
Abstract
Inhalation of ozone by Rhesus monkeys results in epithelial injury and granulocyte influx in both conducting airways and respiratory bronchioles. We have reported that ozone-induced neutrophil recruitment and subsequent epithelial repair can be inhibited in vivo with a CD18 antibody. The antibody-mediated effect is abrogated by local instillation of C5a (a CD18-independent neutrophil chemoattractant), thereby demonstrating a role for neutrophils in lung epithelial repair processes. As an extension of this study, we examined the effect of ozone and neutrophil influx on epithelial expression of the β6 integrin, an adhesion molecule associated with proliferation and repair. Expression of β6 integrin was determined by immunohistochemistry for ozone-exposed monkeys treated with either control immunoglobulins or a CD18 antibody. The tracheal epithelium of ozone-exposed monkeys treated with control immunglobulins expressed the β6 integrin. In contrast, the tracheal epithelium of ozone-exposed monkeys treated with CD18 antibody exhibited very low to undetectable expression of β6 integrin. In association with C5a instillation and neutrophil influx, β6 integrin was also observed in respiratory bronchiolar epithelium from both control and ozone-exposed animals. These findings cumulatively suggest that lung epithelial cell expression of β6 integrin is associated with sites of neutrophil recruitment.
Keywords
E
To further study the role of neutrophils in repair of ozone-injured epithelial cells, Rhesus monkey airway epithelium was evaluated for expression of β6 integrin. The β6 integrin subunit forms a heterodimer with the αv integrin subunit. This adhesion molecule appears to be unique to epithelia and can mediate attachment to matrix proteins (fibronectin, tenascin, vitronectin), as well as TGFβ1 (Sheppard et al. 1990; Busk et al. 1992; Prieto et al. 1993; Huang et al. 1998; Munger et al. 1999). In normal adult human and monkey airways, β6 integrin protein and mRNA are undetectable (Breuss et al. 1993, 1995; Weinacker et al. 1995). However, the β6 integrin is detectable during fetal lung development and in focal regions of inflammation within the airways, suggesting an important role for epithelial remodeling and repair. Furthermore, it has been demonstrated that transfection of cDNA specific for β6 integrin into a human colon carcinoma cell line enhances proliferation (Agrez et al. 1994). To determine how β6 integrin plays a role in ozone-induced epithelial cell injury and repair processes, this study investigated the expression of this integrin sub-unit in monkeys exposed to an acute dose of ozone accompanied by neutrophil influx into the airways.
Materials and Methods
Antibodies
Mouse anti-human CD18 monoclonal antibody R15.7 (IgG1) was a generous gift from Dr. Robert Rothelin (Boehringer Ingelheim Pharmaceuticals; Ingelheim, Germany). IgG1 isotype control immunoglobulins were purified from the mouse plasmacytoma cell line MOPC-21 (ATCC; Rockville, MD). Mouse anti-human β6 integrin monoclonal antibody Csbeta6 (IgG1) was generated by immunizing β6 subunit knockout mice with secreted αvβ6 heterodimer as described (Huang et al. 1998).
Animals/Ozone Exposure
Young male Rhesus monkeys weighing 5.1–7.6 kg and aged from 3 years and 8 months to 3 years and 10 months were randomly assigned to either ozone or control (filtered air) experimental groups as follows: (a) filtered air + CD18 antibody (n = 2); (b) filtered air + isotype control (n = 2); (c) ozone + CD18 antibody (n = 6); and (d) ozone + isotype control (n = 6). All monkeys were given a comprehensive physical examination, including a chest radiograph and complete blood count, before exposures. Before initiation of experiments, monkeys were transferred to exposure chambers for a 1-week acclimatization in filtered air.
Eight hours before ozone/filtered air exposures, monkeys were given either anti-CD18 or isotype control immunoglobulins via the cephalic vein at 1 mg/kg body weight. It has previously been determined by flow cytometry that this concentration of CD18 antibody is saturating for circulating and lavage neutrophils for the duration of a 12-hr ozone exposure/postexposure period (Hyde et al. 1999). Monkeys were exposed individually in chambers of 4.2-m3 capacity that were ventilated at a rate of 30 changes per hour with filtered air at 24 ± 2C and 40–50% relative humidity. Ozone concentrations were monitored using an ultraviolet ozone analyzer (Dasibi Environmental; Glendale, CA). Monkeys were exposed to either 0 ppm (filtered air) or 0.8 ppm ozone for 8 hr. Immediately at the end of the 8-hr exposure period, all monkeys were anesthetized with ketamine and recombinant C5a (Sigma; St Louis, MO) was instilled into the right middle and right caudal lobes. At 4 hr after exposure in filtered air, monkeys were necropsied. All aspects of animal work were performed in accordance with the institutional guidelines for the California Regional Primate Research Center. The right accessory lobe (branch adjacent to the right caudal lobe) and trachea from each monkey were embedded in OCT freezing compound (Sakura Finetek; Torrance, CA) and frozen in isopentane/liquid nitrogen.
Immunohistochemistry
For immunoperoxidase staining, 5-μm cryosections of the accessory lobe and trachea were fixed in ice-cold acetone for 10 min and allowed to dry. To reduce endogenous peroxidase, cryosections were treated with 3% H2O2 for 5 min at room temperature (RT). Neutrophils in cryosections could not be completely blocked for endogenous peroxidase without destroying the integrity of the tissue. To block nonspecific binding of antibodies, cryosections were incubated overnight in goat serum at 4C before immunostaining. All monkeys in this study were treated with purified mouse IgG1 (either MOPC-21 or R15.7) as a part of the experimental/exposure design. Therefore, all tissue sections contained saturating quantities of mouse immunoglobulins as a control for immunohistochemical staining. For detection of β6 integrin, cryosections were incubated at RT with mouse anti-human β6 integrin monoclonal antibody (CSbeta6) for 1 hr. After a PBS wash, cryosections were incubated with biotinylated goat anti-mouse IgG (Vector Laboratories; Burlingame, CA) for 1 hr at 1:100 dilution. Sections were then incubated with a horseradish peroxidase-avidin-biotin complex (Vector Laboratories) for 30 min and developed using 0.5% 3,3′-diaminobenzidine (DAB)/0.01% H2O2 as a chromogen. For detection of β6 integrin in monkeys previously treated with anti-CD18 antibody, CSbeta6 was directly biotinylated using a Sulfo-NHS-LC Biotinylation kit (Pierce; Rockford, IL). After a 1-hr incubation at RT with biotinylated Csbeta6, sections were incubated with the horseradish peroxidase-avidin-biotin complex for 30 min and developed using an enhancer kit for DAB (DAB-Black; Zymed Laboratories, South San Francisco, CA) as a chromogen.
Results
We used an immunohistochemical approach to evaluate protein expression of the β6 integrin subunit by conducting airway epithelium from Rhesus monkeys treated with isotype control immunoglobulins, comparing the effects of 8-hr exposure to 0.8 ppm ozone (n = 6) vs filtered air (n = 2). As expected, tracheal and bronchial epithelium of filtered air control monkeys had very low to undetectable β6 integrin expression (Figure 1A). At the level of the trachea (Figure 1C) and bronchi (not shown), epithelial cell expression for β6 integrin was observed in all ozone-exposed monkeys treated with isotype control immunoglobulins. In the trachea, immunohistochemical staining for β6 integrin was detected in both apical and basolateral regions in individual epithelial cells.
We have previously shown that ozone-induced neutrophil influx into Rhesus monkey airways is CD18-dependent, but instillation of C5a into distal airways results in CD18-independent neutrophil recruitment, most likely from the pulmonary circulation (Hyde et al. 1999). Because all animals in this study received C5a instillations in both the right middle and right caudal lobes, distal airways (alveoli and respiratory bronchioles) of filtered air control monkeys contained neutrophils. However, in contrast to tracheobronchial epithelium, β6 integrin was expressed on respiratory bronchiolar epithelium by both filtered air- and ozone-exposed monkey groups (Figure 2). In the case of filtered air-exposed monkeys, expression of β6 integrin by respiratory bronchiolar epithelial cells was limited to airways that had neutrophil accumulations, a phenomenon associated with C5a instillation. C5a-mediated neutrophil recruitment could not be distinguished from that resulting from oxidant injury in ozone-exposed monkeys.
On the basis of our findings of β6 integrin expression in respiratory bronchioles of both filtered air- and ozone-exposed monkeys, we next evaluated if the expression of β6 integrin was the direct result of neutrophil emigration into airways (as opposed to ozone exposure). Although C5a instillation in distal airways resulted in CD18-independent neutrophil emigration in respiratory bronchiolar epithelium, tracheal epithelium was not affected by this treatment. Therefore, we compared the expression of β6 integrin on tracheal epithelium from ozone-exposed monkeys treated with isotype control immunoglobulins vs ozone-exposed monkeys treated with anti-CD18 immunoglobulins (n = 6). CD18 has been shown to play a significant role in the transepithelial migration of neutrophils across large airway epithelium in vitro (Liu et al. 1996). As expected, neutrophils were rarely observed in the epithelial compartment of tracheas from CD18 antibody-treated monkeys. In contrast to the finding of β6 integrin expression exhibited by tracheal epithelium from monkeys treated with control immunoglobulins, the tracheal epithelium of all animals that were treated with CD18 antibody expressed very low to undetectable β6 integrin, comparable to filtered air controls (Figure 3).
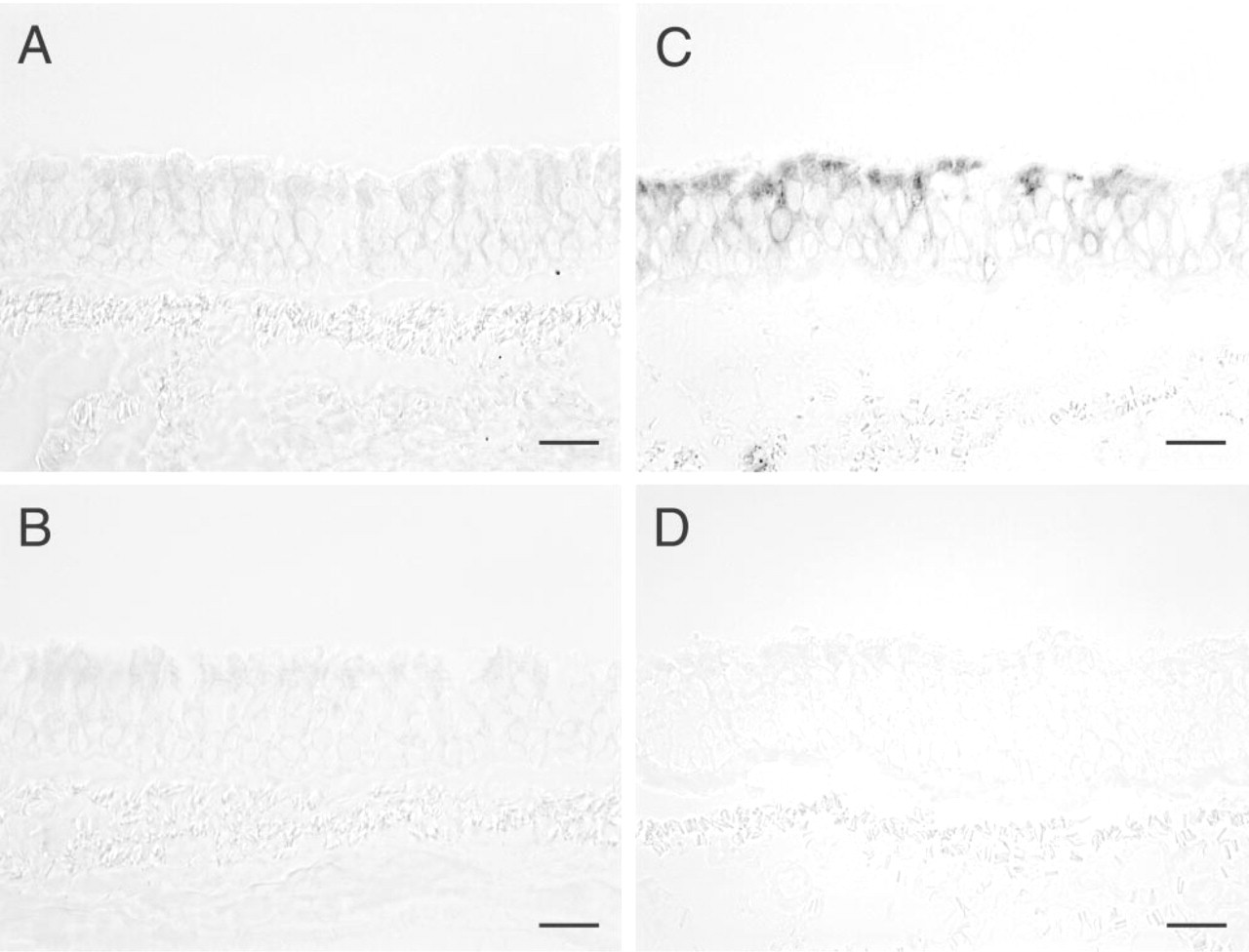
Expression of β6 integrin in Rhesus monkey trachea after exposure to filtered air or 0.8 ppm ozone. Cryosections of rhesus monkey trachea were stained by the immunoperoxidase method for expression of β6 integrin. (
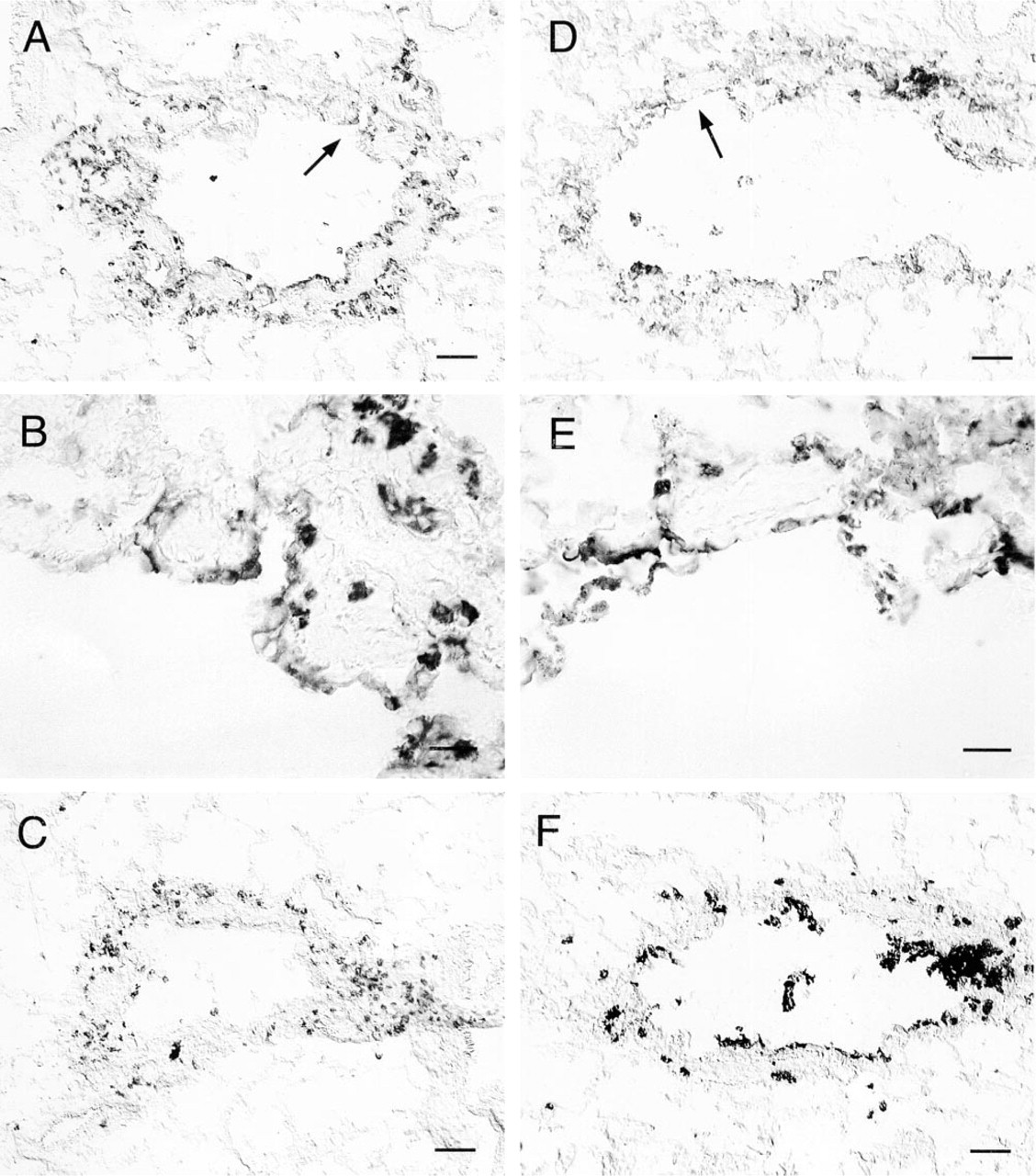
Expression of β6 integrin in Rhesus monkey respiratory bronchiolar epithelium after exposure to filtered air or 0.8 ppm ozone. Cryosections of distal airways from Rhesus monkey were stained by the immunoperoxidase method for expression of β6 integrin. (
Discussion
Depending on the inflammatory stimuli, neutrophil recruitment in the lung can occur via CD18-dependent or CD18-independent mechanisms (Hogg and Doerschuk 1995). It is likely that the requirement for β2 integrin is contingent on the vascular bed (pulmonary vs systemic) from which neutrophils extravasate. We have previously reported that inhalation of 0.8–0.96 ppm ozone for 8 hr results in a defined sequence of CD18-dependent neutrophil recruitment and epithelial cell repair in the non-human primate lung. We have further shown that instillation of C5a into distal airwaysin the lung results in localized CD18-independent neutrophil recruitment, most likely from the pulmonary circulation. In ozone-exposed monkeys treated with neutralizing CD18 immunoglobulins, neutrophil recruitment into distal airways via C5a results in repair of injured epithelium (Hyde et al. 1999). In this study, we have expanded on these prior observations and have found upregulated expression of the β6 integrin subunit by epithelial cells that line monkey trachea and bronchi after a necrotizing dose of ozone (0.8 ppm). The β6 integrin was also highly expressed by respiratory bronchiolar epithelium in filtered-air control animals in this study. Specifically, β6 integrin immunohistochemical staining was associated with airways containing C5a-induced neutrophil accumulations. This apparent independence of β6 integrin expression from ozone-induced epithelial cell injury in the context of this study suggested that upregulation of this adhesion molecule was instead a consequence of inflammatory cell trafficking in airways. Therefore, we examined the tracheal epithelium from ozone-exposed monkeys treated with CD18-neutralizing immunoglobulins, which we have previously shown to significantly inhibit ozone-induced neutrophil emigration in the most proximal airways. By immunohistochemical analysis, we found that expression of β6 integrin by tracheal epithelial cells from ozone-exposed, CD18 antibody-treated animals was comparable to that in filtered-air controls, suggesting that neutrophil movement across airway epithelium is required for upregulation of β6 integrin. In support of this notion, a previous study has demonstrated that alveolar epithelium can express enhanced levels of β6 integrin at focal sites of acute inflammation in patients with bronchiolitis obliterans or allergic alveolitis (Breuss et al. 1995). Experimental models of acute lung inflammation in rabbit and sheep have shown pronounced upregulation of mRNA for β6 integrin in alveolar epithelial cells at 5 hr after injury. In addition, β6 integrin was found to be expressed by proximal airway epithelium from smokers undergoing lung resection (Weinacker et al. 1995).
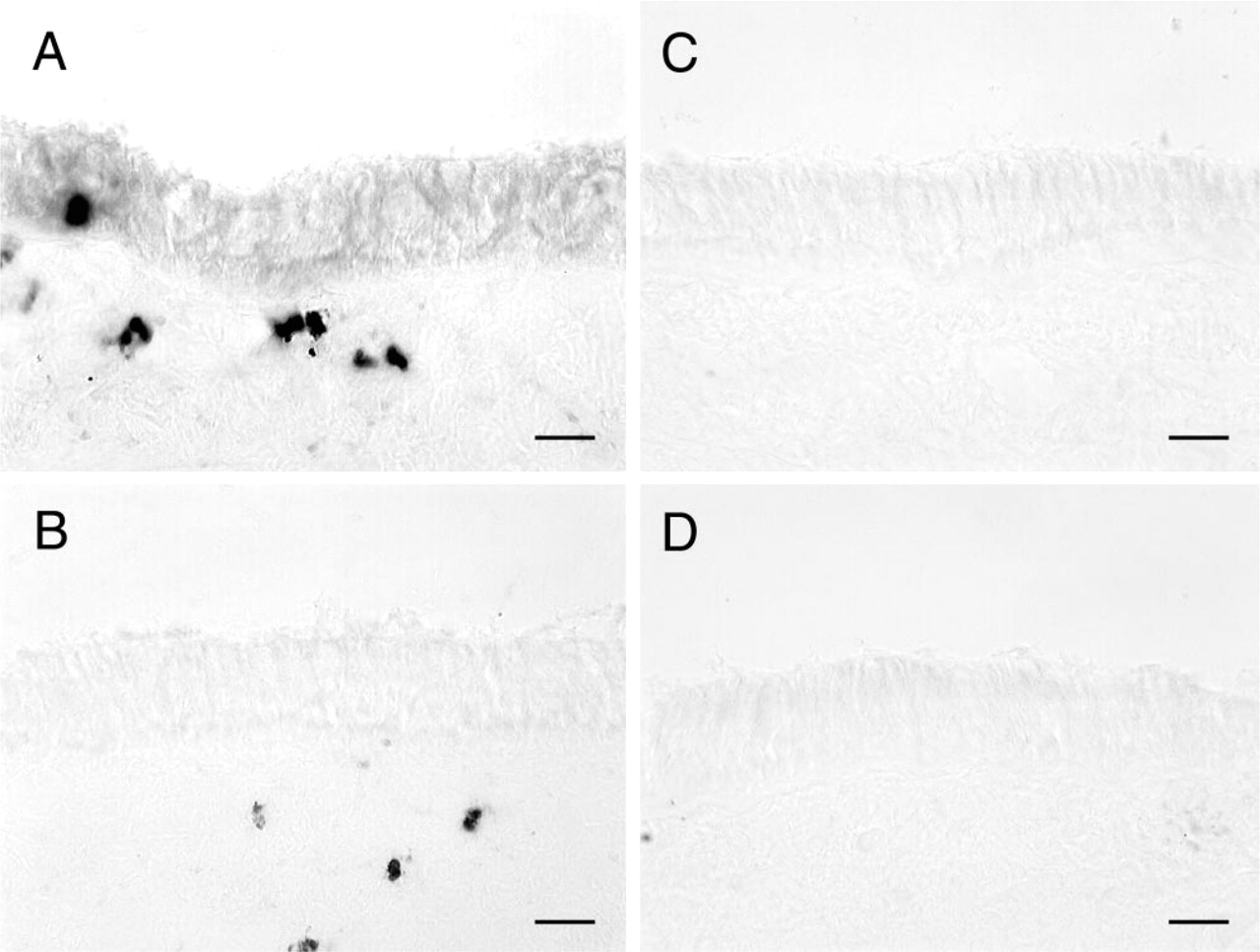
Effect of CD18-mediated inhibition of neutrophil recruitment on expression of β6 integrin in Rhesus monkey trachea after exposure to 0.8 ppm ozone. Cryosections of trachea from ozone-exposed Rhesus monkeys were stained by a nickel-enhanced immunoperoxidase method for expression of β6 integrin. (
The αvβ6 integrin complex can mediate adhesion and spreading of epithelial cells on fibronectin in vitro (Weinacker et al. 1994). Bronchoalveolar lavage samples from human subjects exposed to ambient levels of ozone contain elevated levels of fibronectin (Koren et al. 1989). In the context of lung injury, the interaction of this extracellular matrix protein with αvβ6 may play a role in the migration of airway epithelial cells during reparative processes. In an animal model of epidermal injury, Breuss and colleagues (1995) have described upregulated expression of β6 integrin in migrating keratinocytes that border a wound site in human skin grafts transplanted onto mice. However, mice that are homozygous for a null mutation of the β6 subunit gene do not exhibit abnormalities in the rate of wound healing, indicating that αvβ6 is not essential for resolution of epidermal injury (Huang et al. 1996).
The regulation of αvβ6 integrin expression appears to be linked to epithelial cell proliferation. Although immunostaining of normal human proximal airways does not indicate expression of the β6 subunit, proliferating cultures of primary tracheobronchial epithelial cells isolated from normal human subjects do express high levels of this integrin (Weinacker et al. 1995). Furthermore, several growth factors, including epidermal growth factor, transforming growth factor α, and transforming growth factor β can increase surface expression of αvβ6 on primary cultures of human airway epithelial cells (Wang et al. 1996). As yet, we do not know how the interaction of neutrophils (or other leukocytes) with epithelial cells can enhance expression of β6 integrin. Neutrophils do express granulin, a growth factor that has structural and functional parallels to the family of epidermal growth factor-related peptides; 6-kD granulin peptides are stored in intracellular vesicles (Bateman et al. 1990). Lung epithelial cells express receptors for the granulin/epithelin family of growth factors, suggesting a functional role for neutrophil-derived granulin in the repair of airway epithelium after resolution of acute inflammation (Xia and Serrero 1998).
Mice that are homozygous for a null mutation of the β6 subunit gene phenotypically exhibit pronounced lymphocyte accumulation in conducting airways, suggesting that the αvβ6 integrin complex may play a role in downregulating inflammation in the lung (Huang et al. 1996). An immunomodulatory role for this integrin complex has recently been supported by the finding that αvβ6 can bind and activate the latent form of TGFβ1 (Munger et al. 1999). In vitro, TGFβ can inhibit alveolar macrophage production of proinflammatory cytokines (IL-8, IL-1β, TNF-α) (Fadok et al. 1998). β6 subunit knockout mice are protected from development of TGFβ1-dependent, bleomycin-induced pulmonary fibrosis, yet leukocyte recruitment into the lung after treatment with bleomycin was not inhibited (Munger et al. 1999). This further implies downregulatory functions for αvβ6 during inflammatory events in the airways. Given that localized expression of β6 integrin in the Rhesus monkey appears to be associated with conditions of acute inflammation, these findings cumulatively suggest that expression of β6 integrin by lung epithelium correlates with sites of leukocyte recruitment and may be related to the resolution of inflammatory events in the airways.
Footnotes
Acknowledgments
Supported in part by Tobacco Related Disease Research Program Grants 6KT-0411 and 8IT-0054 (to LAM) and NIEHS ES-00628 (to DMH).