
Abstract
Monoamine oxidase (MAO) is regarded as a mitochondrial enzyme. This enzyme localizes on the outer membrane of mitochondria. There are two kinds of MAO isozymes, MAO type A (MAOA) and type B (MAOB). Previous studies have shown that MAOB activity is found in the pancreatic islets. This activity in the islets is increased by the fasting-induced decrease of plasma glucose level. Islet B cells contain monoamines in their secretory granules. These monoamines inhibit the secretion of insulin from the B cells. MAOB is active in degrading monoamines. Therefore, MAOB may influence the insulin-secretory process by regulating the stores of monoamines in the B cells. However, it has not been determined whether MAOB is localized on B cells or other cell types of the islets. In the present study, we used both double-labeling immunofluorescence histochemical and electron microscopic immunohistochemical methods to examine the subcellular localization of MAOB in rat pancreatic islets. MAOB was found in the mitochondrial outer membranes of glucagon-secreting cells (A cells), insulin-secreting cells (B cells), and some pancreatic polypeptide (PP)-secreting cells (PP cells), but no MAOB was found in somato-statin-secreting cells (D cells), nor in certain other PP cells. There were two kinds of mitochondria in pancreatic islet B cells: one contains MAOB on their outer membranes, but a substantial proportion of them lack this enzyme. Our findings indicate that pancreatic islet B cells contain MAOB on their mitochondrial outer membranes, and this enzyme may be involved in the regulation of monoamine levels and insulin secretion in the B cells.
Keywords
M
The pancreatic islet comprises numerous cell types that synthesize and secrete distinct peptide hormones (Erlandsen 1980; Reddy and Elliott 1988). Four major cell types are recognized in pancreatic islets of many mammalian species (including rat): A cells, which contain glucagon; B cells, which contain insulin; D cells, which contain somatostatin; and PP cells, which contain pancreatic polypeptide (PP) (Erlandsen 1980; Reddy and Elliott 1988). Biochemical studies have shown that MAOA and MAOB activities are present in homogenates of the pancreatic islets of rat, mouse, and golden hamster, with a greater predominance of MAOB activity (Feldman et al. 1980; Lenzen et al. 1987; Stenstrom and Lundquist 1990). In human pancreatic islets, there is an almost equal level of protein expression of MAOA and MAOB (Pizzinat et al. 1999). Immunohistochemical studies have shown that in guinea pig, MAOB-labeled cells appear to be evenly distributed throughout the islets (Kirchgessner and Pintar 1991), although in human, MAOA-positive cells are homogeneously distributed in the islets, and MAOB-positive cells are found in the periphery of the islets (Rodriguez et al. 2000). To our knowledge, which cell types of the pancreatic islets contain MAOA or MAOB remains to be elucidated. Furthermore, their subcellular localization has not been examined.
Recently, we raised and characterized a rabbit antiserum against bovine MAOB, which cross-reacts with rat MAOB (Sagara and Ito 1982; Arai et al. 2002). Using this antiserum, we have now performed double-labeling immunofluorescence histochemistry and electron microscopic immunohistochemistry to examine the cellular and subcellular localization of MAOB in the rat pancreatic islets.
Materials and Methods
Animals
Male Sprague-Dawley rats (n = 6, 180–200 g) were obtained from Japan SLC (Hamamatsu, Japan). All experiments were carried out according to the Guidelines for Animal Experimentation at Shiga University of Medical Science. All efforts were made to minimize both the number of animals used and their suffering.
Primary Antibodies
Sheep antibody against human glucagon (4660–0930, Biogenesis; Pool, England), guinea pig antibody against porcine insulin (A0564, DAKO; Carpinteria, CA), rat antibody against synthetic somatostatin (MAB354, Chemicon; Temecula, CA), and guinea pig antibody against rat PP (4040–01, Lincore-search; Charles, MO) were used. These antibodies have been previously characterized (Brar et al. 1989; Dun et al. 1994; Elayat et al. 1995; Zhang et al. 2000).
MAOB purified from bovine liver mitochondria was used to generate a rabbit anti-MAOB antiserum (Sagara and Ito 1982). This antiserum has been previously shown to immunoprecipitate MAOB of the rat liver (Sagara and Ito 1982). Immunohistochemical analysis has shown that this antiserum stains MAOB-containing cells but not MAOA-containing cells in the rat brain, indicating that the antiserum specifically recognizes rat MAOB (Arai et al. 2002).
Double-labeling Immunofluorescence Histochemistry
Three rats were anesthetized with sodium pentobarbital (60 mg/kg body weight, IP) and perfused through the ascending aorta with 50 ml of 0.01 M PBS (pH 7.4, room temperature), followed by 300 ml of a fixative solution containing 4% paraformaldehyde in 0.1 M phosphate buffer (PB, pH 7.4, 4C). The pancreases were dissected and placed in the same fixative solution for 24 hr at 4C and then immersed in PB containing 15% sucrose for 24 hr at 4C. Thirty-μm thick sections were cut using a cryostat (Cryocut 1800, Reichert-Jung; Heidelberg, Germany) and collected in PBS.
Double labeling of MAOB and each of four islet hormones was performed as follows. Sections were incubated in the following solutions: (a) 0.3% Triton X-100 in PBS for 1 hr at room temperature; (b) 5% normal donkey serum in PBS for 1 hr at room temperature; (c) a mixture of primary antisera:rabbit antiserum against MAOB (1:20,000 in PBS) combined with one of the four antisera:sheep anti-glucagon antiserum (1:200), guinea pig anti-insulin antiserum (1:20,000), rat anti-somatostatin antiserum (1:1,000), and guinea pig anti-PP antiserum (1:12,000), the mixture containing 1% normal donkey serum for 48 hr at 4C; and (d) a mixture of secondary antisera—Cy3-conjugated donkey anti-rabbit IgG (AP182C, Chemicon, 1:200 in PBS) combined with one of the three antisera:fluorescein-conjugated donkey anti-sheep IgG (AP184F, Chemicon, 1:200), fluorescein-conjugated donkey anti-guinea pig IgG (AP193F, Chemicon, 1:200), and fluorescein-conjugated donkey anti-rat IgG (AP189F, Chemicon, 1:200) for 2 hr at room temperature. After each incubation step, the sections were rinsed in PBS for 30 min at room temperature, mounted on glass slides, and cover slipped with a medium containing 90% glycerol and 0.1% p-phenylenediamine in PBS. Finally, MAOB was labeled with Cy3 (red), and glucagon, insulin, somatostatin, and pancreatic polypeptide were labeled with fluorescein (green). The stained sections were observed under a confocal laser scanning microscope (LSM410, Zeiss; Jena, Germany) with appropriate excitation laser beams and emission filters (for Cy3, excitation at 543 nm, emission at >570 nm; for fluorescein, excitation at 488 nm, emission at 510–525 nm). The Cy3 and fluorescence images were stored in memory, and then the superimposition of the two images was made and stored. These images were transferred to a Macintosh computer (Apple Computer; Cupertino, CA) equipped with Photoshop software (Adobe System; San Jose, CA) and printed on a color printer (Pictrography 3500, Fujifilm; Tokyo, Japan).
Electron Microscopic Immunohistochemistry
Three rats were anesthetized with sodium pentobarbital (60 mg/kg body weight, IP) and perfused through the ascending aorta with 50 ml of 0.01 M PBS (room temperature), followed by 300 ml of a fixative solution containing 4% paraformaldehyde, 0.3% glutaraldehyde, and 0.2% picric acid in 0.1 M PB (4C). The pancreases were dissected and placed in another fixative solution containing 4% paraformaldehyde and 0.2% picric acid in PB for 24 hr at 4C. Fifty-μm thick sections were cut using a microslicer (Dosaka; Kyoto, Japan) and collected in PBS.
For MAOB staining, an immunoperoxidase technique using avidin-biotin-peroxidase complex (Hsu et al. 1981) was performed. Sections were incubated in the following solutions: (a) 0.001% trypsin (type III, Sigma; St Louis, MO) in PBS for 5 min at room temperature; (b) 5% normal goat serum in PBS for 1 hr at room temperature; (c) rabbit anti-MAOB antiserum (1:10,000 in PBS) with 1% normal goat serum for 48 hr at 4C; (d) biotinylated goat anti-rabbit IgG (BA-1000, Vector; Burlingame, CA, 1:1,000 in PBS) for 2 hr at room temperature; (e) avidin-biotin-peroxidase complex (PK-4000, Vector, 1:1,000 in PBS) for 2 hr at room temperature; and (f) 0.025% 3,3'-diaminobenzidine (Dojindo; Kumamoto, Japan), 0.6% nickel ammonium sulfate (Nacalai Tesque; Kyoto, Japan), and 0.0075% hydrogen peroxide (H2O2) in 0.05 M trizma hydrochloride (Tris-HCl) buffer (pH 7.6) for 5 min at room temperature. The stained sections were fixed with 1% osmium tetroxide (Nacalai Tesque) in PB for 1 hr at 4C, dehydrated, and flat-embedded in epoxy resin (Luveak-812, Nacalai Tesque). Small areas containing the pancreatic islets were trimmed from the embedded section and cut on an ultramicrotome (Ultracut UCT, Leica; Heidelberg, Germany). Ultrathin sections were collected on grids (200 Cu, VECO; Eerbeek, Holland), stained with 2% uranyl acetate (Merck; Darmstadt, Germany) for 20 min and lead stain solution (Sigma-Aldrich Japan; Tokyo, Japan) for 5 min at room temperature, and examined with a transmission electron microscope (H-7100TE, Hitachi; Tokyo, Japan). Cell types of pancreatic islets were identified by the characters of their granules (Larsson et al. 1976; Roth et al. 1981; Weaver et al. 1986).
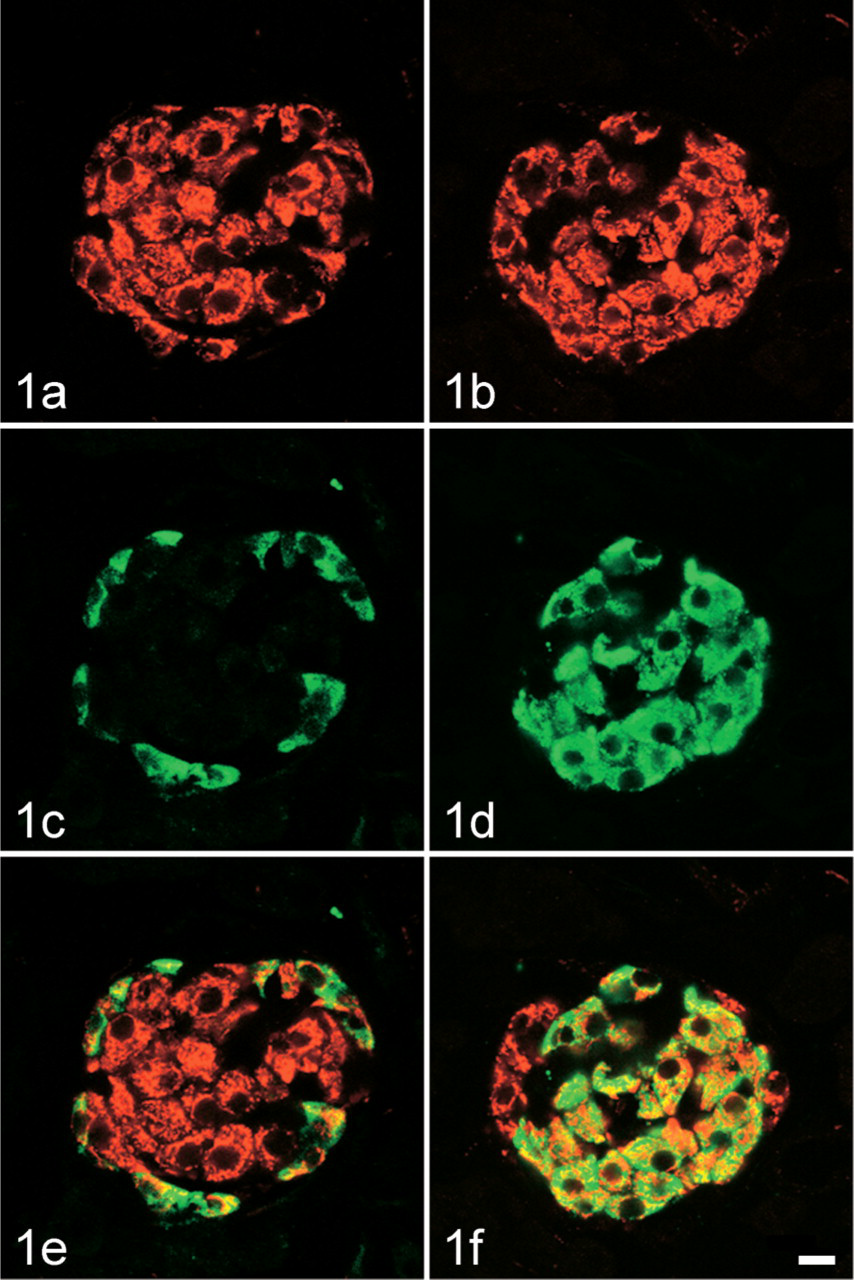
Immunohistochemical examination of monoamine oxidase type B (MAOB) localization in A cells and B cells of the rat pancreatic islet. Sections were processed for a double-labeling immunofluorescence method in combination of anti-MAOB (Cy3) with either anti-glucagon (fluorescein) or anti-insulin (fluorescein), and then observed with a confocal laser scanning microscope. (
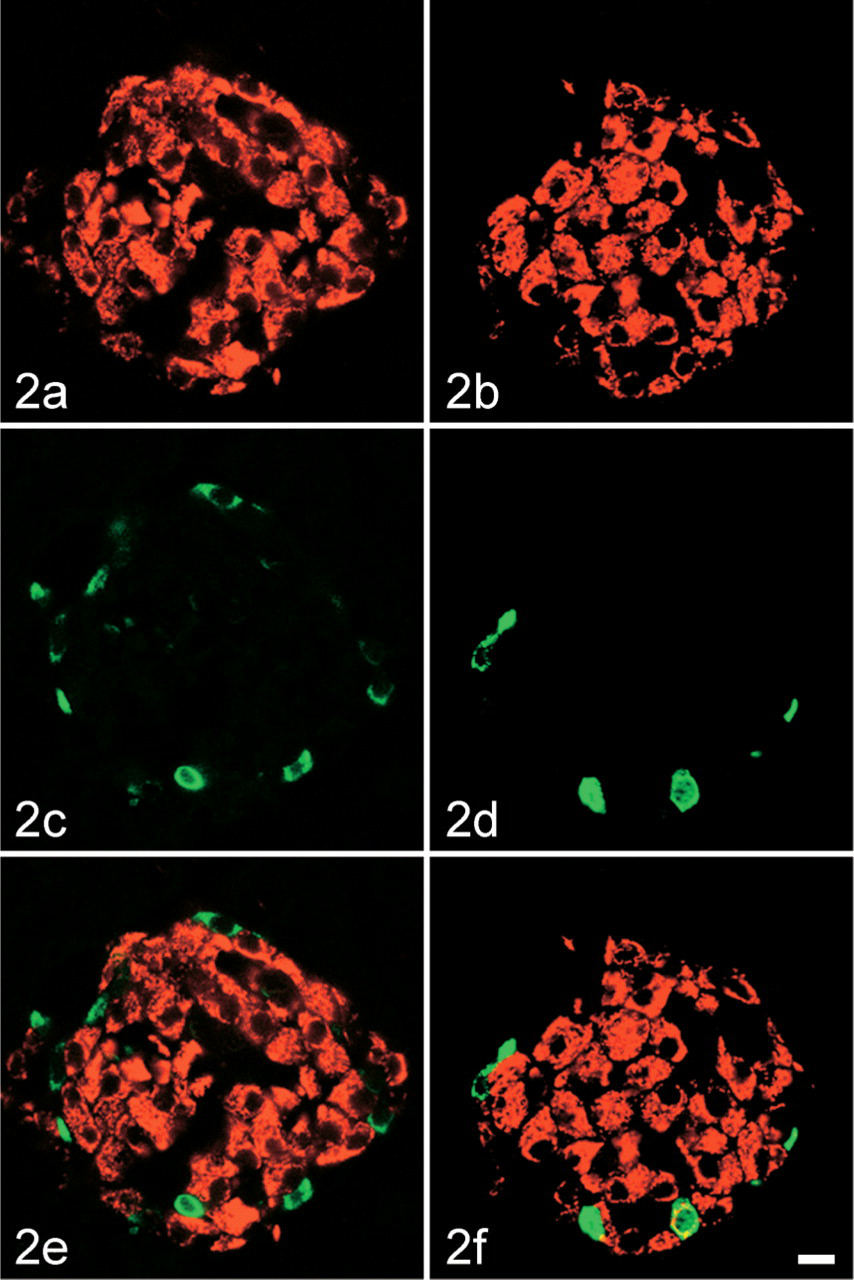
Immunohistochemical examination of monoamine oxidase type B (MAOB) localization in D cells and pancreatic polypeptide (PP) cells of the rat pancreatic islet. Sections were processed for a double-labeling immunofluorescence method in combination of anti-MAOB (Cy3) with either anti-somatostatin (fluorescein) or anti-PP (fluorescein), and then observed with a confocal laser scanning microscope. (
Results
Double-labeling Immunofluorescence Histochemistry
Figure 1 and Figure 2 show the cellular localization of MAOB in the pancreatic islet. All of the A cells that were labeled for glucagon (total number counted = 39, number of islets examined = 3) were also stained for MAOB (Figures 1A, 1C, and 1D). All of the B cells that were labeled for insulin (total number counted = 153, number of islets examined = 4) were also stained for MAOB (Figures 1B, 1D, and 1F). All of the D cells that were labeled for somatostatin (total number counted = 18, number of islets examined = 3) were negative for MAOB (Figures 2A, 2C, and 2D). Fifteen percent of PP cells that were labeled for PP (total number counted = 48, number of islets examined = 12) were positive for MAOB, but 85% of PP cells were negative for MAOB (Figures 2B, 2D, and 2F).
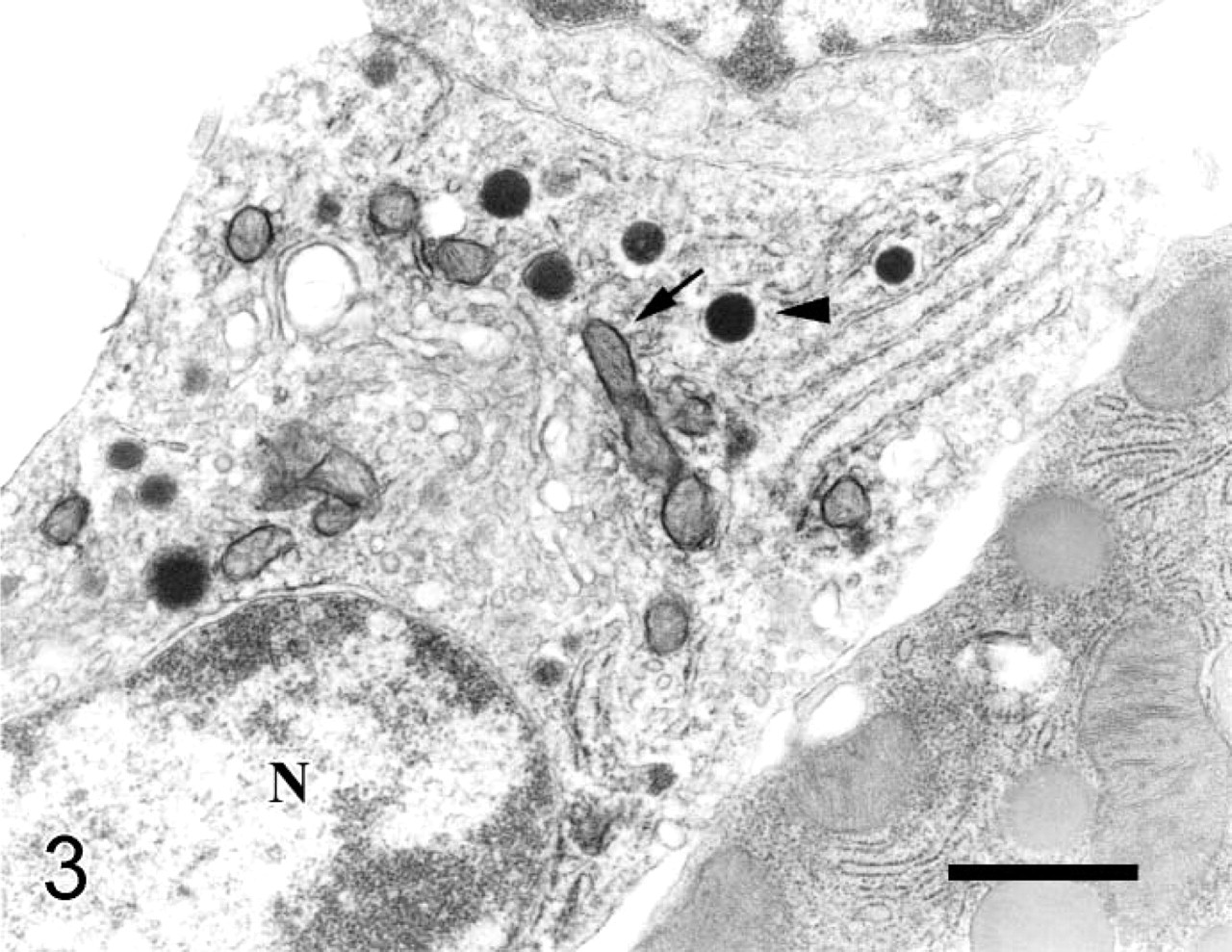
Subcellular localization of monoamine oxidase type B (MAOB) in pancreatic islet A cells. Sections were stained for MAOB with an immunoperoxidase technique and then processed for electron microscopy. Note that MAOB immunoreactivity is found on mitochondrial outer membranes (arrows) of A cells (N, nucleus), which contain electron-dense granules (arrowheads) that are round in shape and enclosed by small clear space. No MAOB immunoreactivity was found in acinar cells. Original magnification x 7200. Bar = 1 μm.
Electron Microscopic Immunohistochemistry
Figures 3–5 show the subcellular localization of MAOB in the pancreatic islet.
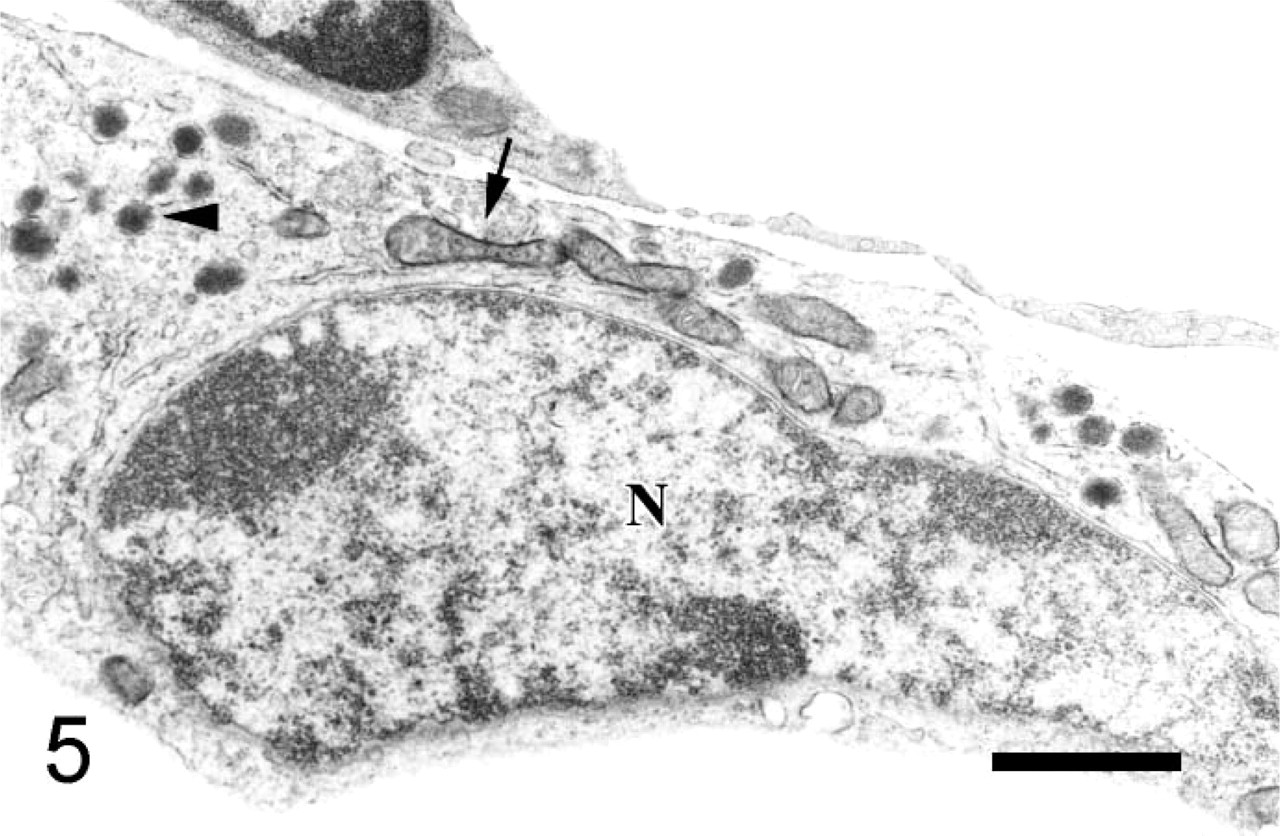
MAOB immunoreactivity is found on mitochondrial outer membranes in A cells, which contain electron-dense granules that are round in shape and enclosed by a smaller clear space (Figure 3); in B cells, which contain electron-dense granules that are mainly round and occasionally irregular in shape and enclosed by a larger clear space (Figures 4A and 4B); and in PP cells, which contain electron-dense granules that are smaller in size and slightly elongated in shape (Figure 5).
Discussion
We have found that MAOB is localized on mitochondrial outer membranes of almost all of both A cells and B cells and 15% of PP cells of the rat pancreatic islets. Virtually all D cells lack MAOB. The present study is the first to determine the cell types that contain MAOB and the subcellular localization of the enzyme in the pancreatic islets. These results are in general agreement with previously reported situations in pancreatic islets from rat, mouse, guinea pig, rabbit, golden hamster, and human populations (Aleyassine and Gardiner 1975; Feldman and Chapman 1975; Feldman and Henderson 1978; Lewinsohn et al. 1978,1980; Lenzen et al. 1983,1987; Egashira and Waddell 1984; Stenstrom and Lundquist 1990; Kirchgessner and Pintar 1991; Saura et al. 1992; Adeghate and Donath 1993; Panagiotidis et al. 1993; Pizzinat et al. 1999), which showed that both MAOA and MAOB were expressed in pancreas. However, we are not in agreement with Rodriguez et al. (2000), who studied MAO localization in human pancreas at light microscopic level. They reported that MAOA was observed in approximately 50% of human islet cells and probably corresponded to B cells, whereas MAOB was less abundant and restricted to the periphery of the islet. Therefore, they conjectured that MAOB-positive cells might correspond to A cells. The reasons may lie in a difference in tissues, antibodies, and methods. They used human tissue, mouse anti-human MAOA/MAOB, and peroxidase-anti-peroxidase technique. On the contrary, we use rat tissue, rabbit anti-bovine MAOB, and both double-labeling immunofluorescence histochemical and electron microscopic immunohistochemical methods.
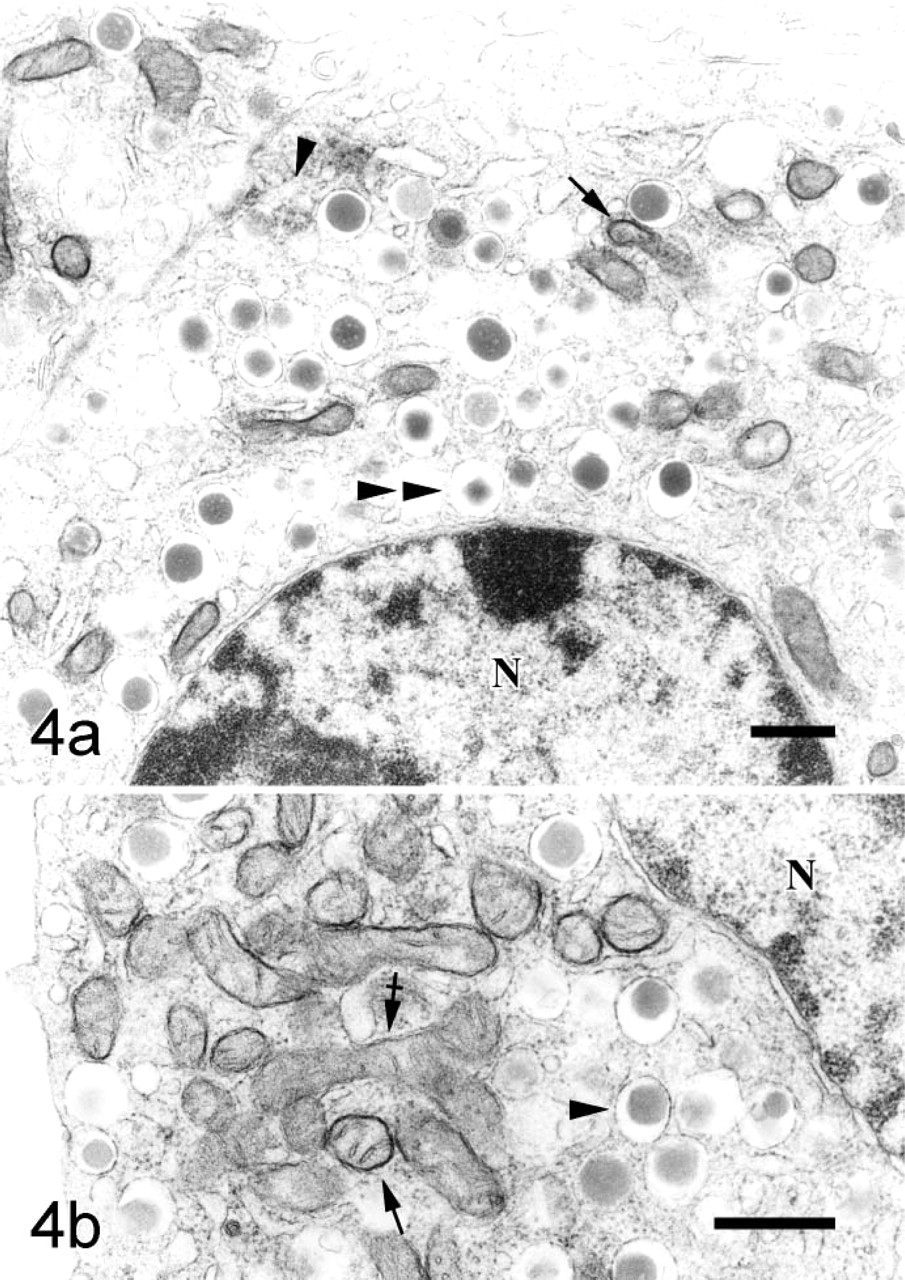
Subcellular localization of monoamine oxidase type B (MAOB) in pancreatic islet B cells. Sections were stained for MAOB with an immunoperoxidase technique and then processed for electron microscopy. Note that MAOB immunoreactivity is found on mitochondrial outer membranes (arrows) of B cells (N, nucleus), which contain electron-dense granules that are mainly round (arrowhead) and occasionally irregular (double arrowhead) in shape and enclosed by large clear space. Some mitochondria (crossed arrows) of B cells are negative for MAOB. Original magnification:
In our research, not all mitochondria exhibited a positive reaction, as judged by the formation of the electron dense precipitate. There were two kinds of mitochondria in pancreatic islet B cells: one contained MAOB on their outer membranes, but a substantial proportion of them lacked this enzyme. This finding may be attributable to one or more of the following factors (Shannon et al. 1974; Muller and Lage 1977): (a) different physiologic regions or states of the outer membranes, in which case the observed activity represents the true distribution of active MAOB; (b) uneven fixation that destroys some active enzyme sites; or (c) artificial, which was made by cutting. Further research is needed to understand the exact reasons for these differences.
Subcellular localization of monoamine oxidase type B (MAOB) in pancreatic islet pancreatic polypeptide (PP) cells. Sections were stained for MAOB with an immunoperoxidase technique and then processed for electron microscopy. Note that MAOB immunoreactivity is found on mitochondrial outer membranes (arrows) of PP cells (N, nucleus), which contain electron-dense granules (arrowheads) that are smaller in size and slightly elongated in shape. Original magnification x 7200. Bar = 1 μm.
MAO is synthesized in cytoplasmic polysome and inserted into the mitochondrial outer membrane (Berry et al. 1994). It appears that this enzyme is probably localized on both sides of the outer mitochondrial membrane (Russell et al. 1979). By radiochemical assay, Russell et al. (1979) determined MAO activity in human liver and brain-cortex nonsynaptosomal and synaptosomal mitochondria suspension. They found that MAO immunochemically accessible β-phenethylamine-oxidizing activity was situated predominantly on the outer surface, the immunochemically accessible 5-hydroxytrptamine oxidizing activity was situated predominantly on the inner surface, and the tyramineoxidizing activity was distributed on both sides of the mitochondrial outer membrane.
MAO has the ability to deaminate certain amines. Its substrate is relatively wide, including all primary, secondary, and tertiary monoamines (Berry et al. 1994). MAO plays a vital role in the metabolism of biogenic and xenobiotic amines in the central nervous system and peripheral tissues (Bach et al. 1988). It is the main degradative enzyme of monoamine hormones and amine neurotransmitters, such as epinephrine, norepinephrine, 5-hydroxytryptamine (5-HT), and DA (Rodriguez et al. 2000). The major function of MAO is the rapid inactivation of free monoamines, rather than the regulation of the content of total monoamines (Gey and Pletscher 1961). In addition, it plays a protective role for the body from the effects of other amines through the degradation of potentially toxic dietary amines and the inactivation of certain endogenous neurotransmitter substrates (Ryder et al. 1979; Thorpe et al. 1987).
In mammals, MAO has been identified in all cell types with the exception of the erythrocyte (Berry et al. 1994). Biochemical and histochemical results indicate large differences of MAO characteristics among animal species and in varying tissues. The specific localization of the two forms of MAO within different body tissues is of biological and clinical significance (Stenstrom and Lundquist 1990). The expression of MAO in pancreatic islets and that MAO inhibitors (clorgyline, deprenyl, pargyline, tranylcypromine, and amezinium) inhibit glucose-induced insulin secretion from isolated pancreatic islets (Lenzen et al. 1983) suggest that this enzyme may play an important regulatory role in the endocrine pancreas.
Previous studies have indicated that monoamines, such as 5-HT, DA, epinephrine, and norepinephrine, can inhibit glucose-mediated insulin secretion in human, golden hamster, rat, and mouse pancreatic islet B cells (Porte and Williams 1966; Feldman and Lebovitz 1970; Feldman et al. 1971; Aleyassine and Gardiner 1975; Lindstrom and Sehlin 1983). The catecholamines, epinephrine and norepinephrine, can regulate insulin secretion directly (Porte et al. 1976). When there is an increase in these sympathetic neurotransmitters, the secretion of insulin is inhibited initially. The initial decrease has been shown to be related to the activation of α2-adrenoceptoers on the B cells (Nakaki et al. 1980; Pizzinat et al. 1999). Other monoamines, such as 5-HT and DA, alter insulin secretion by acting as extracellular or intracellular agents. As extracellular agents, 5-HT and DA influence islet responses by stimulating cellular receptors directly or indirectly and by enhancing the release of norepinephrine from islet sympathetic terminals. As intracellular regulators, both 5-HT and DA have been localized within the islet B cells (Owman et al. 1973). Increased intracellular synthesis of these monoamines has been shown to decrease insulin output in response to standard stimuli (Porte et al. 1976)
In different mammalian species, pancreatic islet B cells have the ability to harbor biogenic monoamines, such as DA, 5-HT, and catecholamine, in their secretory granules (Cegrell 1967; Ekholm et al. 1971; Owman et al. 1973; Ericson et al. 1977; Pizzinat et al. 1999). MAO is the main degradative enzyme of monoamine hormones and amine neurotransmitters and might therefore have a regulatory influence on insulin secretion through its regulation of monoamine stores in the islet B cells (Stenstrom and Lundquist 1990). Panagiotidis et al. (1993) observed that, after an overnight fast in adult lean mice, islet MAO activity was increased and plasma level of glucose was markedly decreased. The data strongly suggest that glucose negatively modulates islet MAO activity. This, in turn, may affect insulin secretion through its effects on the monoamine content (Panagiotidis et al. 1993). However, the exact role of MAO in the regulation of granule-located monoamines levels has not yet been elucidated.
Furthermore, MAO might also act directly on the insulin-secretory mechanisms, because H2O2 evolution induced by islet MAO activity can affect the redox states of the B-cells’ glutathione system, the balance of which is known to influence glucose-induced insulin secretion (Panagiotidis et al. 1993; Pizzinat et al. 1999). However, the exact mechanism of MAO regulating insulin release directly is poorly understood.
The hormone insulin is stored in secretory granules and released from the pancreatic B cells by exocytosis (Wollheim et al. 1996). In glucose-stimulated B cells, both the cytosolic Ca2+ concentration ([Ca2+]c) rises and the insulin secretion are biphasic with a transient first phase and a sustained second phase (Bergsten 1995; Kennedy et al. 1996). The universal intracellular second messenger Ca2+ is the crucial trigger for the exocytosis of insulin (Wollheim 2000), the process by which the insulin-containing secretory granules fuse with the plasma membrane (Maechler and Wollheim 2000). In the consensus model of glucose-stimulated insulin secretion (Maechler and Wollheim 1999,2000), mitochondrial metabolism increase the cytosolic ATP/ADP ratio. This lead to closure of ATP-sensitive potassium channels and depolarization of the plasma membrane. Subsequently, the [Ca2+]c is raised by the opening of voltage-sensitive Ca2+ channels. The increase in [Ca2+]c trigger insulin exocytosis. Nevertheless, the Ca2+ signal alone is not sufficient for the full development of biphasic insulin secretion. A mitochondrial messenger must therefore exist that is distinct from ATP. In 1999, Maechler and Wollheim identified this as glutamate. They show that glucose generates glutamate from B-cell mitochondria. A membrane-permeant glutamate analog sensitizes the glucose-evoked secretory response, acting downstream of mitochondrial metabolism. In permeabilized cells, under conditions of fixed [Ca2+]c, added glutamate directly stimulates insulin exocytosis independently of mitochondrial function. These results demonstrate that glutamate acts as an intracellular messenger that couples glucose metabolism to insulin secretion (Maechler and Wollheim 1999).
MAO has the ability to deaminate certain amines. In the process of deamination, H2O2 and ammonia are generated (Berry et al. 1994). H2O2 alters mitochondrial activation and insulin secretion in pancreatic B cells (Maechler et al. 1999). Exposure of rat islets to H2O2 result in a retarded and sustained increase of [Ca2+]c (Maechler et al. 1999; Nakazaki et al. 2000), a transient increase in insulin release (Maechler et al. 1999). [Ca2+]c is the crucial trigger for the exocytosis of insulin (Wollheim 2000); therefore, the elevated basal insulin release is the consequence of an increase in [Ca2+]c (Maechler et al. 1999). Ammonia can be bound to the α-carbon atom of α-ketoglutarate, and glutamate is formed. This reversible reaction is catalyzed by glutamate dehydrogenase (Fisher 1985; Salway 1999). Glutamate uptake by B-cell granules would participate in the second sustained phase of insulin secretion (Maechler and Wollheim 2000). All this may be interpreted as evidence of the physiological role of MAO in the islet B cells. Up to now, the exact role of MAO in the regulation of monoamines levels and the mechanism of insulin release has not yet been elucidated and requires further investigation.