
Abstract
We studied the distribution of the homeodomain proteins Pdx-1 and Nkx 6.1 in the developing rat pancreas. During early development, nuclear staining for both Pdx-1 and Nkx 6.1 occurred in most epithelial cells of the pancreatic anlage. Subsequently, Nkx 6.1 became more β-cell-restricted, and Pdx-1 also occurred in other islet cell types and in the duodenal epithelium. During early pancreatic development, cells co-storing insulin and glucagon were regularly detected. The vast majority of these did not possess nuclear staining for either Pdx-1 or Nkx 6.1. Subsequently, cells storing insulin only appeared. Such cells displayed strongly Pdx-1- and Nkx 6.1-positive nuclei. Therefore, Nkx 6.1, like Pdx-1, may be an important factor in pancreatic development and in mature insulin cell function.
T
Recently, a mouse homeobox protein, insulin promoter factor 1 (ipf1), was found to be required for development of the murine pancreas (Jonsson et al. 1994; Offield et al. 1996). In addition, ipf1 may be important in regulating the transcription of the insulin genes (Ohlsson et al. 1993; Peers et al. 1994; Petersen et al. 1994; Serup et al. 1995, 1996). Ipf1 is homologous to the rat homeobox gene product stf1/idx1 (Leonard et al. 1993; Miller et al. 1994) and to the Xenopus laevis homeobox 8 (Peshevaria et al. 1994), and these proteins are now collectively referred to as pancreatic duodenal homeobox gene 1 (Pdx-1) (Stein et al. 1996). Homeobox proteins are involved in cell differentiation and tissue determination. A search for new homeobox genes in an insulinoma cDNA library led to the finding of several such genes, one of which (Nkx 6.1) appeared to have an islet cell-restricted distribution (Rudnick et al. 1994). Recent studies have revealed a β-cell-specific expression of Nkx 6.1 in the adult rat pancreas and have detected activation of this gene during progression of islet tumor cells to an insulinproducing phenotype (Jensen et al. 1996).
We decided to study the development of the islet hormone-producing cells in the rat pancreas in relation to the development of Pdx-1- and Nkx 6.1-immunoreactive cells to determine the relation of these homeobox gene products to specific hormonal phenotypes.
Materials and Methods
Animals
Wistar rats (Pan:WIST; The Panum Institute, Copenhagen, Denmark) were mated overnight; noon of the day on which the vaginal plug was discovered was considered as Day 0.5 of gestation (E0.5). Pregnant female rats were sacrificed by CO2 and the fetuses were immersion-fixed overnight in 4% paraformaldehyde in 0.1 M sodium phosphate buffer, pH 7.4. With embryos older than E16.5, the abdominal cavity was opened before fixation. Rat embryos were examined at E11.0, E11.5, and at 1-day intervals from E11.5 to E20.5. Pancreases from postnatal rats 1-13 days of age (P1-P13) were immersion-fixed overnight. Adult rats were intracardially perfused with the fixative and the pancreas immersion-fixed for 1-20 hr. Whole E11.0 and E11.5 embryos, the pancreatic region of E12.5 to E14.5 embryos, and the splenic and duodenal pancreases of E15.5 and older animals were cryoprotected overnight in 30% sucrose, mounted in Tissue-Tek, and frozen in N-hexane cooled by liquid nitrogen. Pancreatic tissue for RNA extraction was immediately transferred to 100 μl RNAzol extraction solution (Cinna/Biotecx; Houston, TX), homogenized, and RNA extracted as described by the manufacturer. At the earliest stage (E13.5), we obtained 2 μg total RNA from one full litter. At later stages, total yield increased.
Multiplex RT-PCR
Random-primed cDNA synthesis and multiplex RTPCR were performed as described earlier (Jensen et al. 1996). Primer sequences for Nkx 6.1 (284 bp), Pdx-1 (224 bp), TBP (TATA-binding protein) (190 bp), insulin (312 bp) (these primers amplify both rat insulin I and II), and tubulin (250 bp), were as described (Jensen et al. 1996). Primer sequences used to amplify rat glial-testis homeobox (Gtx) (160 bp; sequences derived from mouse Gtx, Genbank Acc. MUSGTX.GB_RO) were: upstream, TTTAGCCGCGCTGCACAACAT; downstream, CACCGGCCGTCCCAGCATGTC. This primer set amplifies rat and mouse Gtx to amplicons of the same size. Quantitation of product yield was performed by a Molecular Dynamics series 400 phosphorimager, as described (Jensen et al. 1996).
Immunocytochemistry
Sections of 3-5 μm were stained by indirect immunofluorescence using rabbit antisera to recombinant glutathione-S-transferase (GST) fusion proteins incorporating either the C-terminal region of Pdx-1 (stf1) (Ab. 1856-5) or the C-terminal region of Nkx 6.1 (Ab. 174) (Jensen et al. 1996). The recombinant proteins were produced in bacteria and purified by affinity chromatography on glutathione columns. The GST-Pdx-1 expression plasmid was a kind gift from Dr. J. Leonard (Strang-Cornell Research Laboratory; New York, NY) (Peers et al. 1994). In addition, sections were stained with rabbit antisera to glucagon (No. 4316; a kind gift from Dr. J.J. Holst, Department of Physiology, University of Copenhagen, Denmark), human pancreatic polypeptide (HPP, #615-1054B-248-4; kindly donated by Dr. R.E. Chance, Eli Lilly Co., Indianapolis, IN), guinea pig anti-insulin serum (recognizes both rat insulin I and II) (DAKO A/S; Glostrup, Denmark), and mouse monoclonal antisera to glucagon (GLU 001) or somatostatin (SOM018) (both from NovoClone; Novo-Nordisk, Bagsvaerd, Denmark) as previously detailed (Larsson et al. 1975,1976b; Jackerott et al. 1996). For double and triple immunofluorescence, mixtures of primary antibodies raised in different species were used for staining and detection was achieved by species-specific secondary antibodies conjugated to fluorescein isothiocyanate (FITC), Texas Red, amino-methylcoumarine (AMCA), or biotin (Jackson Immuno-Research Laboratories, West Grove, PA; DAKO A/S and Statens Bakteriologiska Laboratorium, Stockholm, Sweden). The biotin-conjugated antibodies were detected with AMCA-conjugated streptavidin (Vector Laboratories; Burlingame, CA). The secondary antibodies were absorbed overnight with 10μl/ml normal serum from the species recognized by the other secondary antisera, with 10μl/ml of normal serum from the species donating the other secondary antibodies (when applicable), and with 10 μl/ml normal rat serum.
Triple immunostainings for confocal microscopy were performed as above except that Cy5-labeled donkey anti-rabbit IgG (Jackson ImmunoResearch Laboratories) was used instead of AMCA-labeled antibody. The confocal laserscan microscope (Molecular Dynamics; Sunnyvale, CA) was equipped with a x 100, n.a. 1.40 Nikon objective and an argon/krypton laser. Scans were taken using a lateral x-y resolution of 0.13 μm and a z resolution of 0.39 μm. In this way, hormone and homeobox protein coexistence could be established in thin optical sections.
For quadruple stainings, sections were first stained for Nkx 6.1 or Pdx-1 by the peroxidase-anti-peroxidase (PAP) method using diaminobenzidine-H2O2 development (Sternberger 1979), and subsequently underwent triple immunofluorescence staining for islet hormones as above. Because the Nkx 6.1 and Pdx-1 stains were nuclear and islet hormone stains were cytoplasmic, no interference between immunofluorescence and PAP staining occurred. Sections used for PAP staining were first incubated in methanol-H2O2 to quench endogenous peroxidase activity, exposed to normal goat serum, and PAP-stained followed by detection of peroxidase activity by diaminobenzidine-H2O2.
Controls included conventional staining controls (Larsson 1988) as well as absorptions of the antisera with recombinant human insulin and monocomponent porcine glucagon (Novo-Nordisk), synthetic somatostatin-14, synthetic rat pancreatic polypeptide (Peninsula Laboratories; Merseyside, UK), and with the recombinant Nkx 6.1 and Pdx-1 proteins cleaved from the GST fusion protein. Our previous studies have established that the HPP antiserum does not crossreact with peptide YY and neuropeptide Y (Jackerott et al. 1996).
All staining and absorption controls were negative at all stages studied. Stainings with the Nkx 6.1 antiserum were eliminated by absorption against recombinant Nkx 6.1 but not with recombinant Pdx-1, whereas the reverse was true for the Pdx-1 antiserum. As a further control, double stainings for glucagon and PP employed poly-
Results
Nkx 6.1 and Pdx-1 Immunoreactivities Are Present in Most Developing Early Pancreatic Anlage Cells but Show Different Distributions in the Duodenum
At the earliest fetal stage studied (22 somites, approximately E11.0) Pdx-1- and Nkx 6.1-immunoreactive nuclei were detected in epithelial cells of the dorsal (Figures 1A-1D) and ventral pancreatic buds. The nuclear staining for Pdx-1 was more intense than that for Nkx 6.1. At this stage, few and weakly stained glucagon-immunoreactive cells were observed in the dorsal but not in the ventral bud. The nuclei of the early glucagon cells did not stain for either Pdx-1 or Nkx 6.1, whereas most of the other epithelial cells of the dorsal and ventral buds contained Pdx-1-and Nkx 6.1-positive nuclei. By the 30-somite stage (approximately E11.5) the first insulin-immunoreactive cells were observed. All of the early insulin cells were also glucagon-positive. Only a few of these cells contained weakly Pdx-1-and Nkx 6.1-immunoreactive nuclei. Similarly, at this stage, only a few cells staining for glucagon (but not insulin) possessed weakly Pdx-1- and Nkx 6.1-positive nuclei. With both types of cells the frequency of nuclei staining for Pdx-1 approximated 10%, whereas the frequency of nuclei staining for Nkx 6.1 was somewhat lower. However, because at early stages of development the staining for Nkx 6.1 was weaker than that for Pdx-1, this difference may be more apparent than real. At all ages studied (E13.5-16.5 and P54) the duodenal epithelial cells contained Pdx-1-positive nuclei, whereas Nkx 6.1 was absent.
Insulin Cells That Do Not Co-express Glucagon Are Characterized by Intense Nuclear Staining for Pdx-1 and Nkx 6.1
During E11.5-13.5, progressively more glucagon- and insulin-positive cells appeared. These cells were arranged in clusters of cells immunopositive for both insulin and glucagon or for glucagon only. By E13.5, the first insulin-positive cells that did not also stain for glucagonere observed. Such glucagon-negative insulin cells almost invariably displayed intense nuclear staining for Pdx-1 and Nkx 6.1. These cells were few up to E15.5 (Figures 1E-1L). Between E15.5 and about E18.5, some major changes in the number and organization of the different cell types were observed (Figures 1E-1L and 2A-2C). First, the number of single-positive insulin cells possessing Pdx-1- and Nkx 6.1-immunoreactive nuclei increased dramatically and, concomitantly, the number of cells immunopositive for both glucagon and insulin decreased. Second, the frequency of the (weakly) Pdx-1- and Nkx 6.1-positive glucagon cells decreased to the levels observed in adults (<1%). Third, the nuclear staining for Pdx-1 and Nkx 6.1 in the hormone-negative epithelial cells decreased in intensity, and by E18.5 Pdx-1- and Nkx 6.1-positive nuclei were mainly observed in duct epithelium and in insulin cells (Figures 2A-2C). Finally, at E15.5 and E16.5 the majority of the glucagon-negative and Pdx-1- and Nkx 6.1-positive insulin cells appeared as single cells, closely associated with the hormone-negative and Pdx-1- and Nkx 6.1-positive epithelial cells, whereas the insulin and glucagon double-positive cells were found mainly within clusters of glucagon cells (Figures 1E-1L). By E18.5, insulin-glucagon double-positive cells were rare, and the frequency of insulin cells now exceeded the frequency of glucagon cells. These insulin cells almost invariably displayed strongly Pdx-1- and Nkx 6.1-immunopositive nuclei and usually occurred in association with Pdx-1- and Nkx 6.1-positive epithelial duct cells (Figures 2A-2C). At this stage of development, the glucagon clusters assumed an elongated form, as if they were gradually beginning to form a mantle around the insulin cells. Formation of true islet-like structures could be observed during the following 2-3 days. In the adult pancreas, Pdx-1 and Nkx 6.1 immunoreactivities were restricted to islet cells (Figure 2D). In addition, extremely rare hormonenegative epithelial cells with Nkx 6.1- or Pdx-1-positive nuclei were occasionally detected in the vicinity of the islets, among exocrine cells, or in ducts.
Quantitation of Insulin, Pdx-1, Nkx 6.1, and Gtx mRNA Expression During Pancreatic Development
Pancreatic specimens from E13.5E17.5 were analyzed by multiplex RT-PCR. At the earliest stage studied (E13.5) and during the following 2 days of development, low levels of insulin gene expression were detected (Figure 3A). From E15.5 to E17.5, the levels of insulin mRNA increased about 100-fold (inset in Figure 3A). Subsequently, from E17.5 to E19.5, no major changes in insulin gene expression were observed (Figure 3A). Two days after birth (P2), the levels of insulin mRNA were about 10 times the levels observed at E19.5 (Figure 3A). By E13.5, low levels of Nkx 6.1, Pdx-1, and Gtx mRNAs were detected (Figures 3B and 3C). By E14.5 the levels of Pdx-1 and Nkx 6.1 mRNAs increased and then remained relatively constant until E16.5. During this period, the levels of Pdx-1 and Nkx 6.1 mRNAs were four to five times higher than those of Gtx (Figure 3C). By E17.5 the levels of all three mRNAs fell and then rose again by E18.5. Postnatally, significant expression of Nkx 6.1 and Pdx-1 was present in the pancreas but fell to very low levels in adult pancreas. Insulin expression followed a similar pattern, reflecting the development of the exocrine portion of the pancreas.
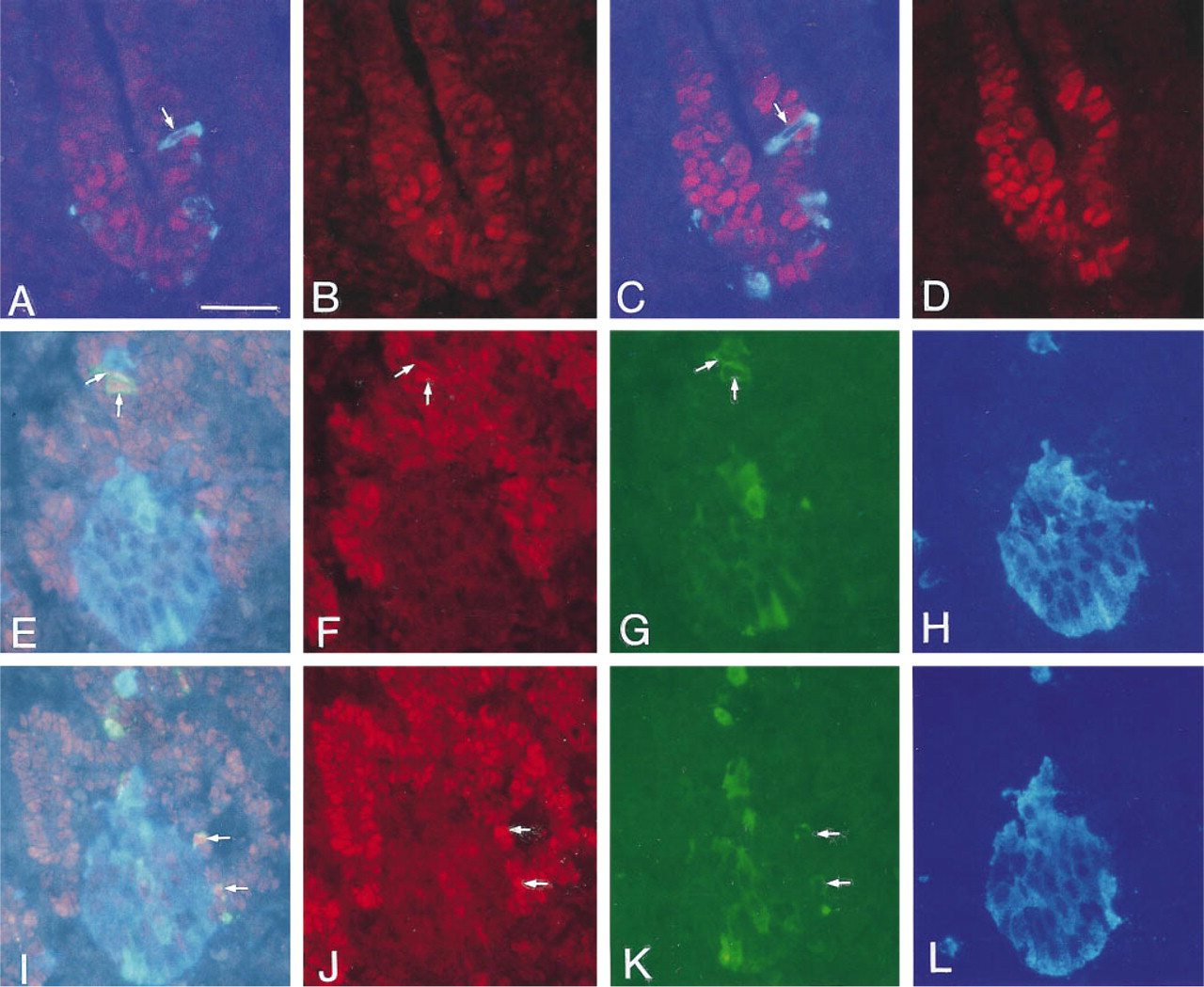
Sections through the dorsal pancreatic bud of a 22-somite rat embryo (E11.0) (
In isolated islets from new-born rats, high levels of Pdx-1 and Nkx 6.1 mRNAs were detected, whereas Gtx mRNA was undetactable.
Developing and Mature Somatostatin Cells Are Devoid of Nkx 6.1 but Show Labeling for Pdx-1
By E16.5, somatostatin cells were first observed. During E16.5-17.5, only a few somatostatin cells occurred and some of these cells also stored insulin (data not shown). No cells co-storing somatostatin and glucagon were detected. Pdx-1-positive nuclei were detected in some but not all somatostatin cells irrespective of whether or not they also stored insulin. No somatostatin cells containing Nkx 6.1-positive nuclei were found. By E18.5, somatostatin cells became much more numerous. At this stage, very few cells staining for both somatostatin and insulin were found. Almost all (>90%) somatostatin cells had Pdx-1-positive nuclei, whereas none had Nkx 6.1-positive nuclei. In addition, during the ensuing development no Nkx 6.1-positive somatostatin cells were seen. After E18.5, the frequency of Pdx-1-positive somatostatin cells decreased to the 10-20% seen in adult pancreas. No cells staining for both insulin and somatostatin were detected after E19.5.
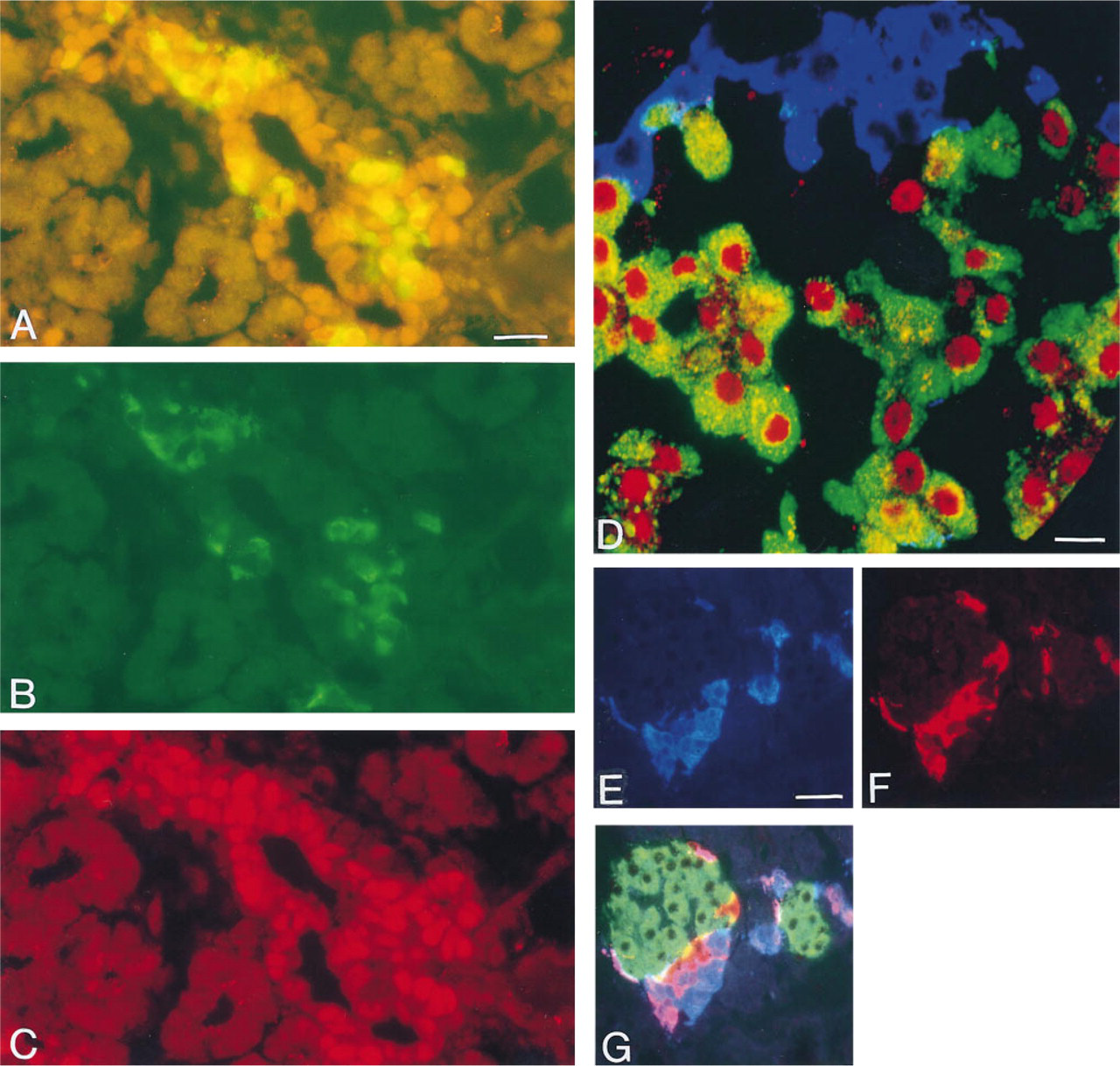
Double (
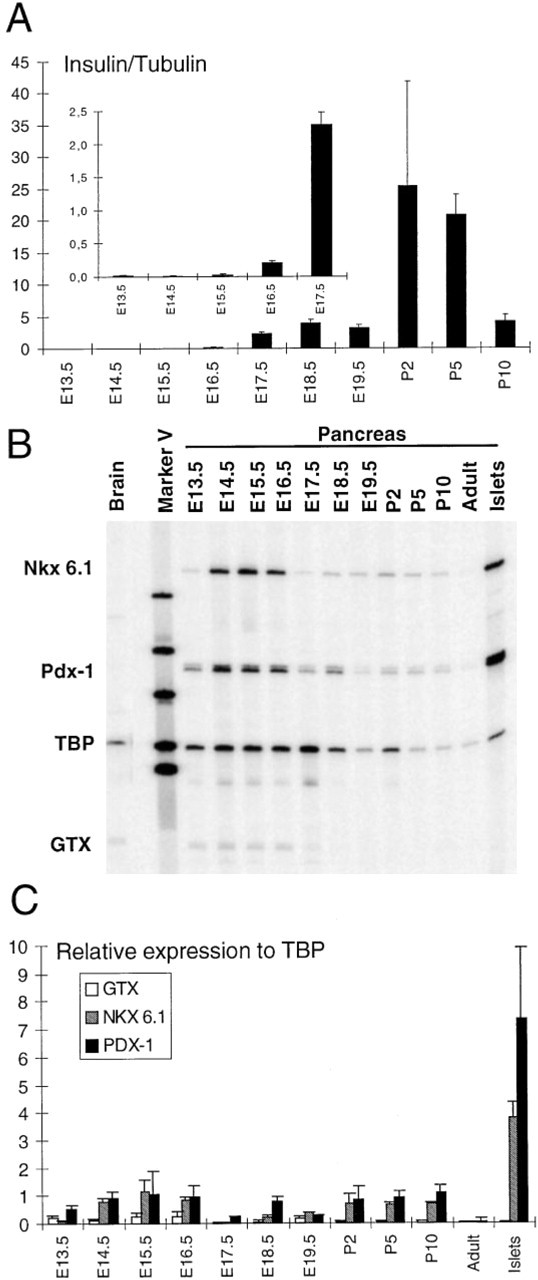
Multiplex RT-PCR analysis of gene expression during rat pancreatic development. (
Developing PP Cells Frequently Contain Glucagon and Are Occasionally Positive for Pdx-1 but Rarely Positive for Nkx 6.1
By E20.5, the first PP-immunoreactive cells were detected. Occasional cells immunopositive for both insulin and PP were seen, but after P3 no such cells could be detected. In contrast, many cells were immunopositive for both glucagon and PP (Figures 2E-2G). During E21.5-P5, counts revealed that the proportion of such double-positive cells increased from about 20-30% of all PP cells (E21.5-P2) to 60-75% at P3-P5. Subsequently, the frequency of double-positive cells decreased again. In adult pancreatic islets, variable numbers of cells immunopositive for both glucagon and PP (17-38% of PP-positive cells) were seen in the duodenal portion and tail of the pancreas. At all stages of development, weak nuclear Pdx-1 staining could be detected in a few (<5%) PP cells. Most of these Pdx-1-positive PP cells were negative for both glucagon and insulin. Only extremely rare (<1%) PP cells contained weakly Nkx 6.1-positive nuclei. These were seen only in the perinatal period and not in adults. About half of the Nkx 6.1-positive PP cells were also immunoreactive for glucagon, but none stained for insulin.
Discussion
This is the first report on the developmental expression of Nkx 6.1 in the pancreas. This homeodomain gene product was previously found to be present in the insulin cells of the adult pancreas and in RNA extracted from antropyloric mucosa of the stomach (Jensen et al. 1996). It shows sequence similarity to the glial-testis homeobox (Gtx) gene product (Komura et al. 1993), raising some concern that crossreactivity may occur. However, analyses of the expression pattern of Nkx 6.1 in developing and adult rat pancreas correlate closely to the immunocytochemical findings, whereas no such correlation is seen with Gtx. Moreover, we have recently found that Nkx 6.1-immunoreactive nuclei are absent from the antropyloric mucosa of mice deficient in Pdx-1, whereas they are readily detected in wild-type control mice. Importantly, comparable levels of Gtx gene expression were discovered in both Pdx-1-deficient and control mice (Øster et al. 1998). These data provide strong evidence that our antiserum specifically detects Nkx 6.1 and does not crossreact with Gtx. Moreover, these data show that Pdx-1 is needed for Nkx 6.1 gene expression.
Against this background, it is very interesting to note the great parallels in Nkx 6.1 and Pdx-1 gene expression in the developing pancreas. During early development the majority of the parenchymal cells were immunopositive for both Pdx-1 and Nkx 6.1, and later during development both immunoreactivities became virtually restricted to islet cells. Pdx-1 showed a wider expression pattern than Nkx 6.1 and was also detected in the duodenal epithelium and in many non-β-cells of the developing and adult pancreas. This pattern agrees with the hypothesis that Pdx-1, together with other factors, may activate Nkx 6.1 gene expression (cf. Øster et al. 1998). Pdx-1-deficient mice do not develop a pancreas (Jonsson et al. 1994; Offield et al. 1996), and it is possible that Nkx 6.1 may also be important to pancreatic development, perhaps by mediating some of the effects of Pdx-1.
We found the majority of the earliest glucagon- and insulin-immunopositive cells to be devoid of Nkx 6.1 and Pdx-1 staining. This contrasts to the report by Guz et al. (1995) that, at all stages of mouse pancreatic development, the majority of insulin cells were Pdx-1-positive, but agrees well with the data from Ahlgren et al. (1996) that the early insulin and glucagon cells also develop in Pdx-1 deficient mice.
All the insulin cells that we observed during early development co-stored glucagon. The majority of these insulin-glucagon double-positive cells were devoid of nuclear staining for Pdx-1 and Nkx 6.1 and occurred intermingled with cells positive for glucagon only. According to Pictet and Rutter (1972), cells containing β-granules are not found during this period (the “protodifferentiated state”). However, during the following days of development (the “secondary transition state”), cells with β-granules appear and increase in number. During the same period we observed a major increase in insulin mRNA and in the number of insulin-positive cells. In contrast to the early insulin cells, these “late” insulin cells were devoid of glucagon immunoreactivity, displayed strongly Pdx-1- and Nkx 6.1-immunopositive nuclei, and appeared as single cells or as small clusters closely associated with the Pdx-1- and Nkx 6.1-positive and hormone-negative epithelial cells. Coinciding with the burst in the number of insulin cells, the staining intensity for Pdx-1 and Nkx 6.1 in nuclei of hormone-negative epithelial cells declined.
It has been suggested that the insulin-glucagon double-positive cells of the developing pancreas represent precursors of mature β-cells (Alpert et al. 1988; Hashimoto et al. 1988). Recently, this theory was challenged by Pang et al. (1994), who reported that the β-cell-associated protein glucose transporter Type 2 (Glut2) protein showed a developmental distribution very similar to the distribution of Pdx-1 and Nkx 6.1 described by us. Pang et al. (1994) suggested that the early glucagon-positive insulin cells (Glut2-negative) and the later appearing single-positive insulin cells (Glut2-positive) represented two different populations of insulin cells, and that the latter were derived from Glut2-immunoreactive, hormone-negative epithelial cells. Similar patterns of distribution during development have also been described for the high-affinity NGF receptor Trk-A (Kanaka-Gantenbein et al. 1995) and fetal antigen 1 (Tornehave et al. 1996). Induction of Glut2 expression and an increase in the number of insulin cells associated with duct cells have been described after duct ligation (Hultquist et al. 1979; Wang et al. 1995). In addition, differentiation of islet cells from adult pancreatic duct epithelium in the presence of fetal mesenchyme has been demonstrated (Dudek et al. 1991). These reports are in accordance with our observations of the scattered appearance of single Pdx-1- and Nkx 6.1-immunoreactive insulin cells among the Pdx-1- and Nkx 6.1-positive hormone negative epithelial cells that we observed during E15.5-E17.5. Subsequently, these cells were found in close association with Pdx-1- and Nkx 6.1-immunoreactive duct cells (cf Figures 2A-2C). Previous quantitative studies of the developing rat pancreas indicate that preexisting islet cells could account for no more than 20% of the large increase in the number of insulin cells (>200%) observed during E20-E22 (Hellerström and Swenne 1985). Taken together, the above observations indicate that the majority of β-cells arise from hormone-negative precursors, such as Pdx-1-and Nkx 6.1-positive duct cells. However, the insulin-glucagon double-positive cells may contribute to β-cell formation as well, because these cells do indeed proliferate (Jackerott et al. 1996). Alternatively, the insulin and glucagon co-storing cells may represent a fraction of immature glucagon cells with aberrant insulin expression, like the gastrinexpressing cells observed during pancreatic development (Larsson et al. 1976a; Gittes et al. 1993).
In contrast to data obtained in the developing mouse pancreas (Guz et al. 1995), we found many glucagon-immunoreactive PP cells in the developing pancreas of the rat, especially during the perinatal period. A similar pattern has been observed in human fetal pancreas (Larsson and Hougaard 1994), in the AN glucagonoma islet cell line, and in islets from newborn rats (Jensen et al. 1996). Therefore, if islet cells develop through multihormonal precursors, these PP-glucagon double-positive cells may represent precursors of mature PP cells. In contrast to Alpert et al. (1988), but in agreement with Upchurch et al. (1994), our study revealed only rare insulin-somatostatin double-positive cells during development. This discrepancy may be explained by the thick sections used by Alpert et al. (1988) (cf. Upchurch et al. 1994). At E18.5, when a major increase in the number of somatostatin cells was observed, the majority of these cells (>90%) displayed Pdx-1-positive nuclei. These observations may indicate that somatostatin cells are derived from Pdx-1-immunoreactive precursor cells. Pdx-1 was originally cloned from somatostatin-producing islet cell lines and was characterized as a somatostatin transactivating factor (Leonard et al. 1993; Miller et al. 1994). However, Pdx-1 does not appear to be obligatory for somatostatin gene transcription, because the majority (>80%) of adult somatostatin cells are Pdx-1-negative and because Pdx-1-deficient mice show normal gastric somatostatin cell numbers (Larsson et al. 1996).
Pdx-1 has also been characterized as an insulin gene transcription factor (Ohlsson et al. 1993; Peers et al. 1994; Petersen et al. 1994; Serup et al. 1995). However, the absence of Pdx-1- and Nkx 6.1-immunoreactive nuclei in early insulin cells indicates that neither Pdx-1 nor Nkx 6.1 is obligatory for transcription of the insulin gene. In agreement with this, the presence of early insulin-positive cells in Pdx-1-deficient mice (Ahlgren et al. 1996) confirms that Pdx-1 is not necessary for insulin gene transcription. Moreover, the observation that the insulin gene, but not the Nkx 6.1 gene, is activated when Pdx-1 is transfected and expressed in glucagonoma cell lines (Serup et al. 1996; Madsen et al. 1997) confirms that Nkx 6.1 is not indispensible for insulin gene transcription. Pdx-1 and Nkx 6.1 may, however, be important for regulating insulin gene transcription and other β-cell-specific functions.
Footnotes
Acknowledgements
Supported by grants from the Danish National Research Fund (Center for Gene Regulation and Plasticity in the Neuroendocrine Network), the Danish MRC, and the Danish Biotechnology Program (Research Center for Medical Biotechnology).