
Abstract
Polyenoylphosphatidylcholine (PPC), a phosphatidylcholine-rich phospholipid extracted from soybean, has been reported to protect liver cells from alloxan-induced cytotoxicity. The present study aimed to investigate whether PPC protects pancreatic β-cells from the cytotoxic injury induced by streptozotocin, thus preserving insulin synthesis and secretion. β-Cells of the PPC-treated rats showed a significant reduction of cell death with lesser destruction of plasma membrane on streptozotocin insult. They demonstrated a rapid recovery of GLUT-2 expression, whereas almost irreversible depletion of membranebound GLUT-2 was seen in β-cells of the rats treated with only streptozotocin. A similar cytoprotective effect of PPC was also monitored in the PPC-pretreated MIN6 cells. These β-cells retained their ability to synthesize and secrete insulin and no alteration of glucose metabolism was detected. These results strongly suggest that PPC plays important roles not only in protecting β-cells against cytotoxicity but also in maintaining their insulin synthesis and secretion for normal glucose homeostasis.
I
Materials and Methods
Animals and PPC Feeding
Male Sprague-Dawley rats (Daehan Experimental Animal Lab; Seoul, Korea) weighing 150–200 g with normal fasting blood glucose levels (70–120 mg/dl) were used for PPC feeding and STZ treatment. Fifty-six rats were randomly assigned to the experimental and control groups. PPC (Rhône-Poulenc; Köln, Germany) was dissolved in drinking water and given to the rats at a daily dose of 200 mg/kg (body weight) for 3 weeks according to previous reports (Buko et al. 1994,1996). One group of rats was treated with STZ 7 days after PPC feeding. STZ (60 mg/kg body weight; Sigma, St Louis, MO) was given as a single IV injection into the jugular vein after anesthesia with inhalation of gaseous nitrous oxide-oxygen and isofrane. A second group of rats was injected with STZ without any PPC feeding, and the animals became diabetic, whereas a normal control group of rats was subjected to neither STZ nor PPC treatment. Animals were allowed free access to a normal diet (Samyang Feed; Seoul, Korea). Fasting blood glucose was monitored daily with blood samples collected from the tail vein using a glucometer (Roche; Mannheim, Germany). Three weeks after the experiment, the entire pancreatic tissue was excised from the rats after laparotomy and blood samples were collected from the heart for insulin radioimmunoassay (RIA) determination. The experimental animals were treated humanely and surgical procedures were performed according to the Animal Use and Care protocol.
Tissue Preparation
For light microscopic examination, the pancreatic tissues were fixed overnight with Bouin's solution or 4% paraformaldehyde and 5-μm-thick paraffin sections were mounted on silanized slides (Fisher Scientific; Pittsburgh, PA). For electron microscopic studies, small fragments of tissue were fixed with 4% paraformaldehyde and embedded in Unicryl resin (BBI International; Cardiff, UK).
Immunocytochemical Labeling for Light Microscopy
Immunostaining for light microscopy was carried out using the avidin-biotin-peroxidase complex (Hsu et al. 1981). Briefly, the tissue sections were incubated with the antibodies, either mouse anti-insulin (BioGenex; San Ramon, CA), rabbit anti-glucagon (Dako; Carpinteria, CA), rabbit anti-GLUT-2 (Alpha Diagnostic; San Antonio, TX), or mouse anti-PCNA (Zymed; South San Francisco, CA) and subsequently reacted with the biotinylated anti-mouse or antirabbit immunoglobulins (Vector; Burlingame, CA). After treatment with ABC (Vector), the tissues were oxidized by diaminobenzidine (Sigma; St Louis, MO) and counterstained with hematoxylin for microscopic observation. Some tissues sections fixed with 4% paraformaldehyde were subjected to TUNEL.
Immunoelectron Microscopic Labeling for Insulin
To locate insulin in the secretory granules of β-cells at the subcellular level, electron microscopic immunogold labeling was performed on ultrathin sections as described previously by Bendayan (1984). The tissue sections were incubated with guinea pig anti-insulin antibody (Dako), and the reaction was detected by the protein A-gold complex. After immunogold labeling, the grids were counterstained with uranyl acetate and lead citrate.
In Situ Hybridization
The tissues fixed with 4% paraformaldehyde were subjected to in situ hybridization as described previously (Lee and Park 2000; Kim et al. 2001). Briefly, tissue sections were incubated in the buffer containing digoxigenin-tagged oligonucleotide for insulin (2 μg/ml; R&D Systems, Minneapolis, MN) after digestion with proteinase K (5 μg/ml in Tris-buffered saline, pH 8.0). After hybridization, the sections were incubated with alkaline phosphatase-conjugated anti-digoxigenin antibody (Boehringer Mannheim; Mannheim, Germany). Hybridization signal was revealed by nitroblue tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate toluidinium.
Image Analysis for Evaluation of Immunocytochemistry and In Situ Hybridization
The β-cell number and islet mass were assessed by morphometry on tissue sections immunostained for insulin or glucagon. The entire area of each tissue section was photographed and digitalized for planimetry. The percent area of islet to pancreatic tissue was assessed using an image analyzing system (Image-Pro Plus; Media Cybernetics, Silver Spring, MD). Populations of β- and α-cells were estimated by manual calculation of insulin or glucagon positive cells in each islet. This morphometric assessment was carried out on all islets present in random sections obtained from at least five different rats of each experimental group. It is possible that the actual numbers of insulin cells are higher than the estimated numbers because only the immunoreactive cells displaying their nuclei were counted as positive cells. On the other hand, insulin content in secretory granules of the β-cell was determined in terms of the insulin-immunogold labeling index at the electron microscopic level (Bendayan 1984,1989). After determining the number of immunogold particles (Ni) and the area of each secretory granule of the beta cell (Sa), the labeling index (I) was calculated according to the formula
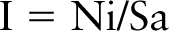
Northern Blotting Analysis for Insulin
Total cellular RNA was isolated from pancreatic tissue samples according to the protocol previously described (Chomczynski and Sacchi 1987). Total RNA was transferred to a nylon membrane (Roche) and prehybridized in hybridization buffer (7% SDS, 50% formamide, 5 X SSC, 2% blocking reagent, 50 mM sodium phosphate, pH 7.0, 0.1% N-lauroylsarcosine). Hybridization was performed with digoxigenin-labeled rat insulin cRNA, and the hybridization signal was detected with CSPD (Roche) in X-ray film.
Assessment of Insulin Secretion by RIA
Insulin levels were determined in sera and culture media. Culture media were collected each day. An insulin RIA kit was used according to the manufacturer's protocol (Linco Research; St Charles, MO).
Determination of Proliferation Activity of Islet Cells
To identify the proliferating activity of islet cells, immunostaining for PCNA was carried out on pancreatic tissues using a specific antibody (Zymed). Proliferation rate is presented as the percentage of the PCNA-labeled cells in each islet or in cell culture.
Cell Culture
We induced in vitro cytotoxic injury on a mouse β-cell line (MIN6) which was highly susceptible to STZ. The cells were grown in RPMI 1640 medium (Gibco Life Technologies; Grand Island, NY) supplemented with 10% heat-inactivated fetal bovine serum and 1% antibiotics. The cells were cultured with PPC (0.5 mM) for 1 day and then incubated with STZ (4 mM) for 2 additional days in the presence of PPC. In a second experiment, pretreatment with PPC was omitted, while both PPC and STZ treatments were excluded for control experiment. Media collected from each culture condition were subjected to RIA for insulin.
Assessment of Glucose-stimulated Insulin Secretion
The cells cultured under the above conditions were washed with PBS and then transferred to serum-free Krebs-Ringer bicarbonate buffer (KRBB; Sigma) for the test of insulin secretion responding to glucose stimulation. The cells were consequently exposed to 5 mM glucose in KRBB and 20 mM for 2 hr each, and insulin contents in KRBBs were determined by combining immunoprecipitation and western blotting (Yang et al. 1994). Briefly, the media were immunoprecipitated with the rabbit anti-insulin antibody. After denaturation they were subjected to 17% SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a nitrocellulose membrane (0.22 μm; Schleicher & Schuell, Keene, NH). Insulin was detected by chemiluminescence.
Assessment of Cell Death
The cultured cells undergoing apoptosis were determined by TUNEL assay (Sgonc et al. 1994) using a detection kit (Roche Diagnostics). Briefly, after fixation with 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) for 10 min, the cells were digested with proteinase K (1 μg/ml in 0.1 M Tris buffer, pH 7.4) and incubated with TUNEL reaction mixture containing terminal deoxynucleotidyl transferase and fluorescein-labeled nucleotides. The reaction was detected by reactions with horseradish peroxidase-conjugated sheep anti-fluorescein antibody and with diaminobenzidine. The same procedure was applied to the paraffin sections of pancreatic tissues fixed with 4% paraformaldehyde.
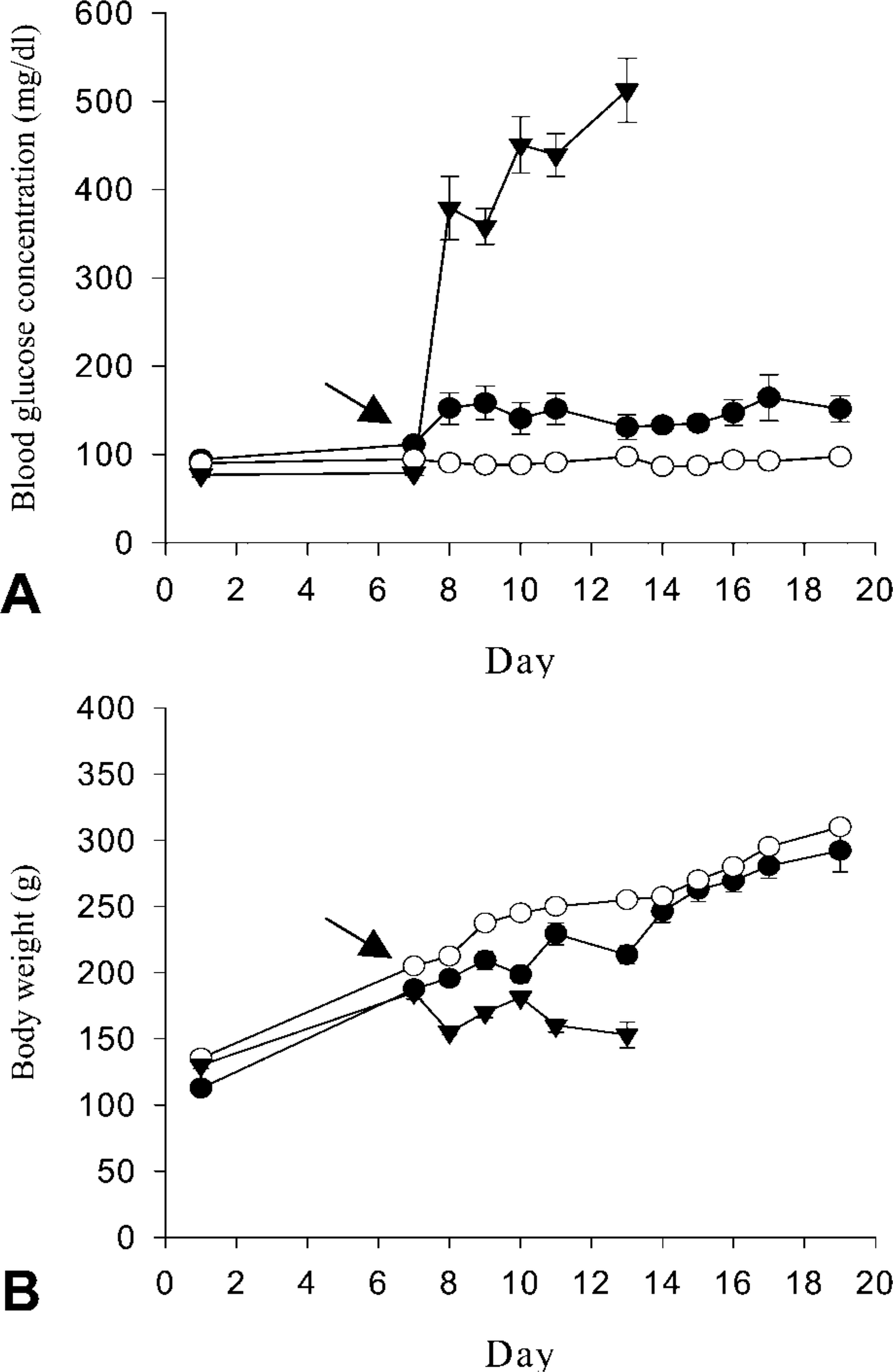
Blood glucose concentration (A) and body weight (B) of normal (open circles), diabetic rats (solid triangles), and rats fed PPC before STZ treatment (solid circles). Blood glucose levels were monitored daily at 1100–1200 hr after 3 hr of fasting. STZ (arrow) was injected on day 7 except for the controls. Data are mean values ± SE; n=12 for control, n=15 for diabetic animals, n=17 for PPC-fed rats.
Results
Modulation of Diabetes Induction in PPC-Treated Rats
The rats fed with PPC maintained normal blood glucose levels (139.28 ± 5.79 mg/dl) after STZ treatment (Figure 1A) and demonstrated normal growth and regular weight gain. However, the rats not treated with PPC but having STZ showed severe hyperglycemia (427.92 ± 27.40 mg/dl) and growth retardation with diabetic symptoms, including polyuria (Figure 1B).
Modification of GLUT-2 Expression
We examined modification of GLUT-2 expression in islet cells by immunocytochemistry (Figure 2). Because GLUT-2 is specifically localized in the plasma membrane of β-cells, it can also demonstrate alteration of membrane integrity of β-cells. GLUT-2 expression was significantly reduced by STZ intoxication in both PPC-pretreated and non-treated control rats, but the latter were more severely affected. GLUT-2 was seen in all β-cell membranes, with higher immunoreactivity in normal pancreas, but only some β-cells showed a weak immunoreaction for GLUT-2 in PPC-pretreated rats (Figure 2A and 2D). In non-pretreated control rats, moreover, no GLUT-2 was detectable in the islet cells 24 hr after STZ treatment (Figure 2B). This may be caused not only by loss of the plasma membrane of dying β-cells but also by complete impairment of GLUT-2 expression by STZ. Membrane-bound GLUT-2 was remarkably recovered after the cytotoxic insult in the PPC-pretreated rats than in those treated only with STZ (Figures 2B,2C,2E and 2F). As shown in Figure 2G, β-cells showing GLUT-2 expression in the plasma membrane were rapidly increased by 75% of the population of normal ones 3 weeks after STZ insult, while poor retrieval of GLUT-2 expression was seen in the diabetic controls at the corresponding period.
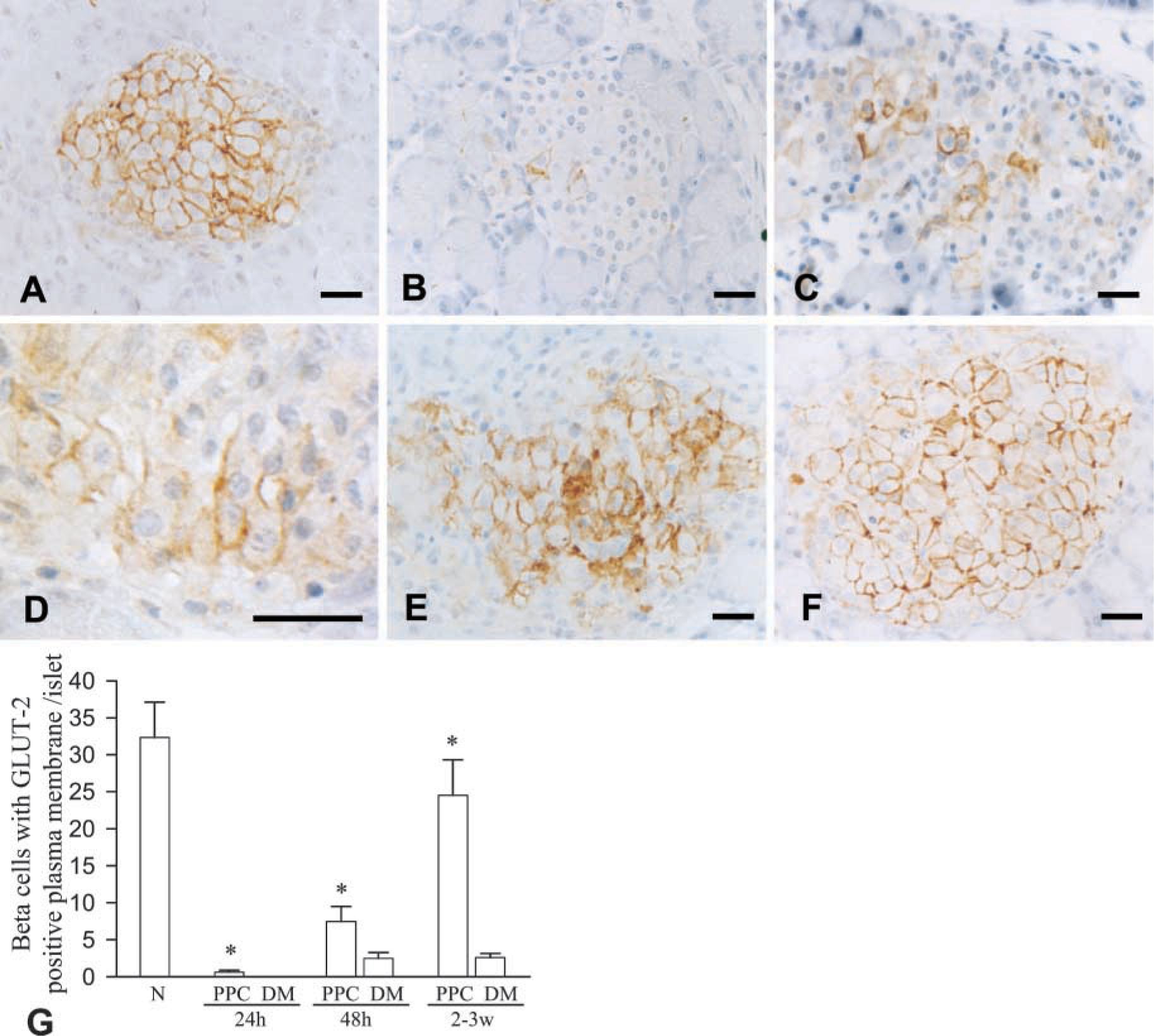
Modification of GLUT-2 expression in β-cells. GLUT-2 expression detected by immunocytochemistry demonstrates the contour of the β-cell membrane in normal controls (
Modification of Islet Cell Death and Proliferation
β-Cell death and alteration of islet cell population were prominent in the STZ diabetic rats that were not treated with PPC. β-Cell necrosis showing chromatin condensation and dissolution of plasma membrane was seen 24–48 hr after STZ injection, and some cells demonstrated apoptotic cell death (Figure 3A and 3C). β-Cell death in the core of islets led to shrinkage in islet mass, particularly at 24–48 hr after STZ treatment. However, subsequent α-cell proliferation resulted in a certain regaining of islet mass and modification of the β-cell to α-cell ratio in the islets. In contrast, such loss of β-cells and proliferation of non-β-cells were not seen in the islet cells of the PPC-fed rats in spite of the STZ treatment. Moreover, mitotic features were frequently seen in the islet cells of the rats fed PPC (Figure 3B). We examined proliferation activity of the islet cells by immunostaining for PCNA, a specific marker for proliferating cells (Figure 3D and 3E). The PCNA labeling index, the number of labeled cells in each islet (Figure 3F), was higher in PPC-fed rats (2.91 ± 0.33) than in the diabetic (0.07 ± 0.05) or control (1.17 ± 0.22) animals.
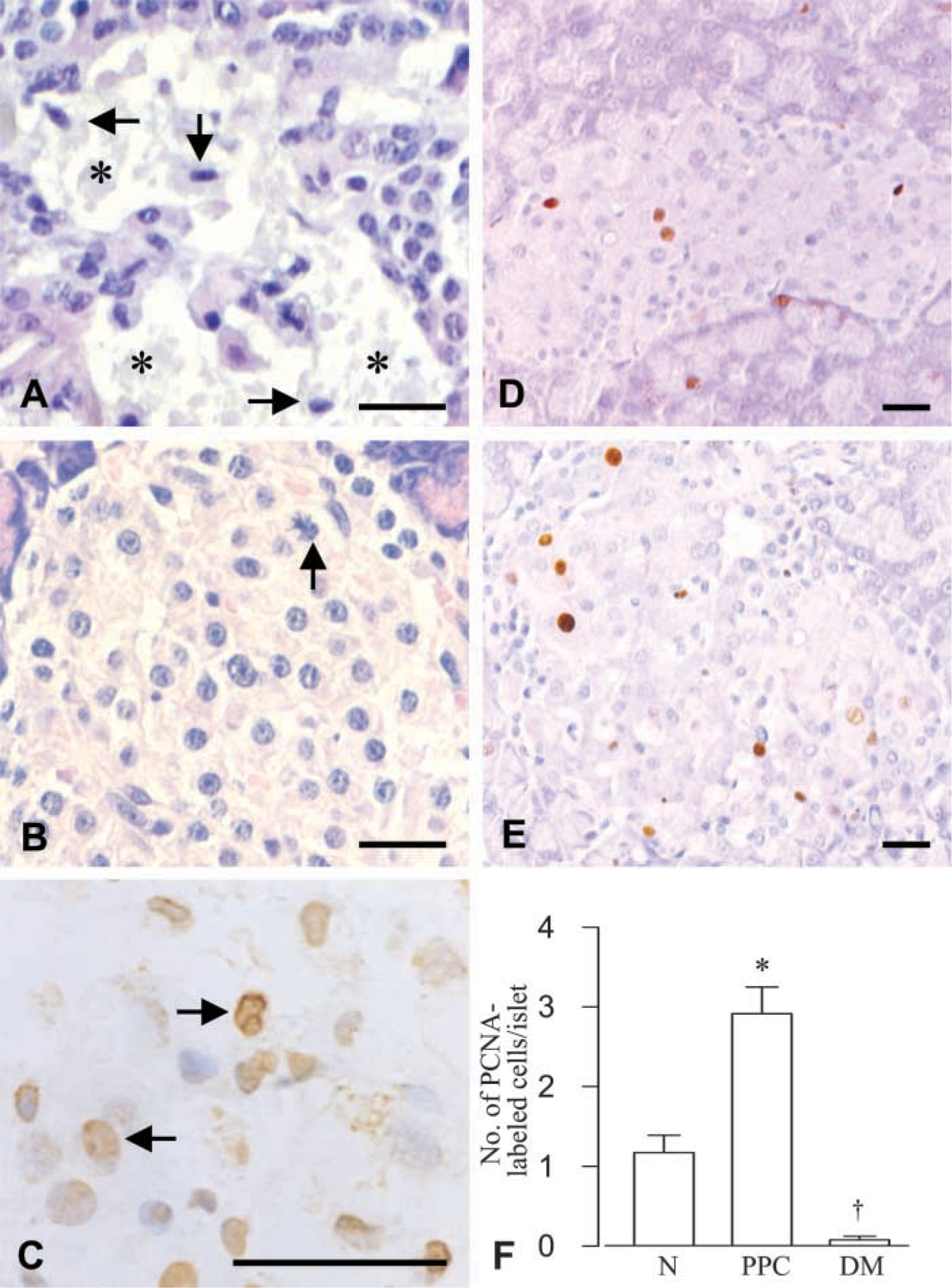
Islet cell death and replication represented by hematoxylineosin (
Morphological Alteration of Pancreatic Islets and β-cells
The islets of the PPC-treated rats retained their conventional features. They showed a normal population of β-cells centrally located, with α-cells at the periphery (Figure 4). However, diabetic rats demonstrated a remarkable increase in α-cells in the central area where only few β-cells remained, resulting in significant modifications of β- to α-cell ratio (Figures 4C-4F and 4H). At the light microscopic level, the highest insulin immunoreactivity was found in β-cells of the PPC-fed rats, whereas only few cells displayed immunoreaction with variable intensities in the diabetic animals (Figure 4B and 4C). Immunoreaction for insulin at the electron microscopic level reflects subcellular insulin content in β-cells (Bendayan 1989). The PPC-treated rats demonstrated typical profiles of β-cell ultrastructure as seen in control rats. Their secretory granules were characterized by a relatively wide halo surrounding inner dense cores, which were labeled by the insulin antibodyimmunogold complex. They demonstrated labeling densities similar to those of normal animals (Figures 5A,5B, and 5D). In contrast, poor immunolabeling was found on secretory granules of β-cells in diabetic animals. These granules were difficult to find due to the considerable degranulation of the cells (Figure 5C and 5D).
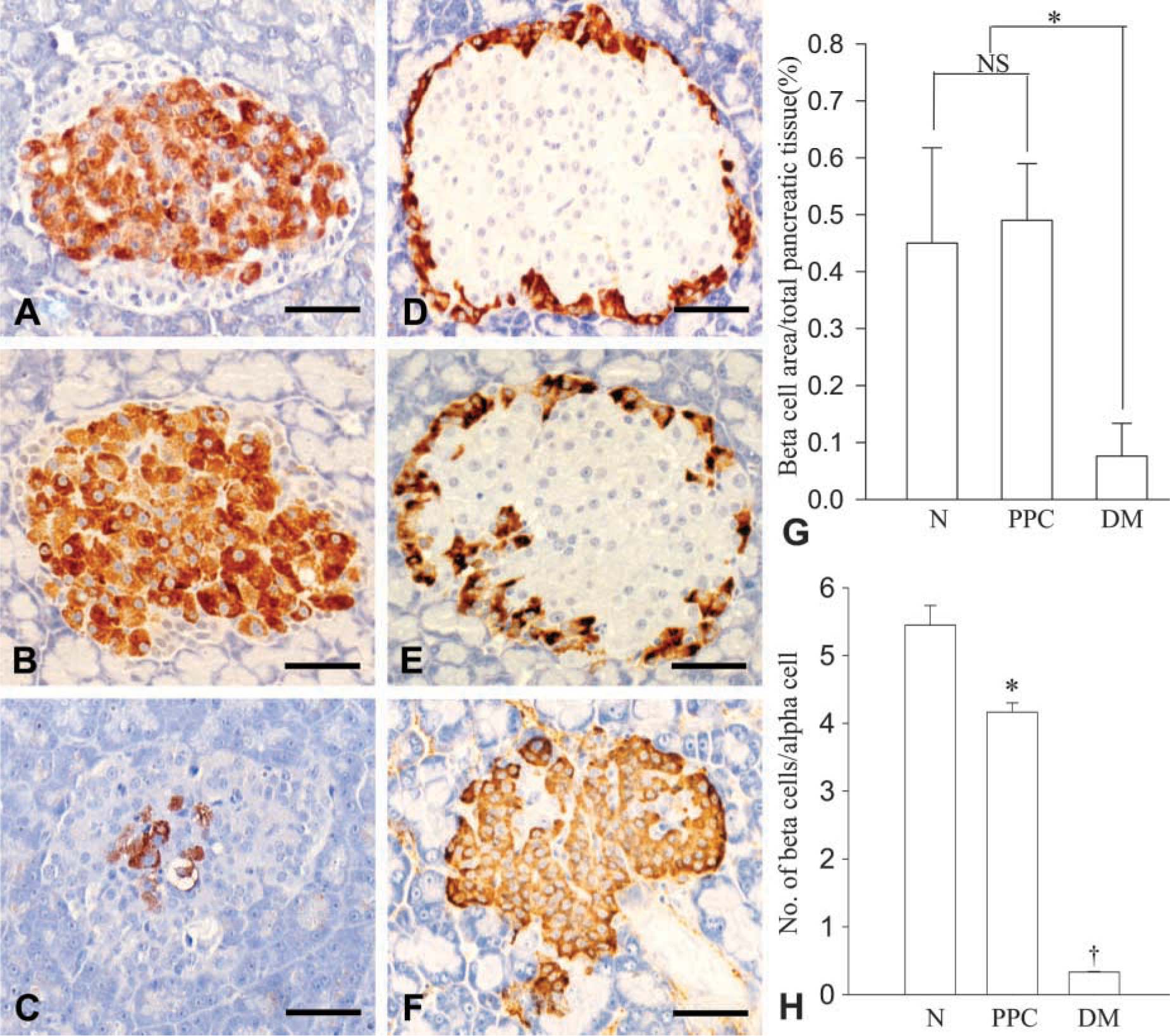
Population of insulin (
Insulin mRNA Expression
Expression of insulin mRNA, as an indication of insulin synthesis, was evaluated by in situ hybridization and Northern blotting analysis. Hybridization signals for insulin mRNA were more intense in β-cells of the PPC-treated rats than in those of the normal and diabetic animals (Figures 6A-6C). Higher expression of insulin mRNA was also detected in the pancreatic tissue of the rats fed with PPC by Northern blotting analysis, showing intense expression of insulin mRNA equivalent to that of the normal animals (Figure 6D and 6E).
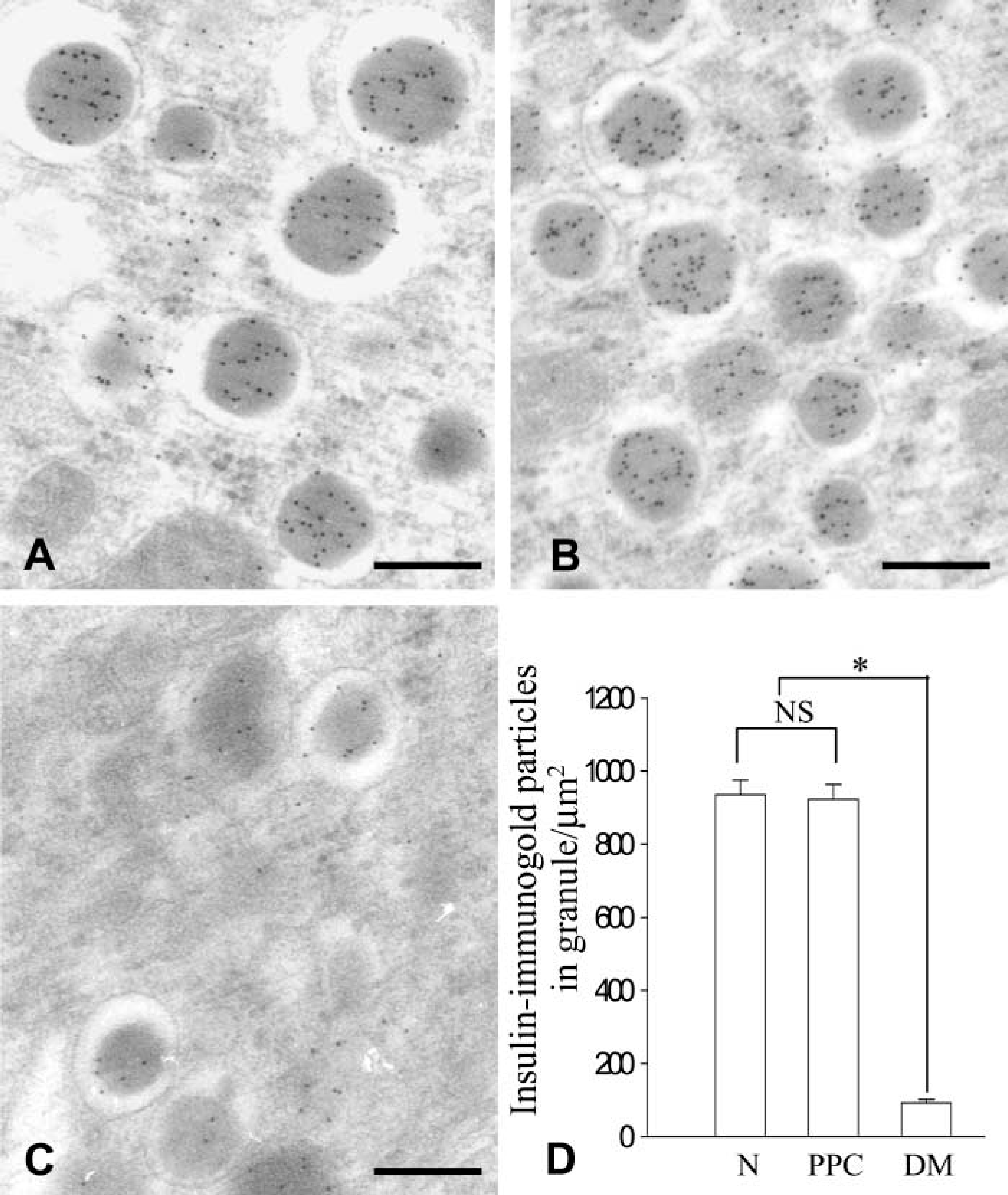
Ultrastructure and insulinimmunogold labeling of β-cells in pancreas of normal (
Modulation of Insulin Secretion and Glucose Metabolism
Fasting plasma insulin level in the rats fed with PPC (0.38 ± 0.05 ng/ml) was within the normal range, whereas that of the diabetic animals (0.05 ± 0.01 ng/ml) was significantly lower (Figure 7A). PPC-fed rats showed a normal glucose tolerance value of 75 g IPGTT, whereas the diabetic rats manifested impaired glucose tolerance (Figure 7B).
Preservation of Cell Viability and Insulin Secretion by PPC in βcell Culture
β-Cell death was examined in MIN6 cell culture by TUNEL assay. A higher rate of apoptotic cell death (7.65 ± 3.04%) was monitored after 48 hr of treatment with STZ, whereas a much lesser number of apoptotic cells was demonstrated in PPC-pretreated cells before the STZ insult (1.47 ± 0.378%) and in normal control cells (0.60 ± 0.20%) (Figures 8A-8D). Alteration of insulin secretion and glucose sensitiveness in the MIN6 cells was examined by RIA and immunoprecipitation-Western blotting analysis. Insulin concentration in culture media significantly declined after STZ treatment, whereas the media from PPC-treated cells showed no decrease in insulin concentration in spite of the STZ treatment (Figure 9A). We assessed glucose-stimulated insulin secretion in these cultured cells. As shown in Figure 9B, PPC-pretreated MIN cells produced higher levels of insulin level in response to glucose stimulation, whereas STZ-treated cells without PPC treatment displayed loss of glucose responsiveness.
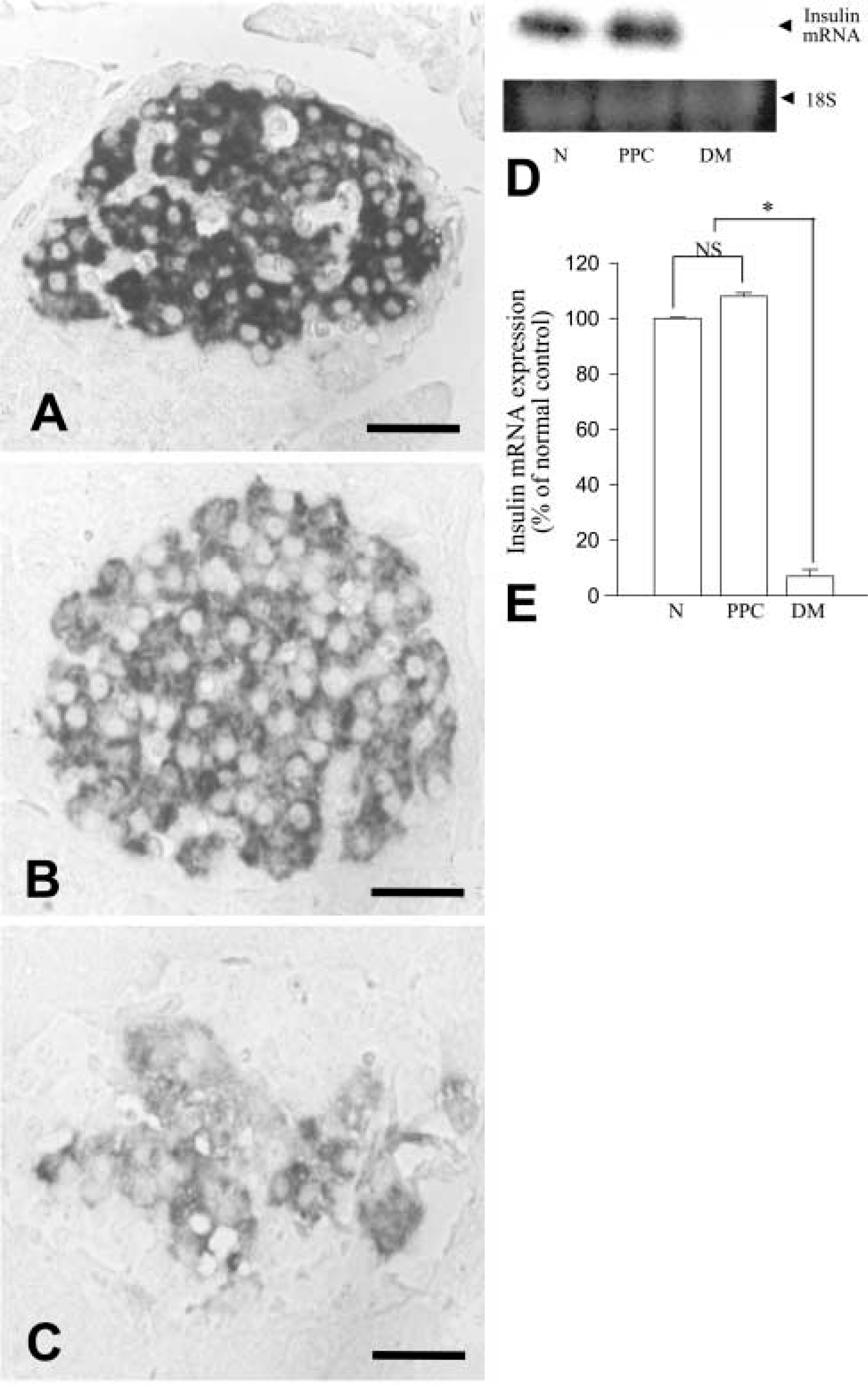
Insulin mRNA expression detected by in situ hybridization (A-C) and Northern blotting analysis (
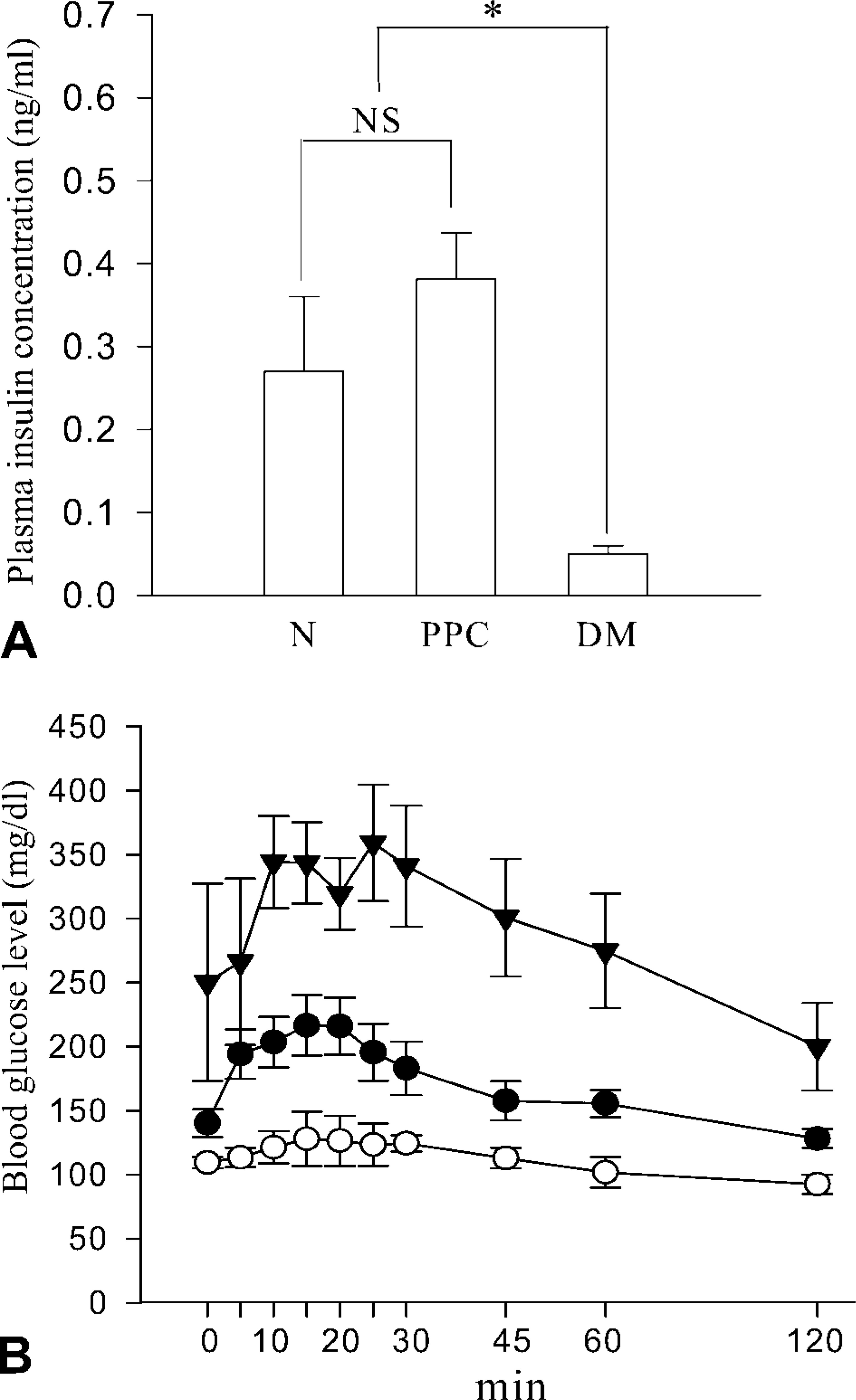
Changes in plasma insulin concentration (
Discussion
We investigated the effects of PPC as a cytoprotector of β-cells preventing development of diabetes induced by STZ. Membrane phospholipids are important regulators of cell functions such as growth, viability, homeostasis, and signal transduction (Awad et al. 1996; Wang et al. 1998; Flores et al. 2000). Modification of phospholipid composition could therefore deteriorate cell functions by altering membrane fluidity, permeability, and receptor activity (Buko et al. 1994; Saragovi et al. 1998; Alvarez et al. 2001; Valcarcel et al. 2001). STZ, a diabetogenic compound that induces death of β-cells, modifies the molecular structure of phospholipids, particularly phosphatidylcholine (PC), which is a major phospholipid of the outer leaflet of the plasma membrane, resulting in decrease of membrane fluidity in β-cells (Vecchini et al. 2000; Detmar et al. 1994; Wieder et al. 1995; Cui et al. 1996). In the present study we found that β-cell death was considerably inhibited with amelioration of membrane destruction in the PPC-treated rats, although their membrane-bound GLUT-2 expression was significantly suppressed. GLUT-2 is not only a structural protein specific for β-cell membrane but is also a crucial constituent for recognition and entry of glucose as well as the glucose analogue STZ (Noel and Newgard 1997). This indicates that suppression of β-cell death in PPC-treated animals is not associated with the facility of STZ entry by GLUT-2 in the β-cell membrane. PPC-treated rats, moreover, exhibited an increased proliferation activity with a rapid retrieval of membranebound GLUT-2 in the islet cells after STZ insult. PPC is a polyunsaturated phosphatidylcholine bearing two linoleic acids in its molecule. Dietary PPC can provide unsaturated fatty acid-rich phospholipids to the β-cell membrane, thus facilitating reconstruction of membrane integrity as reported in rat liver tissue (Buko et al. 1996). It is therefore likely that PPC facilitates retrieval membrane constituents of β-cells as well as their cell replicability.
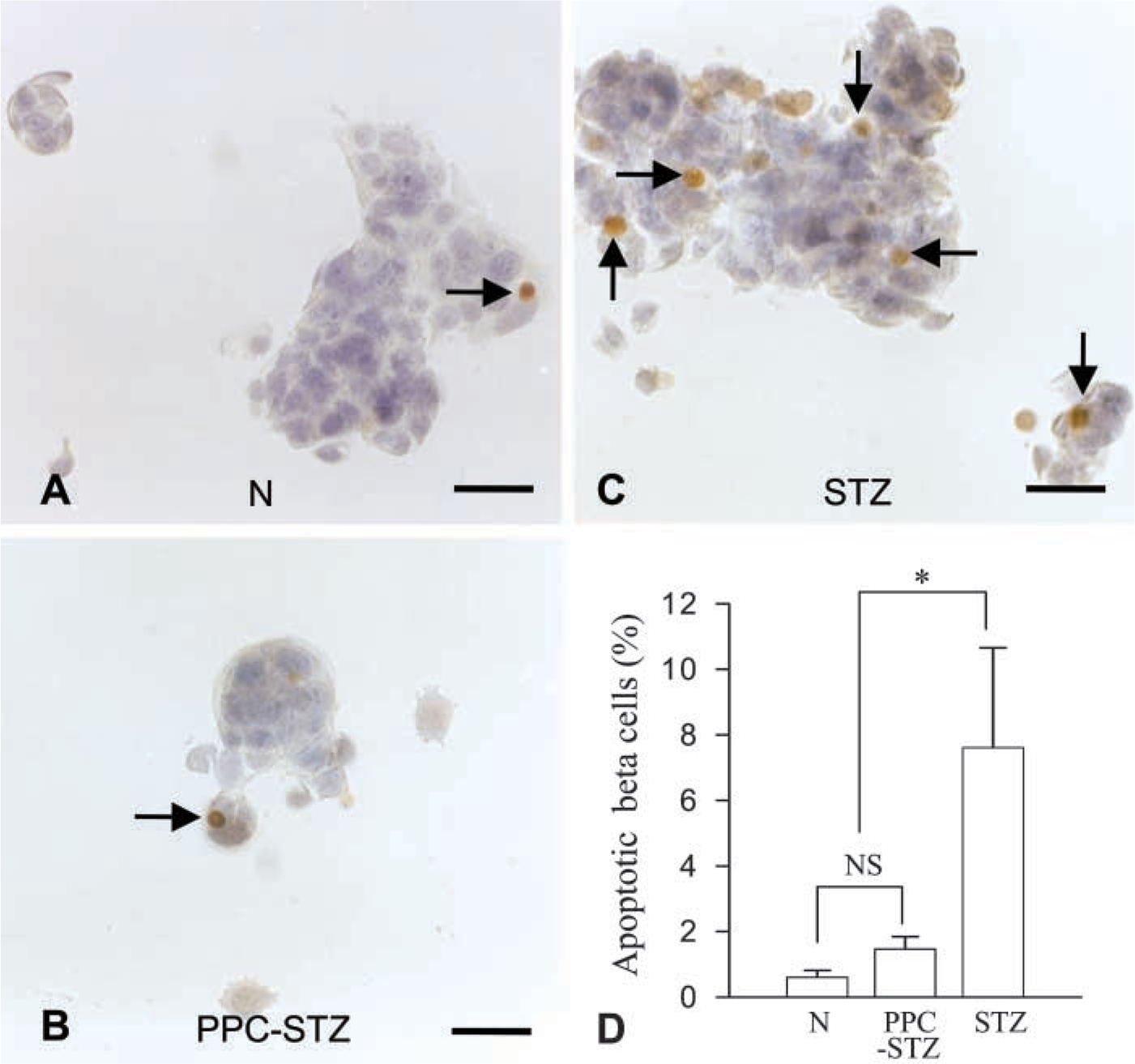
In-vitro detection of cell death by TUNEL assay. The nuclei of apoptotic cells (brown nuclei indicated by arrows in A-C) were seen in MIN6 cells cultured with PPC before STZ treatment (PPC-STZ) or STZ treatment only (STZ). The cells for normal controls (
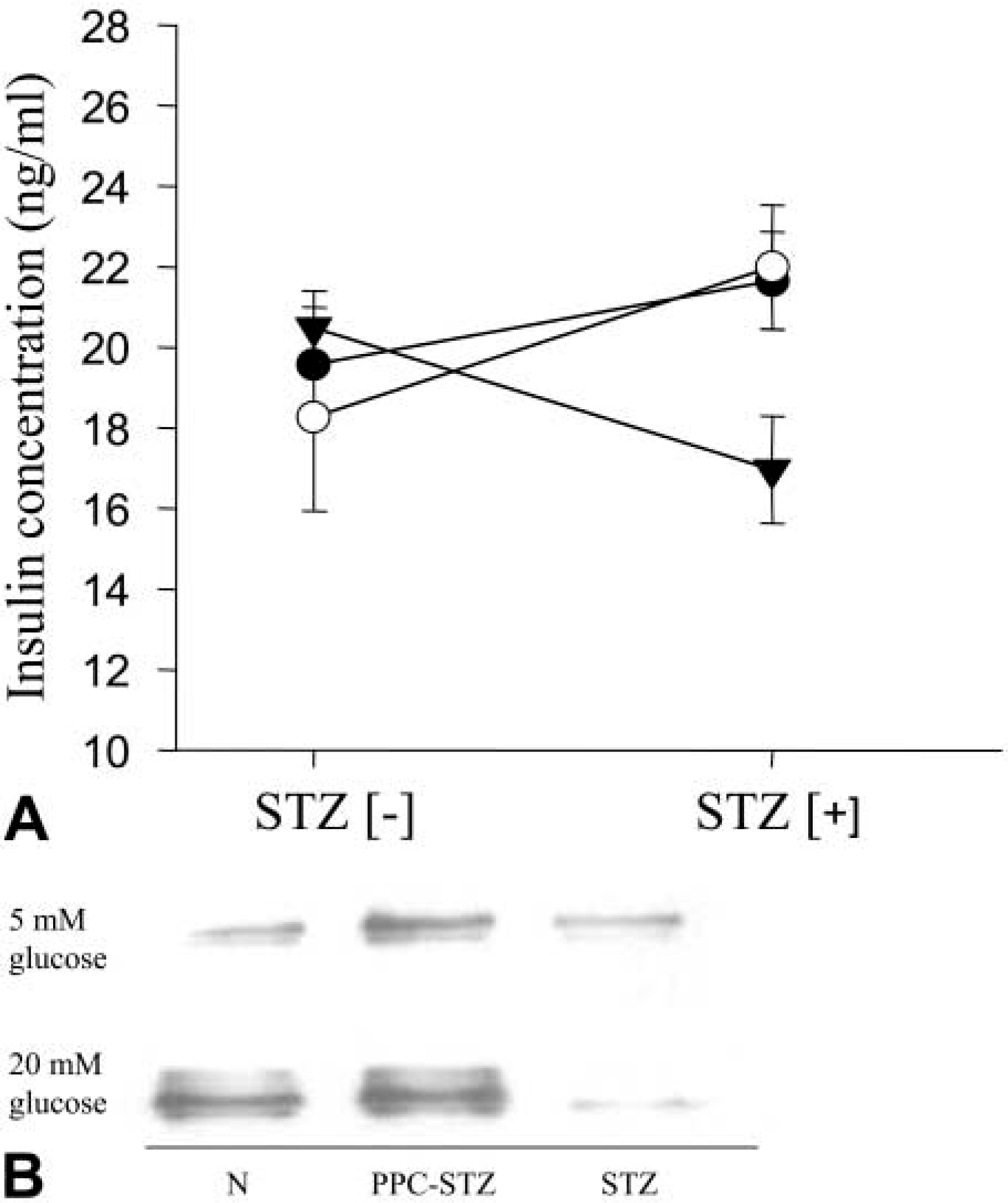
Insulin concentrations of culture media from different conditions. Media were collected after 24 hr of culture without STZ treatment (STZ [-]) and 24 hr of culture in the presence of STZ (STZ [+]). There is a significant decrease in insulin concentration after STZ treatment in the culture without PPC treatment (solid triangles), whereas no reduction was noted in both the PPC-pretreated culture (open circles) and the untreated control (solid circles). Data in A are mean values ± SE (n=3 independent experiments). Glucose-stimulated insulin secretion was determined in these cultured cells by combining immunoprecipitation and Western blotting (
The β-cells, along with preservation of their population in PPC-treated rats, did not manifest impaired insulin secretion. They displayed well-preserved insulin immunoreactivity and insulin mRNA expression as well as glucose-stimulated insulin secretion, indicating an almost intact process for insulin synthesis and secretion. In fact, the secreted insulin in bloodstream or in culture media was within the normal range in PPC-treated groups.
We suggest that a sufficient supply of phospholipid with unsaturated fatty acids should be considered not only to improve β-cell function but also to suppress the diabetogenic process in diabetes-prone subjects.
Footnotes
Acknowledgements
Supported by a grant from the Oriental Medicine R&D Project, Ministry or Health and Welfare, Republic of Korea (HMP-00-CO-06-0006).
We wish to thank Dr V. U. Buko (Institute of Biochemistry, National Academy of Science of Belarus) for kind discussion and suggestions.