the foregut followed by lateral branching to the segmental level (embryonic stage) (Ten Have-Opbroek 1991). Further sequential division of the airways occurs by branching morphogenesis during the pseudoglandular stage (E12.0–E16.5). The blood-air barrier is formed during the canalicular stage (E16.5–E17.5) accompanied by further epithelial cell differentiation. The distal airspaces expand into alveolar saccules, with a decrease in interstitium during the saccular stage (E17.5 to PN5). Finally, true alveoli are formed by secondary septation within the saccules during the alveolar stage (5–PN30).
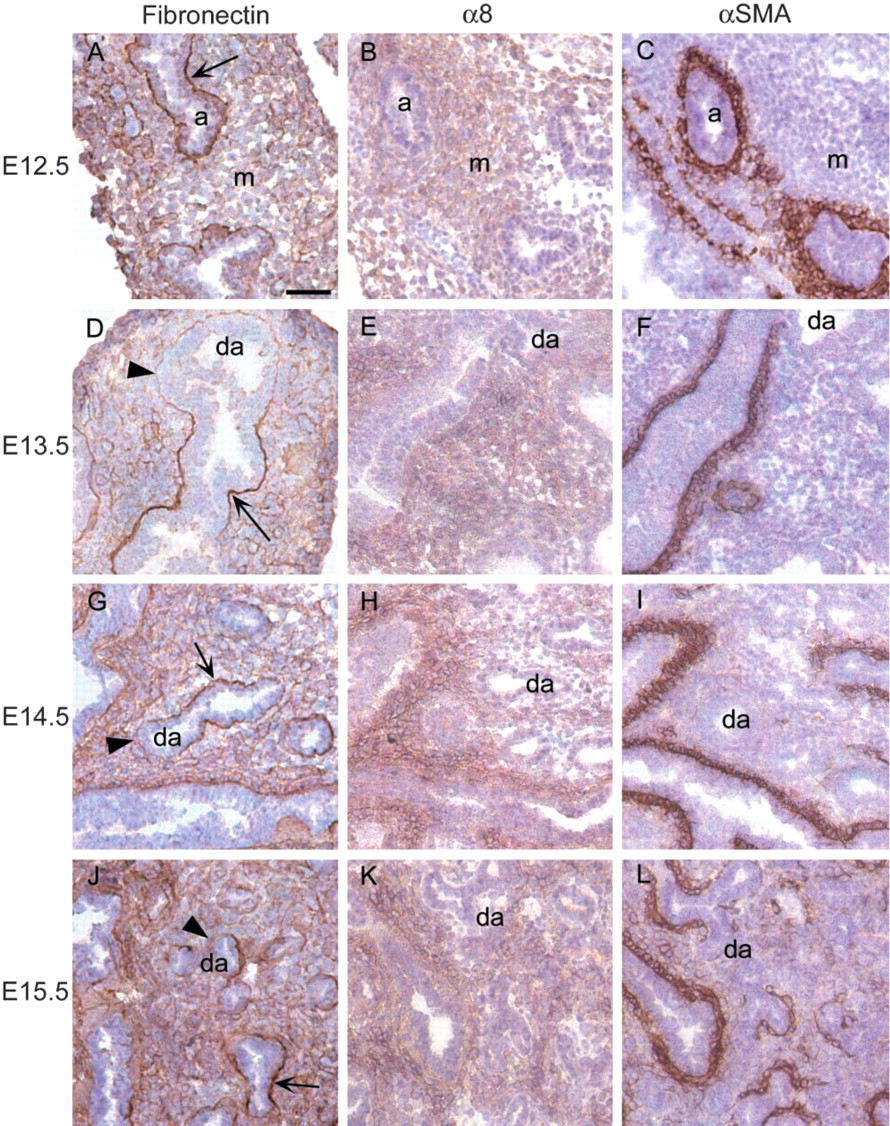
Distribution of fibronectin, α8, and αSMA in the pseudoglandular stage (E12.5–E15.5). α8 immunoreactivity is present in the mesenchyme (m) throughout the pseudoglandular stage (
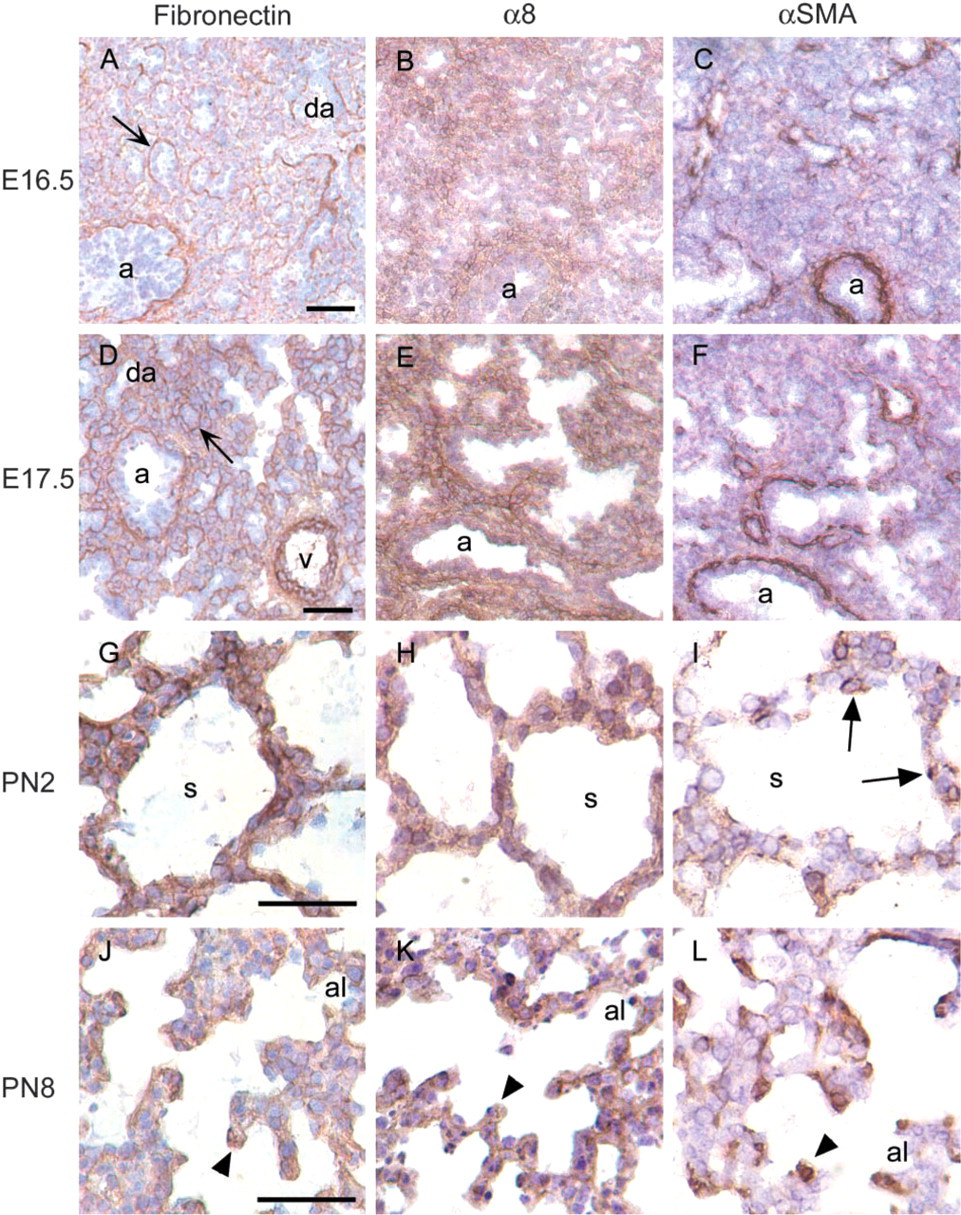
Distribution of α8 and αSMA in the late pseudoglandular (E16.5), canalicular (E16.5–E17.5), saccular (E17.5, PN2), and alveolar stages (PN8). During the late pseudoglandular, canalicular, and saccular stages, multiple small distal airways (da) invest the mesenchyme and are outlined by fibronectin in basement membrane at the epithelial-mesenchymal interface (arrows in
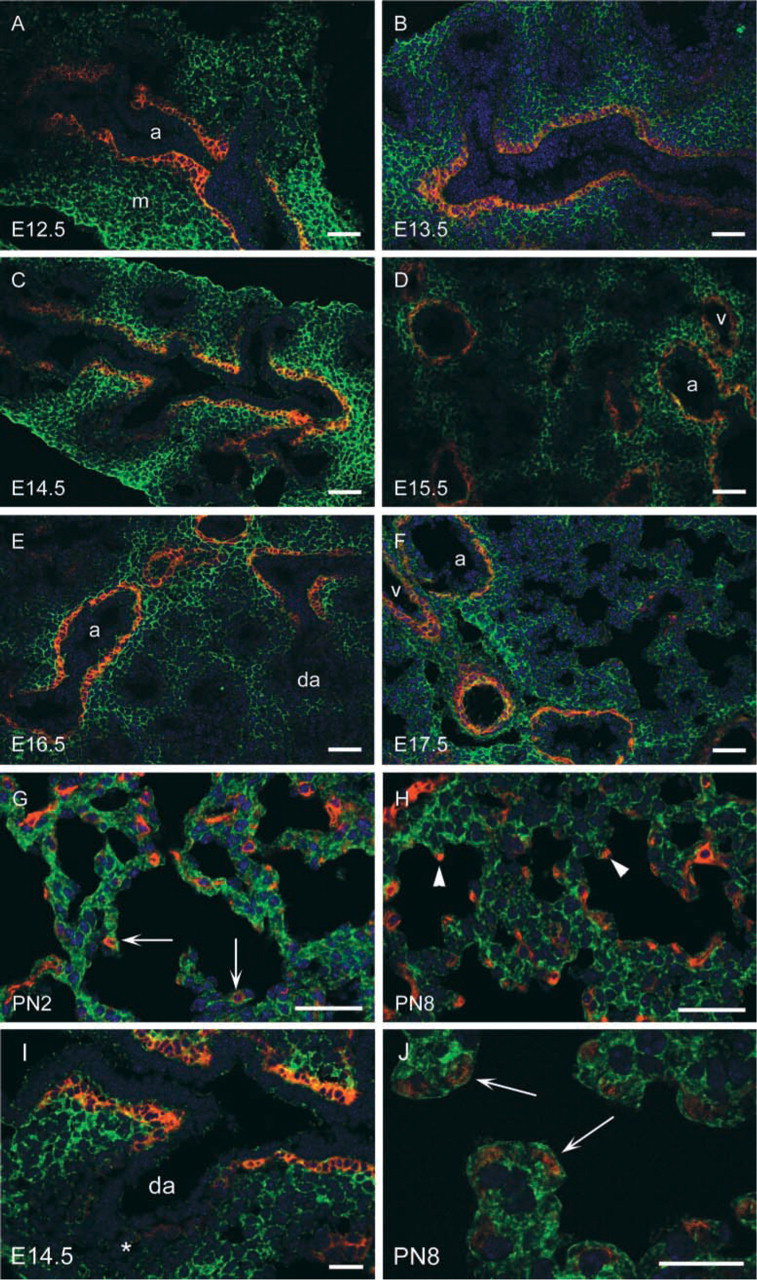
(
αSMA has been identified as one of the earliest cytoskeletal markers for smooth muscle cell differentiation (Mitchell et al. 1990). We show that α8 co-localizes with αSMA in developing smooth muscle cells as early as E12.5, demonstrating that it is also an early marker of smooth muscle cell differentiation. Fibronectin is prominently expressed in the basement membrane at the epithelial-mesenchymal interface throughout development. Starting in the mid-pseudoglandular stage, fibronectin co-localized with α8 and αSMA in peribronchial and perivascular regions. In vitro, fibronectin has been shown to affect smooth muscle cell proliferation and activation of focal adhesion kinase (FAK) (Morla and Mogford 2000). Fibronectin has also been shown to affect myoblast differentiation (Menko and Boettiger 1987). We speculate that α8 may play a role in mediating signals from the extracellular matrix to developing smooth muscle cells during lung development.
Fibronectin is thought to play an important role in branching morphogenesis in the lung (Roman and McDonald 1992; Roman 1997). Branching morphogenesis is inhibited in lung explants exposed to RGD-containing peptides that compete with ECM proteins for integrin binding (Roman et al. 1991). The basement membrane remodeling hypothesis (Bernfield 1981; Mollard and Dziadek 1998) proposes that bud outgrowth is the result of a decrease in basement membrane components, such as fibronectin, accompanied by increased epithelial cell proliferation. Fibronectin is localized most prominently in the mesenchymal-epithelial interface at sites of airway bifurcation and clefts of airway formation, and it is decreased or absent around budding airways. Interestingly, mesenchymal cells were diffusely α8-positive except for those surrounding distal newer airways. Decreased fibronectin associated with lack of α8 expression at the sites of distal airways may provide a permissive environment for airway budding. Conversely, α8-fibronectin interactions may influence cleft formation and airway bifurcation.
Similar to lung development, kidney development occurs by branching morphogenesis. In an α8 knockout model, homozygotic mice displayed renal agenesis or dysgenesis and most died within the first few days of birth (Muller et al. 1997). Interestingly, branching morphogenesis of the ureteric bud into the renal mesenchyme was abnormal, suggesting that α8 plays a critical role in the mesenchymal-epithelial interaction in kidney development. The initial report showed grossly normal lungs (Muller et al. 1997). However, subtle abnormalities in branching morphogenesis of the lung have been recently described (Schittny et al. 2002). In 60% of the α8-null lung explants, branching was reduced by 33–55% and the pattern was irregular. Redundancy between the function of α8 and that of other integrins, such as α5, may account for more subtle abnormalities in lung development in surviving mice. Our α8 expression data also support a role for α8 in branching morphogenesis.
By the late saccular stage and the alveolar stage, the distribution of α8 was diffuse throughout the interstitium (Figures 3G and 3H), similar to the pattern seen in adult tissues (Schnapp et al. 1995a). We previously showed that the widespread distribution of α8 in mature alveolar walls localizes to contractile interstitial cells (Levine et al. 2000). α8-positive mesenchymal cells during development probably represent precursors to contractile interstitial cells. Furthermore, α8 co-localized with αSMA in alveolar myofibroblasts during secondary septation. Co-localization in these cells has also been described in alveolar myofibroblasts, which are present in the adult lung in response to injury (Levine et al. 2000). Interestingly, fibronectin also localized to the tips of the secondary septae where α8- and αSMA-positive alveolar myofibroblasts are located. Therefore, α8 is in a position to mediate signals from the extracellular matrix (fibronectin) to alveolar myofibroblasts and hence to contribute to secondary septation.
In summary, integrin subunit α8 is expressed in the mesenchyme throughout lung development and co-localizes in the peribronchial and perivascular regions with αSMA. Along with αSMA, α8 is an early marker for smooth muscle cells. α8 may interact with fibronectin in the peribronchial regions and play a role in smooth muscle cell differentiation. α8 is also an early marker for lung mesenchymal cells. Decreased fibronectin expression in distal budding airways is associated with α8 negativity of mesenchymal cells in the same regions. Considering the data from the α8 knockout model (Schittny et al. 2002), we speculate that expression of α8 may play a role in branching morphogenesis by interacting with ECM components, such as fibronectin, to coordinate airway growth, cleft formation, and airway bifurcation. Furthermore, α8 may mediate signals from the ECM to alveolar myofibroblasts and participate in secondary septation.
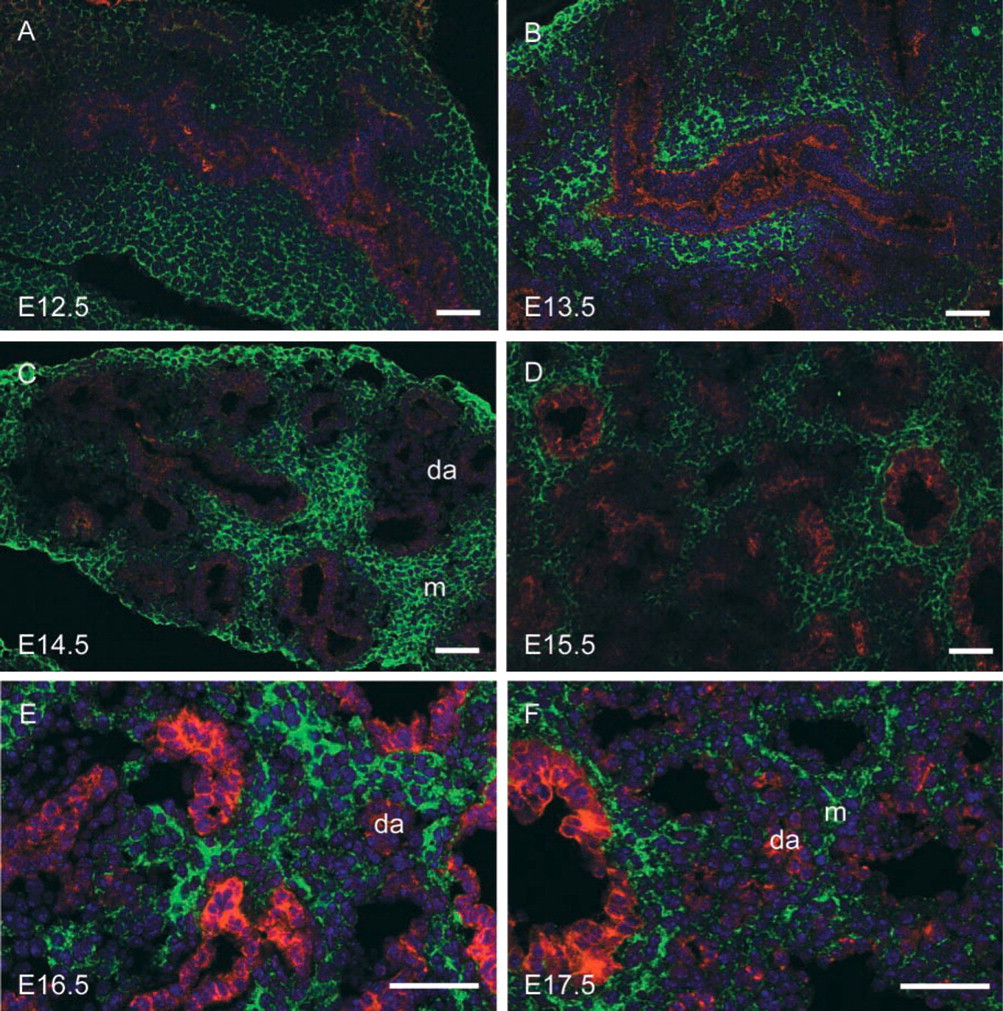
Immunofluorescence of α8 and cytokeratin (E14.5–E17.5). The mesenchymal distribution of α8 (green) in the late pseudoglandular and early saccular stages is demonstrated by using immunofluorescent labeling of pan-cytokeratin (red) to differentiate epithelial cells of distal airways (da) from mesenchymal/interstitial cells (m). The nuclear counterstain is blue. α8 is expressed in the mesenchyme-interstitium throughout development, except for mesenchymal cells surrounding the more distal and newly formed airways (da, distal airways). Bars = 40 μm.
Footnotes
Acknowledgements
Supported by an AHA grant-in-aid, Northwest Affiliate (LMS).
We thank Jeremy Ehly for technical support on the confocal microscope.