
Abstract
The uptake of monoamines into the secretory granules of monoamine-storing neuroendocrine cells is mediated by vesicular monoamine transporter protein 1 or 2 (VMAT1 or VMAT2). This study analyzed the expression of VMAT1 and VMAT2 in endocrine cells of normal human and monkey pancreas. The expression of VMAT1 and VMAT2 was also examined in infants with hyperinsulinemic hypoglycemia and in adults with pancreatic endocrine tumors (PETs). Using immunohistochemistry (IHC) and in situ hybridization (ISH), we demonstrated the mutually exclusive expression of VMAT1 in endocrine cells of the duct system and of VMAT2 in many cells of the islets of Langerhans. By confocal laser scanning microscopy, VMAT1-positive cells were identified as enterochromaffin (EC) cells and VMAT2-positive cells as β-cells. In PETs, VMAT1 was found exclusively in all serotonin-containing tumors. In contrast, VMAT2 expression was lost in many insulinomas, independent of their biological behavior. VMAT2 was expressed by some non-insulin-producing tumors. The mutually exclusive expression of VMAT1 in EC cells and of VMAT2 in β-cells suggests that both cell types store monoamines. Monoamine storage mediated by VMAT1 in EC cells is apparently maintained in EC cell tumors. In contrast, many insulinomas appear to lose their ability to accumulate monoamines via VMAT2.
Keywords
G
These biogenic amine-containing neuroendocrine cells express specific proteins that enable them to store and secrete monoamines. These include plasma membrane transporters that scavenge and recycle monoamines from the extracellular space and intracellular membrane transporters for monoamine uptake and storage in secretory granules. Visualization of monoamine uptake mechanisms in neuroendocrine cells provides insights into their cell differentiation during development and in transformed cells and enables the role of monoamine-storage mechanisms to be studied in both neoplastic and non-neoplastic disease.
The uptake of monoamines from the cytoplasm into secretory granules is mediated by vesicular monoamine transporters (VMATs) and requires an ATPase-generated proton gradient (Erickson et al. 1996; Eiden 2000; Erickson and Varoqui 2000; Eiden et al. 2002). Two isoforms of the vesicular monoamine transporter (VMAT1 and VMAT2) have been characterized in rodent and human tissue (Erickson et al. 1992; Liu et al. 1992; Erickson and Eiden 1993). Extensive studies employing IHC and ISH have shown that these two transporters are differentially expressed in monoamine-handling cells of the central nervous system, the autonomic nervous system, the hematopoietic system, and the neuroendocrine cell system. The expression of the two transporters in a given cell type is usually, but not always, mutually exclusive (Weihe et al. 1994; Peter et al. 1995; Erickson et al. 1996; Fon et al. 1997; Hansson et al. 1998; Schütz et al. 1998; Eiden 2000; Weihe and Eiden 2000; Eiden et al. 2002).
VMAT1 is expressed in the serotonin-producing EC cells throughout the GI tract, and VMAT2 in the histamine-accumulating ECL cells of the oxyntic mucosa of the stomach (Weihe et al. 1994; Peter et al. 1995; Erickson et al. 1996; Hansson et al. 1998; Schütz et al. 1998). VMAT1 and VMAT2 are therefore excellent markers for identification of GI neuroendocrine tumors deriving from EC cells and ECL cells, respectively (Eissele et al. 1999; Rindi et al. 2000; Jakobsen et al. 2001).
The differential expression of VMAT1 and VMAT2 in the normal endocrine pancreas has never been examined in detail. Therefore, the question of whether VMAT1 and 2 are also useful in distinguishing among tumors originating from the pancreas is still open, although VMAT1 and VMAT2 protein expression in a small number of pancreatic endocrine tumors has been previously examined (Rindi et al. 2000; Jakobsen et al. 2001).
The aim of this study was to investigate VMAT1 and VMAT2 gene expression in the normal human and non-human primate pancreas by radioactive ISH and to determine the hormonal phenotype of the VMAT1- and VMAT2-positive pancreatic cell populations by confocal laser scanning microscopy and dual-color IHC. The expression profiles of VMAT1 and VMAT2 in the normal endocrine pancreas were used to determine whether VMAT1- and VMAT2-positive cell lineages could be distinguished in a large series of infants with hyperinsulinemic hypoglycemia and in pancreatic endocrine tumors in adults.
Materials and Methods
Origin of Tissues
Samples of non-neoplastic human pancreas were collected during abdominal surgery from patients suffering from pancreatic adenocarcinoma (two men and two women; mean age 64 years, range 58–79; classical Whipple resection in two cases and pancreatic tail resection in two cases). Pancreatic specimens were also obtained from five rhesus monkeys (Rausch et al. 1994). The specificity and sensitivity of the VMAT1 and VMAT2 antibodies and the VMAT1 and VMAT2 antisense probes were tested in various human and monkey tissues, including adrenal medulla, stomach, small intestine, one gastric ECL cell tumor, and two intestinal EC cell tumors.
Further non-neoplastic samples were obtained from the pancreas of 11 children with hyperinsulinemic hypoglycemia of infancy showing focal (n = 2) or diffuse (n = 9) β-cell hyperplasia. All children suffered from recurrent hypoglycemia and revealed high serum insulin levels.
Tumor tissue was obtained from 92 patients. All tumors were classified according to the WHO criteria (Solcia et al. 2000) including size, cell differentiation, angioinvasion, proliferation index, metastatic spread, hormone expression, and functional activity. PETs were classified as well-differentiated neuroendocrine tumors (NETs), well-differentiated neuroendocrine carcinomas (NECs), and poorly differentiated NECs.
A total of 61 of 92 patients with PETs had elevated serum levels of the respective hormones and exhibited specific clinical syndromes [insulinoma (n = 44), gastrinoma (n = 11), EC cell tumors (n = 2), VIPoma (n = 1), ACTH-secreting tumors (n = 2), and one poorly differentiated NEC with paraneoplastic hypercalcemia]. A total of 31 PETs were functionally inactive; they included five well-differentiated NETs, 22 well-differentiated NECs, and four poorly differentiated NECs.
Tissue Preparation
For IHC analysis the human tissues were fixed in either Bouin-Hollande fixative, 10% formalin, or 4% formaldehyde/PBS for 48 hr and then embedded in paraffin. The monkey tissues were perfused with 4% formaldehyde/PBS before postfixation in Bouin-Hollande for 24–48 hrs as described previously (Rausch et al. 1994).
After dehydration in a graded series of 2-propanol solutions, the tissues were embedded in Paraplast Plus (Merck; Darmstadt, Germany). Adjacent sections (3-μm or 7-μm thick) were cut and deparaffinized. Antigen retrieval to increase the sensitivity of immunodetection was performed by heating the sections at 92–95C for 15 min in 0.01 M citrate buffer (pH 6) according to the DAKO protocol (Hamburg, Germany). Nonspecific binding sites were blocked with 5% bovine serum albumin (BSA; Serva, Heidelberg, Germany) in PBS, followed by an avidin-biotin blocking step (avidin-biotin blocking kit; Boehringer, Ingelheim, Germany).
For ISH, human non-neoplastic pancreatic tissues and 11 endocrine tumors were immediately frozen on dry ice and stored at −80C. Cryosections (14–16-μm thick) were placed on pre-silanized glass slides, fixed in 4% phosphate-buffered formaldehyde for 60 min, followed by three 10 min washes in 50 mM PBS (pH 7.4). The slides were then briefly rinsed in distilled water, incubated in 0.1 M triethanolamine (pH 8.0) for 1 min, and for 10 min in the same solution containing 0.25% v/v acetic anhydride under rapid stirring. They were then quickly rinsed in 2 × SSC, dehydrated in 50 and 70% ethanol, and air-dried.
Immunohistochemistry
Rabbit antiserum no. VMAT1/10 raised against the C-terminal sequence of human VMAT1 and antiserum no. 80182 raised against the C-terminal sequence of human VMAT2 were employed for the immunohistochemical investigations (Table 1). Several antisera against general neuroendocrine markers, serotonin, and various peptide hormones were used (Table 1).
Tissue sections were incubated with the primary antibodies overnight at 18C (diluted as shown in Table 1) and further incubated for 2 hr at 37C. After washing in distilled water and in 50 mM PBS, sections were incubated with species-specific biotinylated secondary antibodies (Dianova; Hamburg, Germany) for 45 min at 37C, washed several times, and incubated for 30 min with the ABC reagents (Vectastatin Elite ABC kit; Boehringer). Immunoreactions were visualized with 3′3-diaminobenzidine (DAB; Sigma, Deisenhofen, Germany) enhanced by the addition of 0.08% ammonium nickel sulfate (Fluka; Buchs, Switzerland), resulting in dark blue staining. No binding was detected in the absence of the primary antibody. The specificity of the IHC staining was demonstrated by preabsorbing the antisera with 25 μmol of the C-terminal human VMAT1 and VMAT2 peptide, respectively. VMAT1 and VMAT2 were detectable in tissues fixed either in Bouin-Hollande or in 4% (para)formaldehyde with or without antigen retrieval.
The number of VMAT1- and VMAT2-positive cells in neuroendocrine tumors was estimated semiquantitatively as approximate percentage and scored on a scale from − (absent), (+) (extremely sparse), + (< 10%), ++ (10-<20%), +++ (20-<40%), ++++ (40-<60%) to +++++ (60–100%).
In Situ Hybridization
To generate specific probes for the localization of VMAT1 mRNA, a 247-bp-long DNA restriction fragment of the hVMAT1 cDNA (Erickson et al. 1996) corresponding to nt. 436–682 was subcloned into Bluescript II KS+ (Stratagene; Heidelberg, Germany). For VMAT2 mRNA detection, a 269-bp-long DNA fragment of the hVMAT2 cDNA (Erickson and Eiden 1993) corresponding to nt. 244–512 was sub-cloned into pcDNAI (Invitrogen; Leek, Netherlands). Inserted sequences were verified by double-stranded DNA sequencing. In vitro transcription of the two vector constructs using [35S]-UTP-labeled nucleotide yielded antisense riboprobes for hVMAT1 after linearization with XhoI and incubation with T7 RNA polymerase, and for hVMAT2 after linearization with XbaI and incubation with SP6 RNA polymerase. To increase the tissue penetration of probes, generated transcripts were reduced to about 200 nt. fragments by limited alkaline hydrolysis, as described by Angerer et al. (1987).
To each section hybridization buffer (3 × SSC, 50 mM NaPO4, pH 7.4, 1 × Denhardt's solution, 0.25 mg/ml yeast tRNA, 10% dextran sulfate, 50% formamide, 10 mM dithiothreitol) was applied. The hybridization mix contained 50,000 dpm/ml of 35S-labeled RNA probes. Sections were coverslipped and incubated in moist chambers at 60C for 16 hr. The coverslips were then removed in 2 × SSC. The sections were then subjected to the following posthybridization steps: RNAse treatment (20 μg/ml RNase A and 1 U/ml RNase T1 in 10 mM Tris, pH 8.0, 0.5 M NaCl, 1 mM EDTA) for 60 min at 37C and successive washes in decreasing salt concentrations (2 X, 1 X, 0.5 X, and 0.2 × SSC) for 10 min each, followed by incubation in 0.2 × SSC at 60C for 60 min. For autoradiography, slides were dipped in NTB-2 nuclear emulsion (Eastman Kodak; Rochester, NY) and developed after 3 weeks of exposure time. Developed sections were stained with hematoxylin and eosin, analyzed, and photographed in darkfield and brightfield modes with an Olympus AX 70 microscope (Hamburg, Germany).
List of primary antibodies
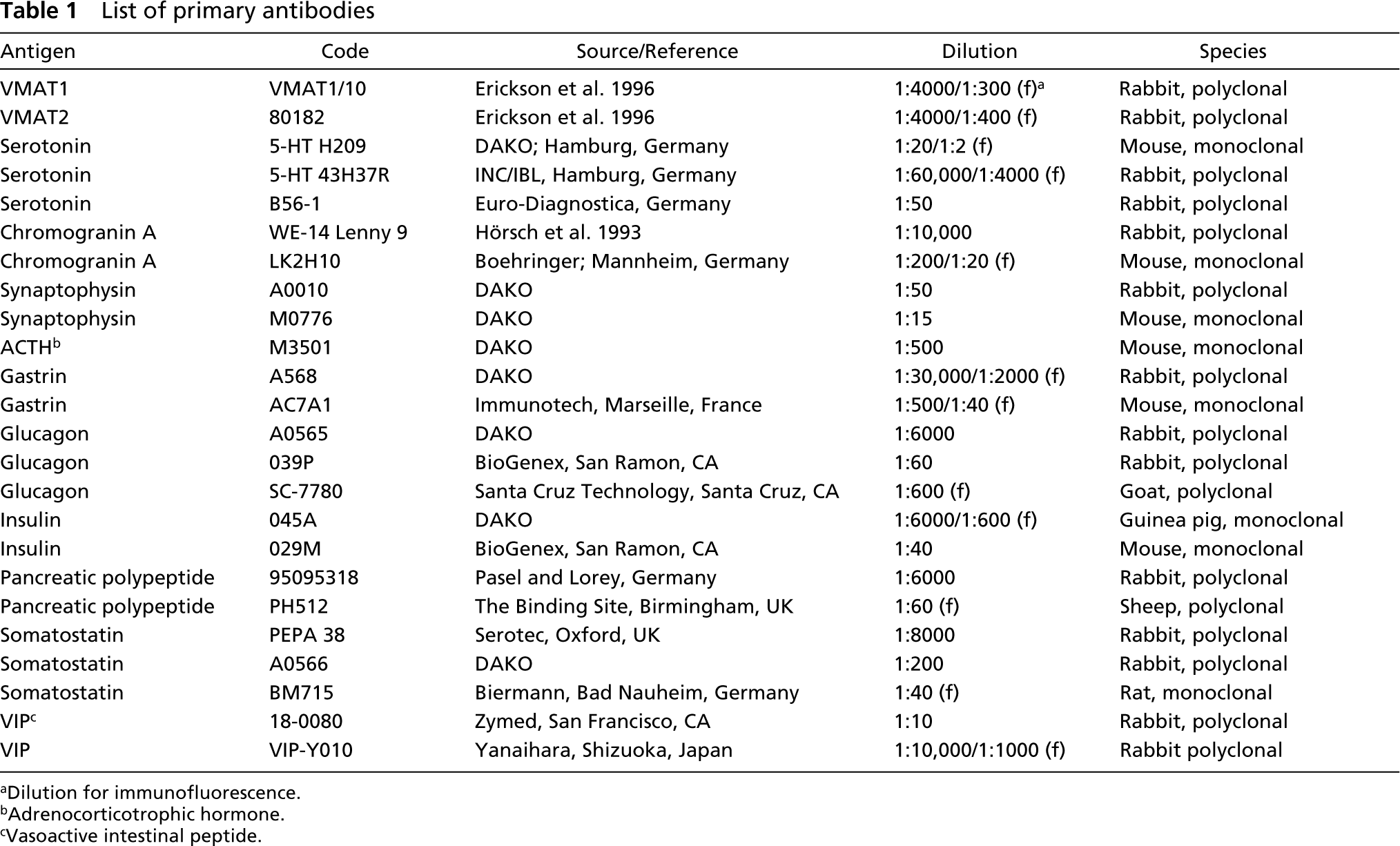
Dilution for immunofluorescence.
Adrenocorticotrophic hormone.
Vasoactive intestinal peptide.
Co-localization Studies
To study the co-localization of VMAT1 and VMAT2 with the respective neuroendocrine markers, three strategies were used: (a) alternate staining of adjacent semithin sections (2–3 μm thick), (b) the two-color immunoperoxidase technique (Hancock 1986), and (c) double-fluorescence labeling (Hörsch et al. 1993). The cellular distribution of antigens visualized by double fluorescence was analyzed by confocal laser scanning microscopy.
For adjacent section analysis, randomly selected pairs of semithin sections were stained for each marker combination as described above. For the two-color peroxidase technique, the first primary antibody was detected with the nickel-enhanced DAB procedure. After dehydration through a graded series of 2-propanol and one passage through xylene, the sections were rehydrated in a graded series of 2-propanol and treated with BSA and the avidin-biotin reagents to block potential nonspecific binding of the second avidin-biotin-peroxidase complex. The second primary antibody was then visualized by DAB/peroxidase reaction without nickel enhancement, resulting in a brown staining product. In control sections, the primary antibodies were omitted.
Double immunofluorescence detection was performed by covering the sections with a mixture of the two different primary antibodies in appropriate dilutions (Table 1) and by subsequent labeling with the species-specific secondary antibodies bearing the Alexa fluorochromes A647, A594, or A488 (MoBiTec; Göttingen, Germany) or the fluochromes Cy2 or Cy3 (Dianova; Hamburg, Germany). Furthermore, streptavidin coupled with Alexa fluochromes A647, A488, or A594 (MoBiTec) or with Cy2 or Cy3 was used in combination with biotinylated species-specific secondary antisera bearing the appropriate fluochrome. For conventional double immunofluorescence, sections were analyzed and photographed with an AX 70 microscope (Olympus) equipped with the appropriate filter cubes for discriminating among the different fluochromes). Confocal double immunofluorescence analysis was performed with the Fluoview laser scanning microscope (Olympus). Digital confocal images were presented in false color.
Ethics
The procurement of human material during surgery was approved by the Ethics Committee of the Medical Faculty of the University of Marburg. Oral informed consent was obtained from each patient before surgery. Rhesus monkey tissues, generously supplied by Dr. Lee E. Eiden, (NIMH, Bethesda, MD), were obtained in accordance with NIH/NIMH governmental rules.
Results
Mutually Exclusive Expression of VMAT2 in β-cells and VMAT1 in EC Cells of the Primate Pancreas
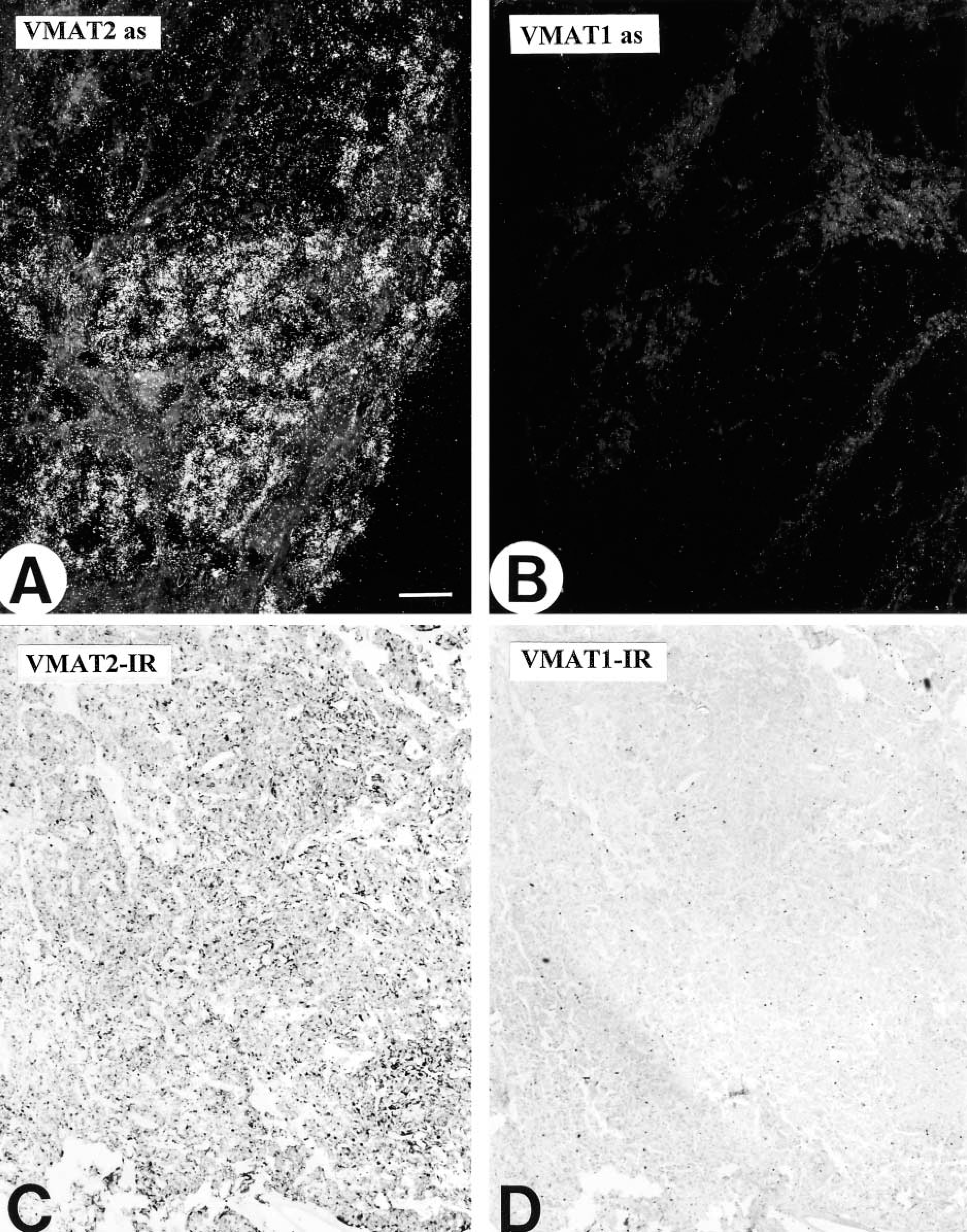
In all regions of the human and monkey pancreas examined in this study, VMAT2 was present in a considerable subpopulation of pancreatic islet cells, as demonstrated by IHC and ISH (Figure 1 and 2). VMAT2 was also detectable in varicose nerve fibers innervating the pancreatic islets, intra- and interlobular ducts, the exocrine parenchyma, and blood vessels (Figure 1). In contrast, islet cells did not reveal any positivity for VMAT1 and serotonin (data not shown). ISH revealed specific strong signals for VMAT2 in the majority of islet cells (Figure 2). In contrast, VMAT1 mRNA was lacking from pancreatic islets (Figure 2).
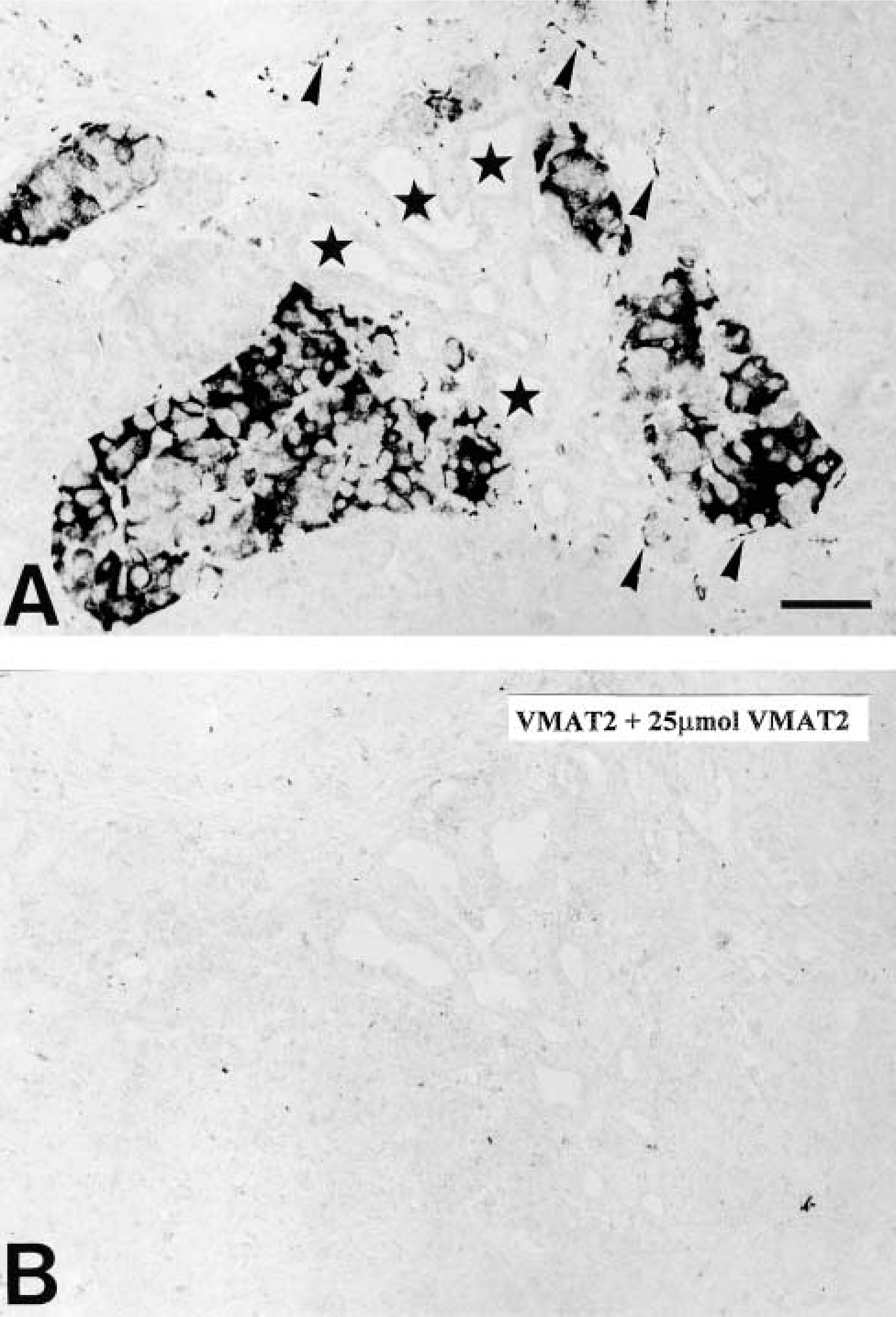
Immunohistochemistry for VMAT2 in human pancreas. (
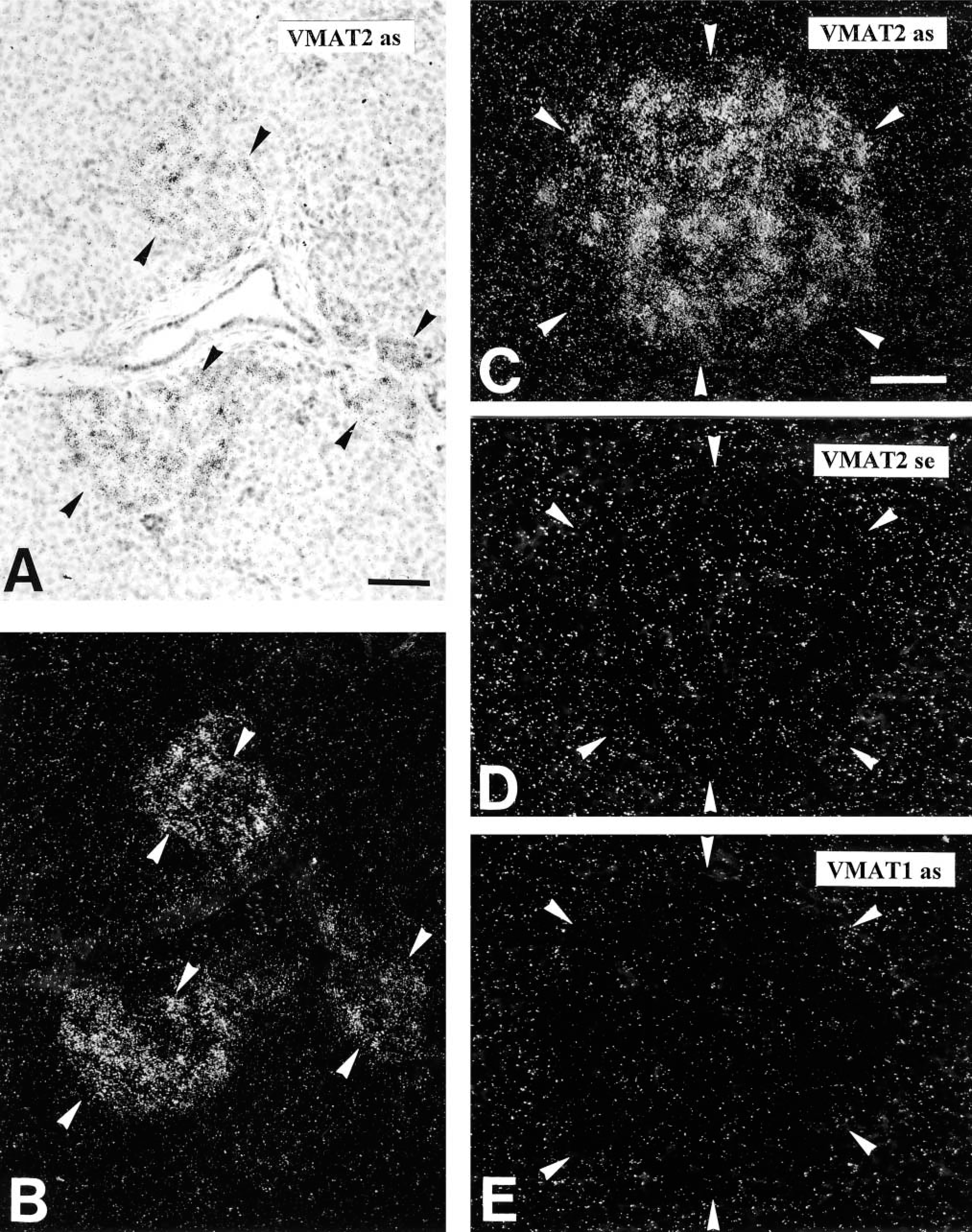
Comparison of VMAT1 and VMAT2 mRNA expression in the human pancreas. (
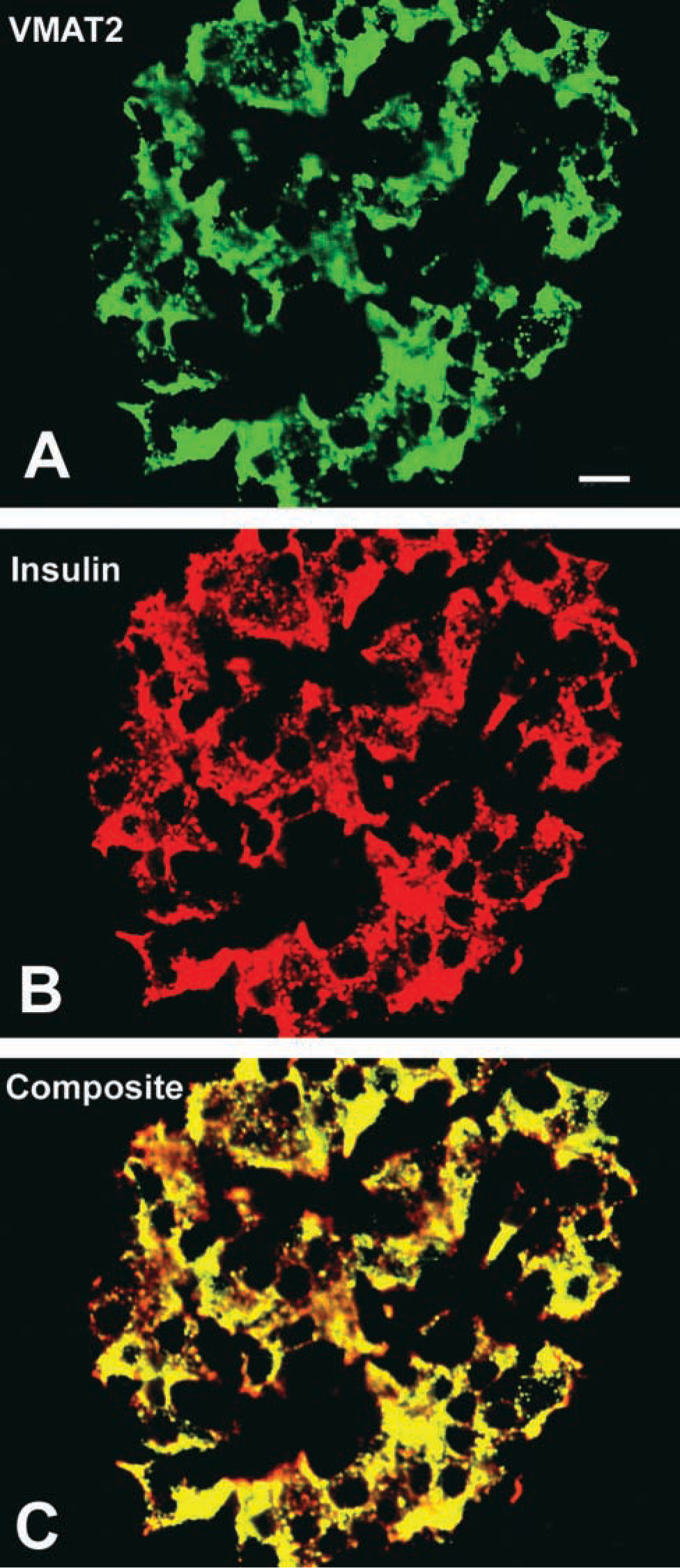
Identification of the cellular phenotype of VMAT2-immunoreactive cells in human pancreatic islets by confocal laser scanning microscopy. (
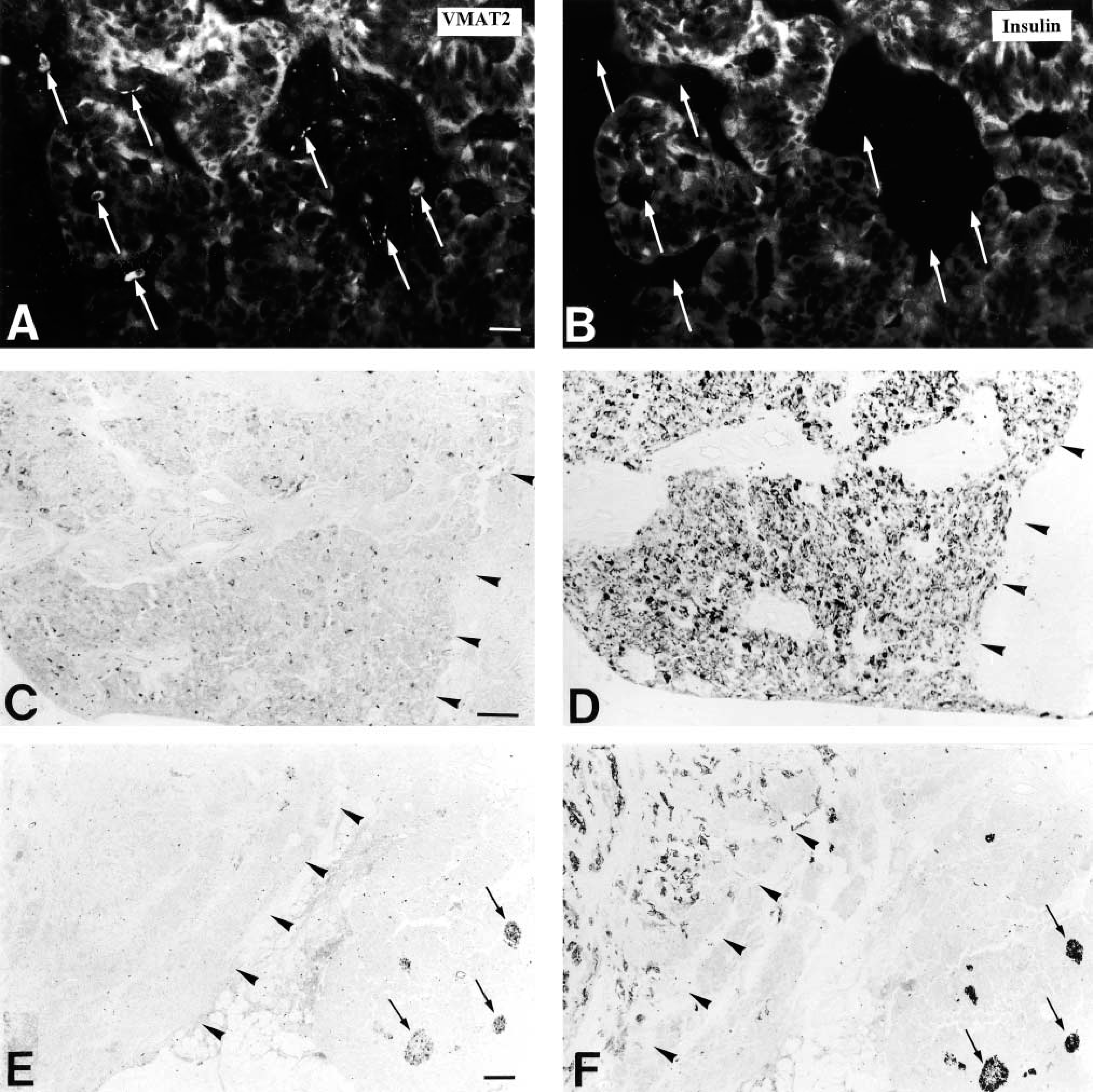
Confocal laser scanning microscopy demonstrated the presence of VMAT2 in insulin-immunoreactive cells (Figure 3). In contrast, VMAT2 was strictly absent from glucagon, somatostatin, and pancreatic polypeptide cells, as shown by the analysis of semithin adjacent sections, dual-color IHC, and confocal laser scanning microscopy (Figure 4). This observation was also confirmed in all cases of infants with hyperin-sulinemic hypoglycemia. In diffuse or focal β-cell hyperplasia all β-cells expressed exclusively VMAT2 but not VMAT1 (data not shown). VMAT1 was exclusively present in cells of the intra- and interlobular duct system in the pancreas. All VMAT1-immunore-active cells were co-positive for serotonin and CgA (Figure 5), but negative for insulin, glucagon, somatostatin, and pancreatic polypeptide.
The specificity and sensitivity of the VMAT1 antibody and the VMAT1 antisense probe were tested in various human and monkey tissues. VMAT1 protein and mRNA were easily visualized in these control tissues (data not shown). Neither VMAT1 nor VMAT2 immunoreactivity nor mRNA was expressed in pancreatic exocrine cells.
Loss of VMAT2 in Many Insulinomas, Expression of VMAT2 in Tumors Other than Insulinomas, and Conservation of VMAT1 in EC Cell Tumors
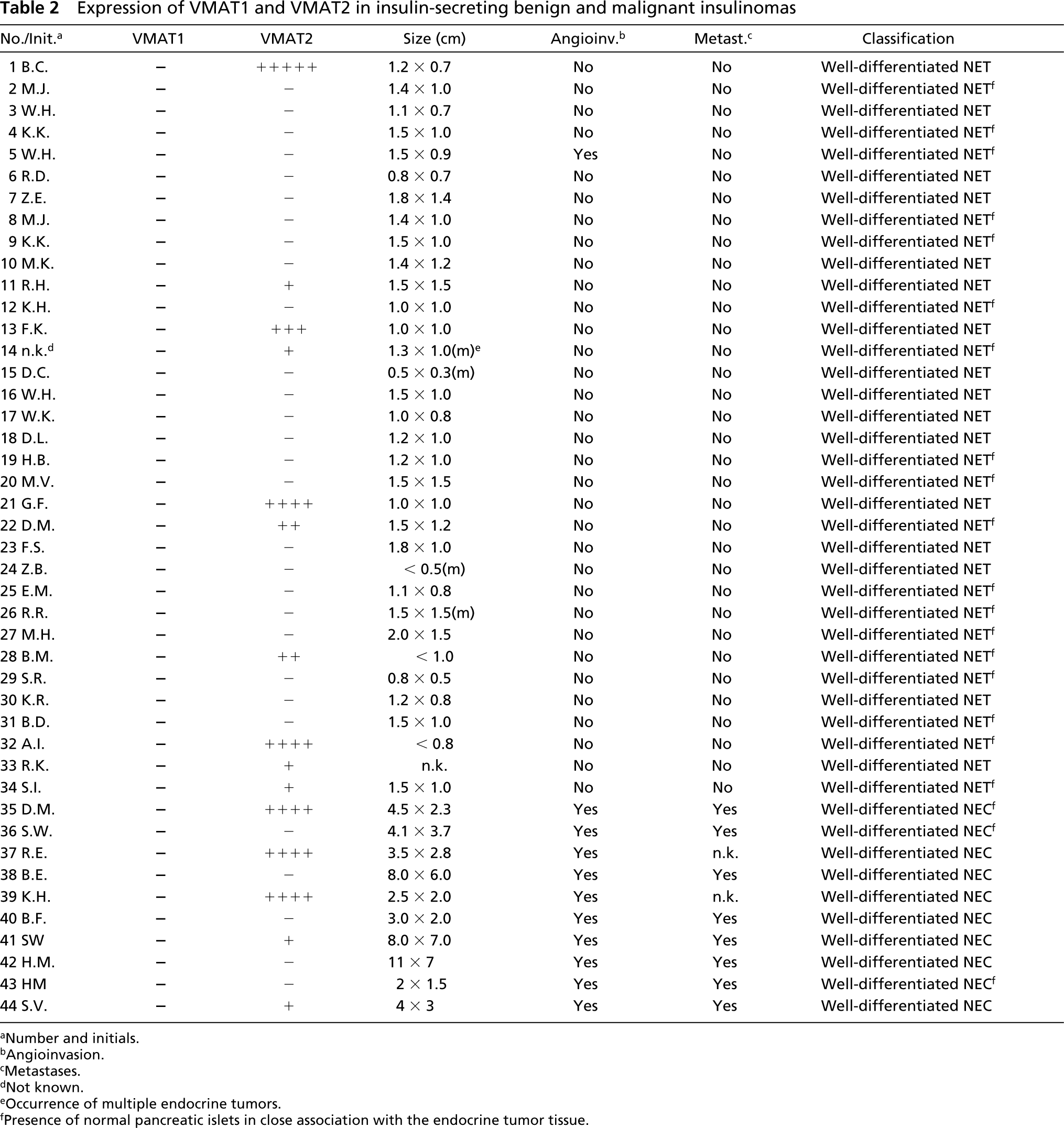
A minor proportion of insulinomas revealed expression of VMAT2, as demonstrated by IHC and ISH. VMAT2-positive cells were detected in 10 of 34 benign insulinomas and 5 of 10 malignant insulinomas (Figure 6; and Table 2). The VMAT2-positive cells were present in subpopulations of insulin-positive tumor cells and were distributed either diffusely or focally (Figure 7; and Table 2).
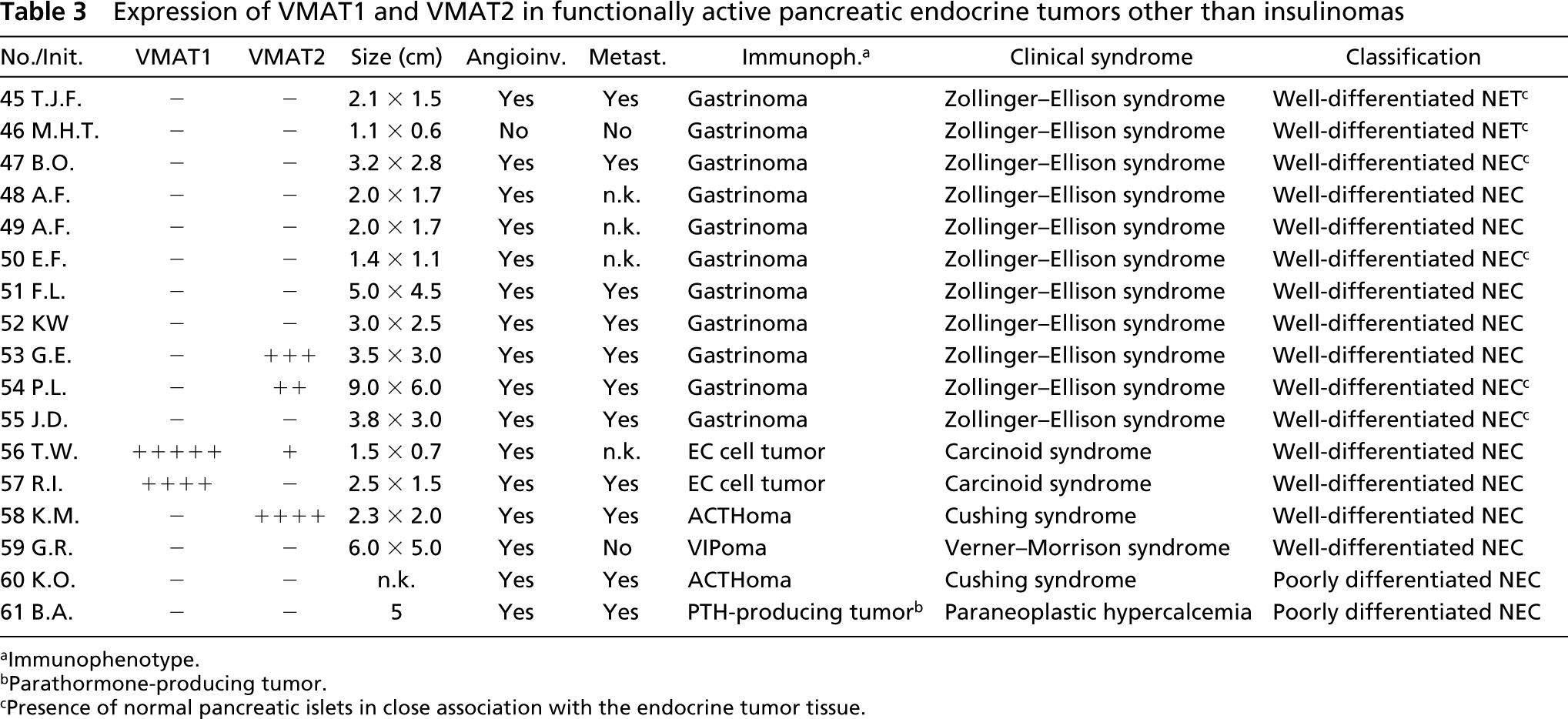
Some non-insulinomas revealed expression of VMAT2 in subpopulations of tumor cells. These consisted of two of 11 gastrinomas, one EC cell tumor, one ACTH-producing tumor, and eight of 31 nonfunctioning pancreatic neuroendocrine tumors (Tables 3 and 4). All beta cells in islets adjacent to the neuroendocrine tumors showed positivity for VMAT2 (Tables 2–4).
VMAT1 was absent from all but three PETs, which were found to express serotonin (Tables 2–4).
Discussion
The monoamine-containing neuroendocrine cell system is characterized by the expression of VMAT1 or VMAT2, which mediate monoamine uptake into secretory granules (Weihe and Eiden 2000; Eiden et al. 2002). In this study we demonstrated that in the normal endocrine pancreas only EC (serotonin-containing) cells express VMAT1 and only β-cells express VMAT2. Neoplastic beta cells (i.e. insulinoma cells), however, partially lose the feature of VMAT2 expression, while neoplastic EC cells (i.e. the cells of serotonin-containing tumors) retain the capacity to express VMAT1.
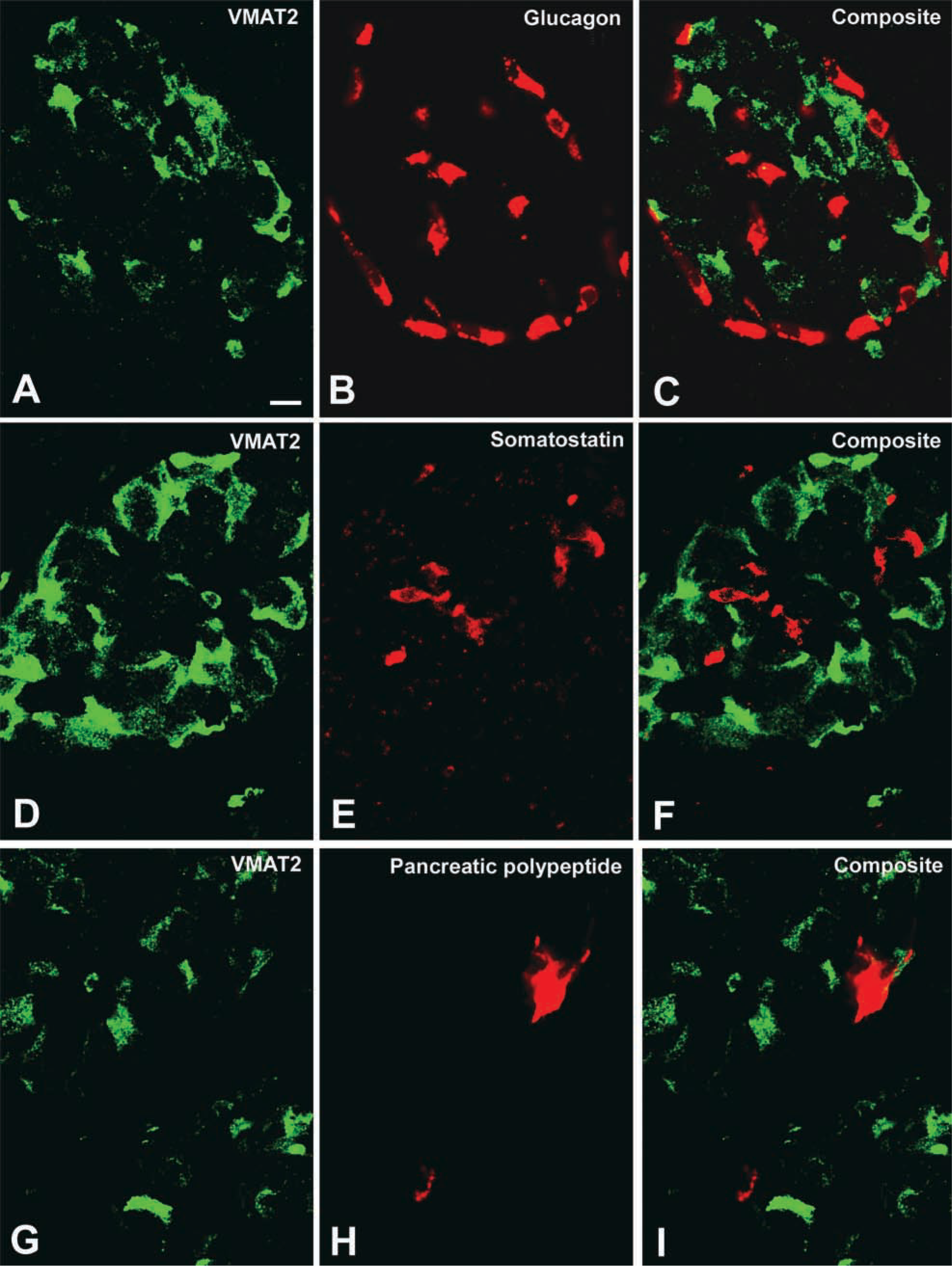
Absence of VMAT2 from glucagon, somatostatin, and pancreatic polypeptide cells demonstrated by confocal laser scanning microscopy. (
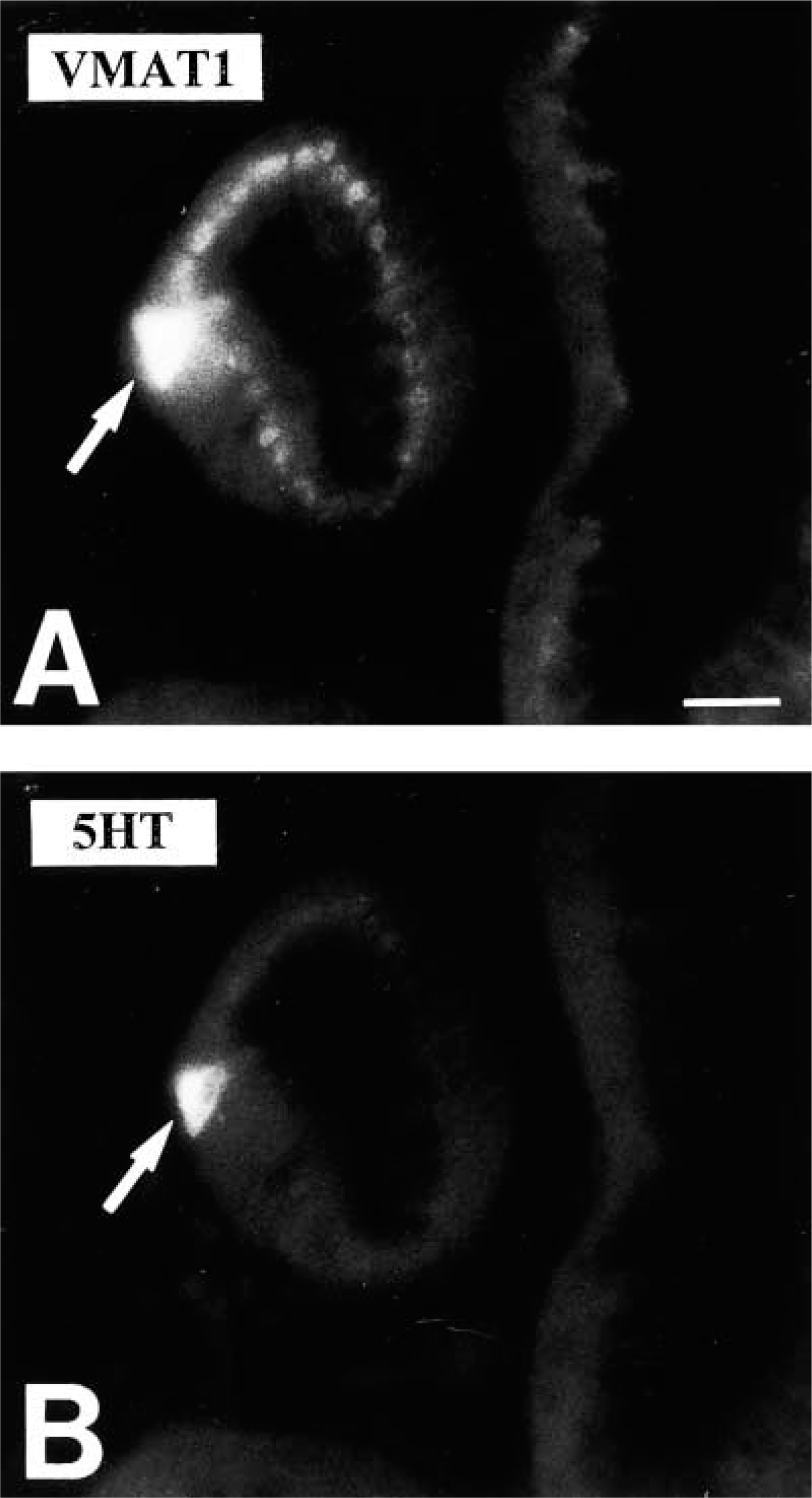
Identification of the cellular phenotype of VMAT1-immunoreactive cells in the rhesus monkey pancreatic duct system by double fluorescence analysis. (
Normal Pancreas and Hyperinsulemic Hypoglycemia of Infancy (Nesidioblastosis)
We demonstrated the exclusive expression of VMAT2 in normal beta cells in both humans and monkeys. The expression of VMAT2 in beta cells did not appear to be changed by hyperfunction, since the beta cells of children with hyperinsulinemic hypoglycemia of infancy showed the same expression pattern.
Vesicular monoamine transporters are responsible for the uptake of biogenic amine into secretory granules to make it available for exocytotic release (Erickson et al. 1996; Eiden 2000). The selective expression of VMAT2 in beta cells adds a new important feature to the functional spectrum of this particular cell type. Expression of VMAT2 is an indication of vesicular accumulation of monoamines, which are most likely released together with insulin from beta-cells. The physiological action of monoamines such as histamine, norepinephrine, dopamine and serotonin on the secretion of islet hormones is well documented (Lindstrom and Sehlin 1983; Ahren and Lundquist 1985; Jones et al. 1987; Sener et al. 1990; Persaud et al. 1993; Cable et al. 1995; Sjoholm 1995; Coulie et al. 1998; Yajima et al. 2001). Previous studies on the presence of monoamines, monoamine-synthesizing enzymes, and monoamine uptake in the islets of the non-primate pancreas (rat, mouse, hamster, and guinea pig) using various histochemical and chemical methods have yielded widely differing results (Mahony and Feldman 1977; Ahren and Lundquist 1985; Lundquist et al. 1989; Cetin 1992; Iturriza and Thibault 1993; Teitelman et al. 1993; Furuzawa et al. 1994; Barbosa et al. 1998). The specific phenotype and function of the monoamine(s) in normal primate β-cells remain to be identified. Our own observations in mouse, rat, and dog suggest marked species and developmental differences in the cell type-specificity of VMAT1 and VMAT2 expression in pancreatic islets, which fundamentally differ from human and rhesus monkey (Anlauf and Weihe, unpublished observations; and this study) and need to be fully explored in future studies. This underlines the unique situation in primates documented in the present study in which VMAT2 was found to be expressed in insulin-secreting β-cells but not in glucagon, somatostatin, and pancreatic polypeptide cells.
VMAT1 has been established as a marker of the normal EC cell in the gut (Weihe et al. 1994; Erickson et al. 1996). Scattered VMAT1-positive cells were previously observed in pancreatic islets closely associated with endocrine tumor tissue (Jakobsen et al. 2001). However, in our investigation of the normal primate endocrine pancreas, VMAT1-immunoreactive and VMAT1 mRNA-expressing cells were detected only in cells of the inter- and intralobular ducts, and not in or surrounding pancreatic islets themselves or in individual hormone-secreting cells found throughout the exocrine pancreas. The VMAT1-positive cells of the pancreas are neuroendocrine cells, based on their chromogranin A-positive phenotype. Double-fluorescence analysis revealed that these VMAT1-positive duct-associated cells contain serotonin. Their distribution corresponded to serotonin-producing cells described by others in the pancreatic ducts (Carlei et al. 1993). Regarding the function of VMAT1, it is of interest that recent studies in the guinea pig and rat indicate that serotonin-positive cells in the pancreatic ducts most likely mediate the fluid secretion of duct cells expressing serotonin receptors (Masuda et al. 1997; Li et al. 2000; Suzuki et al. 2001).
Distribution of VMAT1 and VMAT2 in an insulinoma investigated by IHC and ISH. (
Expression of VMAT1 and VMAT2 in insulin-secreting benign and malignant insulinomas
Number and initials.
Angioinvasion.
Metastases.
Not known.
Occurrence of multiple endocrine tumors.
Presence of normal pancreatic islets in close association with the endocrine tumor tissue.
Pancreatic Endocrine Tumors (PETs)
Based on the mutually exclusive expression of VMAT1 in EC cells within the pancreatic duct system and of VMAT2 in the β-cells of the islets of Langerhans, we anticipated that VMAT1 and VMAT2 might also serve as markers for the neoplastic counterparts of these cells in the neuroendocrine tumors of the pancreas. This is indeed the case for VMAT1, which labeled only the three serotonin-positive neoplasms in our series of 92 different PETs, including all relevant types belonging to this tumor group. We can therefore extend the marker function of VMAT1 from EC cell tumors of the gut (Eissele et al. 1999) to the same tumor type in the pancreas. Concerning VMAT2, it has been reported that it occurs not only in insulinomas, but also in other functioning and nonfunctioning PETs (Rindi et al. 2000; Jakobsen et al. 2001). We extend this observation by showing that approximately one third of insulinomas express VMAT2, whether they are benign or malignant.
Differential expression of VMAT2 in three different types of insulinomas investigated by IHC. (
Expression of VMAT1 and VMAT2 in functionally active pancreatic endocrine tumors other than insulinomas
Immunophenotype.
Parathormone-producing tumor.
Presence of normal pancreatic islets in close association with the endocrine tumor tissue.
Expression of VMAT1 and VMAT2 in non-functioning pancreatic endocrine tumors

Insulin.
Pancreatic polypeptide.
Glucagon.
Somatostatin.
Gastrin.
Serotonin.
Synaptophysin.
Chromogranin A.
Presence of normal pancreatic islets in close association with the endocrine tumor tissue.
We do not yet know which additional features distinguish the two thirds of insulinomas that do not express VMAT2 from those that do express VMAT2. However, it has long been known that the presence of typical or atypical secretory granules and the abnormal conversion of proinsulin to insulin in insulinoma cells reflect the state of differentiation of the tumor cell, compared with normal β-cells (Creutzfeldt et al. 1973; Capella et al. 1977; Roth et al. 1992,1995; Azzoni et al. 1998; Komminoth et al. 1999). Electron microscopic analysis and examination of hormone processing of VMAT2-positive and VMAT2-negative insulinomas will be required to determine whether VMAT2 expression is correlated with a specific granule phenotype or abnormal prohormone processing pathways.
VMAT2 positivity in non-insulinomas was not restricted to a certain tumor type, but it occurred only in malignant tumors. However, the number of benign PETs among the non-insulinoma cases was rather small. Therefore, this finding needs to be confirmed in a larger series. PETs are frequently multihormonal. Functional and non-functional PETs may therefore contain cells that are positive for insulin, in addition to normal pancreatic cells entrapped within the tumor. We carefully screened all PETs for insulin and VMAT2. We failed to detect VMAT2 positivity even in the scattered cells of these tumors, indicating that the insulin-positive cells in neuroendocrine tumors differ from the β-cells present in the islets of Langerhans and VMAT2-positive insulinomas. There is no clear correlation between a given function of PETs and VMAT2 expression, suggesting that neither the gain nor the loss of the capability to accumulate and secrete monoamines via VMAT2 affects the functional status of neoplastic endocrine cells.
In summary, we have demonstrated the mutually exclusive expression of VMAT1 in EC cells of the duct system and of VMAT2 in β-cells of the normal primate pancreas. This expression pattern suggests that both cell types store and release monoamines. In PETs, however, only VMAT1 retains its marker function for EC cells, whereas VMAT2 can be found in both non-insulin-secreting tumors and in insulinomas. Therefore, VMA2, although not a definitive marker for determining the cell origin of pancreatic endocrine tumors, may provide a basis for diagnosis, imaging, and treatment of malignant and non-malignant nonfunctional PETs.
Footnotes
Acknowledgements
Supported by grants of the Volkswagen-Stiftung (to LEE and EW) and the Kempkes-Stiftung, Marburg, Germany (to EW).
We wish to thank Dorothee Bondorden, Hena Buck, Petra Lattermann, Maike Pacena, Anja Paulus, Kerrin Ingwersen, Elke Rodenberg-Frank, Petra Sack, and Marion Zibuschka for their excellent technical assistance and Heidemarie Schneider for the brilliant photo documentation. We are indebted to Prof M. Rothmund, Dr C. Nies, and Dr B. Stinner (Department of General Surgery, University of Marburg) for providing some of the human pancreatic tissue. We thank K. Dege for critically reading the manuscript.