
Abstract
The generation of a functional nervous system is dependent on precise path-finding of axons during development. This pathfinding is directed by the distribution of local and long-range guidance cues, the latter of which are believed to be distributed in gradients. Gradients of guidance cues have been associated with growth cone function for over a hundred years. However, little is known about the mechanisms used by growth cones to respond to these gradients, in part owing to the lack of identifiable gradients in vivo. In the developing grasshopper limb, two gradients of the semaphorin Sema-2a are necessary for correct neuronal pathfinding in vivo. The gradients are found in regions where growth cones make critical steering decisions. Observations of different growth cone behaviors associated with these gradients have provided some insights into how growth cones respond to them. Growth cones appear to respond more faithfully to changes in concentration, rather than absolute levels, of Sema-2a expression, whereas the absolute levels may regulate growth cone size.
T
The growth cone is a highly dynamic, actin-rich structure capable of recognizing and responding to a variety of guidance cues. It uses these cues to elicit a number of stereotyped behaviors, including outgrowth, turning, fasciculation, retraction, and stalling (Suter and Forscher 2000). In situations where guidance cues are distributed as concentration gradients, the growth cone is believed to discern small changes in external concentrations across its spatial extent (Goodhill and Urbach 1999). The growth cone then translates these changes into intracellular signals, leading to a reorganization of the actin and microtubule cytoskeleton and thus to the correct pathfinding behavior (Goodhill and Urbach 1999). Two mechanisms have been put forth to explain how growth cones distinguish these subtle changes in concentration. A major difference between the two models lies in whether a growth cone responds to either the absolute or the fractional change in concentration across its area (Walter et al. 1990; Goodhill 1998; Goodhill and Baier 1998). Although many attempts have been made to resolve this issue using elegant in vitro preparations, there has been little effort to apply these models to in vivo gradients. We have recently identified gradients of a guidance cue in the grasshopper limb bud that are important for neuronal guidance (Isbister et al. in press). Careful immunocytochemical examination of these gradients has enabled us to characterize the nature of the gradients and their effects on growth cone behavior and function. This review discusses some of these observations and their importance for growth cone guidance mechanisms in vivo.
The Ti1 Pioneer Pathway
The developing grasshopper limb bud serves as a model system in which to examine growth cone dynamics and guidance in response to a gradient of ligand in vivo. This system is suitable for investigating many aspects of axon guidance, including the distribution and role of guidance cues and mechanisms used by growth cones in response to them. In addition, the relatively large size of grasshopper growth cones allows direct observations of their behavior when labeled with lipophillic dyes such as DiI or DiO (O'Connor et al. 1990; Isbister and O'Connor 1999).
During development of the grasshopper, a pair of afferent neurons arises in the distal tip of each limb bud. These tibial (Ti1) pioneer neurons extend axons along a stereotyped pathway on route to the CNS (see Bentley and O'Connor 1992). At 30% of embryogenesis, a single mother cell arises from the distal limb tip epithelium to give rise to the pair of Ti1 neurons (Bate 1976; Bentley and O'Connor 1992). These neurons extend axons proximally along the limb epithelium and establish a stereotyped pathway to the CNS that is completed 24 hr later at 35% of development (Figure 1). The pathway is characterized by a number of critical growth cone steering decisions and, in general, the growth cones exhibit stereotyped behaviors during these steering decisions (O'Connor et al. 1990; Sabry et al. 1991; Isbister et al. 1999).
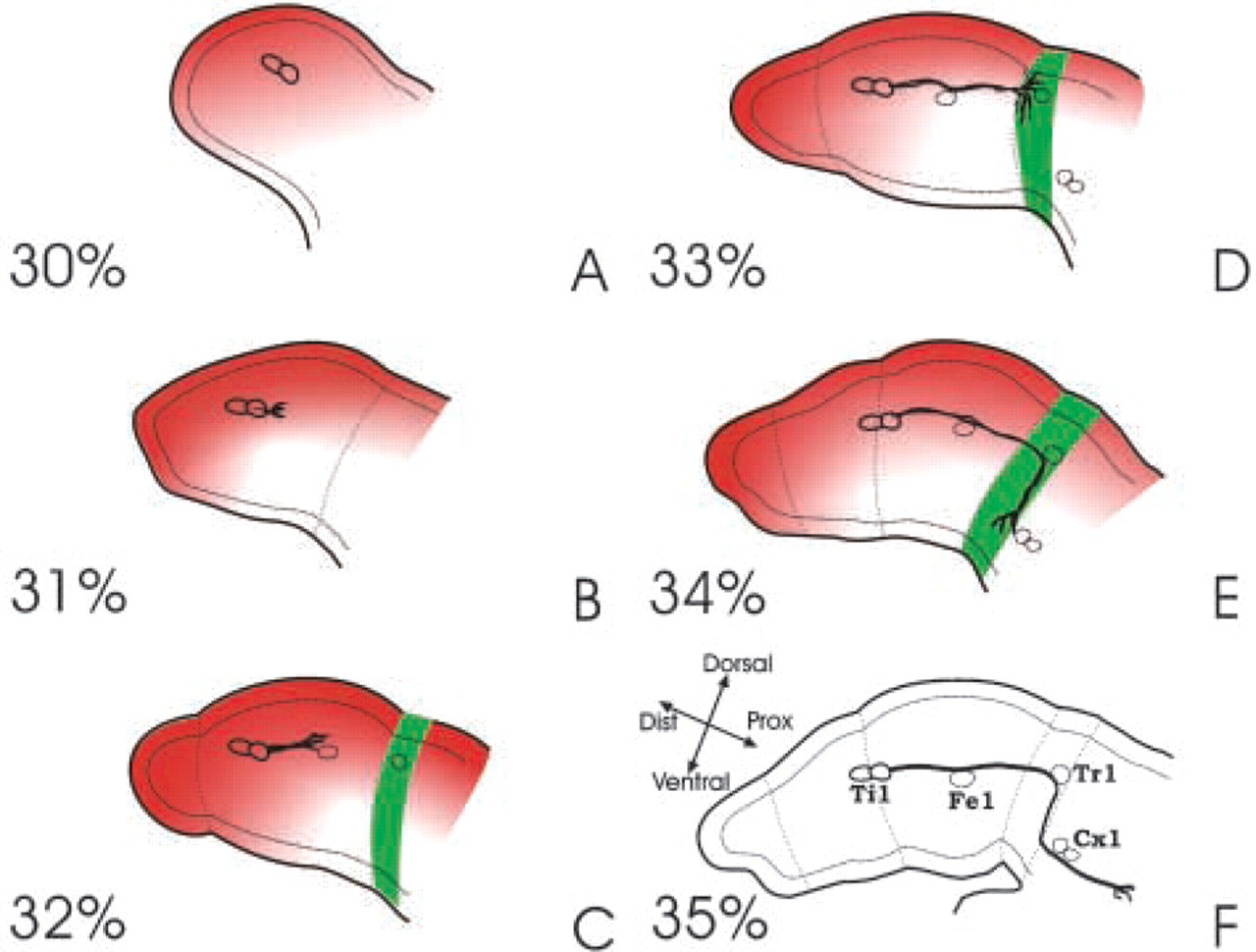
Schematic of the Ti1 pioneer neuron pathway and semaphorin distribution from 30% to 35% of development. The repulsive semaphorin 2a (red) is expressed as a gradient as early 30% of development (
The Ti1 neurons act as pioneers, laying down a neuronal path to which later developing axons fasciculate and follow. As the Ti1 growth cones extend, they migrate between the basal surface of the epithelial cells and the basal lamina (Bate 1976; Keshishian and Bentley 1983; Lefcort and Bentley 1989). Growth cones encounter a number of different guidance stimuli along the way, including guidepost cells (neuronal precursor cells derived from the epithelium that act as intermediate targets for the growth cone), mesodermal cells (located within the limb lumen and along the basal lamina), the extracellular matrix, including the basal lamina, and the limb epithelium. The first steering decision that the Ti1 neurons make is to extend their axons proximally in the limb. This decision is made quite reliably (see below), with even the distal Ti1 neuron quickly reorienting its axons on the few occasions that it initiates a distal axon. After the initial proximal extension, the Ti growth cones continue to grow proximally until they contact a preaxonogenesis neuron, the Tr cell, located in the trochanter segment of the limb (Figures 1D and 1F). On contact with the Tr cell, the growth cones pause and then make a second steering decision to turn ventrally along the trochanter epithelium before contacting another intermediary target, the Cx cells, located near the ventral midline of the embryo (Figure 1E). After Cx contact, the neurons make a proximal turn and continue to the CNS (Figure 1F). The distribution of guidance cues plays an important role in each of these steering decisions. The first two decisions, in particular, appear to be mediated entirely or in part by the graded distribution of a repulsive guidance cue, Sema-2a, and disrupting the function of this cue leads to a number of guidance errors at these precise steering regions (see below).
Semaphorin Expression Along the Ti1 Pioneer Pathway
The semaphorins are a family of guidance cues that are important for the development of the nervous system. Two members of the semaphorin family are expressed in the developing limb at the stage of Ti1 outgrowth and have been shown to play a role in guiding the Ti1 axons (Kolodkin et al. 1992; Wong et al. 1997; Isbister et al. 1999). With more than 20 different family members identified to date, the semaphorins comprise one of the largest families of guidance molecules. They can be subdivided into at least eight different classes on the basis of domain structure and species of origin (Semaphorin Nomenclature Committee 1999). Semaphorins comprise a diverse family of secreted and membrane-associated proteins characterized by the presence of a conserved semaphorin (Sema) domain, 500 amino acids in size. The first identified semaphorin, grasshopper Sema-1a (previously named fasciclin IV), was initially described and implicated in axon guidance by the use of antibody perturbation experiments (Kolodkin et al. 1992). Subsequent purification of a secreted semaphorin, chick collapsin-1 (Sema3A), demonstrated the chemorepulsive nature of this semaphorin as acute addition led to rapid actin depolymerization and growth cone collapse of DRG neurons in vitro (Luo et al. 1993). After this initial characterization, a number of studies demonstrated the repulsive nature of semaphorins for a number of neuron populations (Kolodkin et al. 1993; Matthes et al. 1995; Tanelian et al. 1997; Bagnard et al. 1998; Chedotal et al. 1998; Shoji et al. 1998; Winberg et al. 1998a,b; de Castro et al. 1999). Some semaphorins appear to affect pathfinding decisions by regulating axon fasciculation and target selection (Matthes et al. 1995; Winberg et al. 1998a,b; Yu et al. 1998; Cloutier et al. 2002). In addition to functioning as repulsive guidance cues, semaphorins also function as attractive cues. For example, whereas Sema3A is repulsive for cortical axons, another semaphorin, Sema3C, acts as an attractive guidance cue for the same neurons (Bagnard et al. 1998). Similarly, in the developing grasshopper limb, the Ti1 neurons turn away from their normal pathway to contact cells ectopically expressing Sema-1a, suggesting an attractive response (Wong et al. 1999). In addition, the activation of the cGMP pathway in Xenopus spinal neurons can convert Sema3A repulsion to attraction (Song et al. 1998). Additional examples of both repulsive and attractive roles for these proteins in vitro and in vivo have been demonstrated (see Raper 2000).
In the grasshopper, a band of epithelial cells corresponding to the trochanter segment of the limb express Sema-1a uniformly (Figure 1). This band of Sema-1a expression corresponds with a proximal to ventral reorientation of the Ti1 neurons and provides a pathway for neurons to migrate along until reaching the Cx1 guidepost cells (Figure 1). In the presence of antibodies that block Sema-1a function, the Ti axons pathfind normally until they reach the trochanter, where they defasciculate and branch extensively, often extending into the coxa (Kolodkin et al. 1992) (Figure 2A). However, despite their altered morphology the neurons still turn ventrally. The secreted semaphorin Sema-2a is expressed distally at the tip of the limb bud and dorsally along the length of the limb (Isbister et al. 1999) (Figure 1). The initial proximal extension of the Ti1 neurons then occurs in a region of high Sema-2a expression in the distal limb bud. Once these extending axons approach the band of Sema-1a expression at the trochanter, the growth cones pause and extend a number of filopodia into the surrounding environment. The neurons then turn ventrally away from the dorsal Sema-2a expression and begin to migrate down the band of Sema-1a-expressing cells. Blocking Sema-2a function with antibodies raised against the semaphorin domain leads to guidance errors that include distal projections in the distal limb tip, along with dorsal projections into regions normally expressing high levels of Sema-2a (Isbister et al. 1999) (Figures 2B and 2C). Therefore, high distal and dorsal levels of Sema-2a prevent axons from venturing into these regions, suggesting a repulsive function for Sema-2a. In contrast, blocking Sema-1a function causes growth cones to wander off of the Sema-1a-expressing cells (Kolodkin et al. 1992), suggesting an attractive or permissive function for Sema-1a in the developing grasshopper. Culturing embryos in the presence of Sema-1a and Sema-2a antibodies combined leads to a combination of errors characteristic of both Sema-2a and Sema-1a blocking (Isbister et al. 1999). Thus, these two proteins play a role in correctly guiding neurons in the grasshopper limb bud.
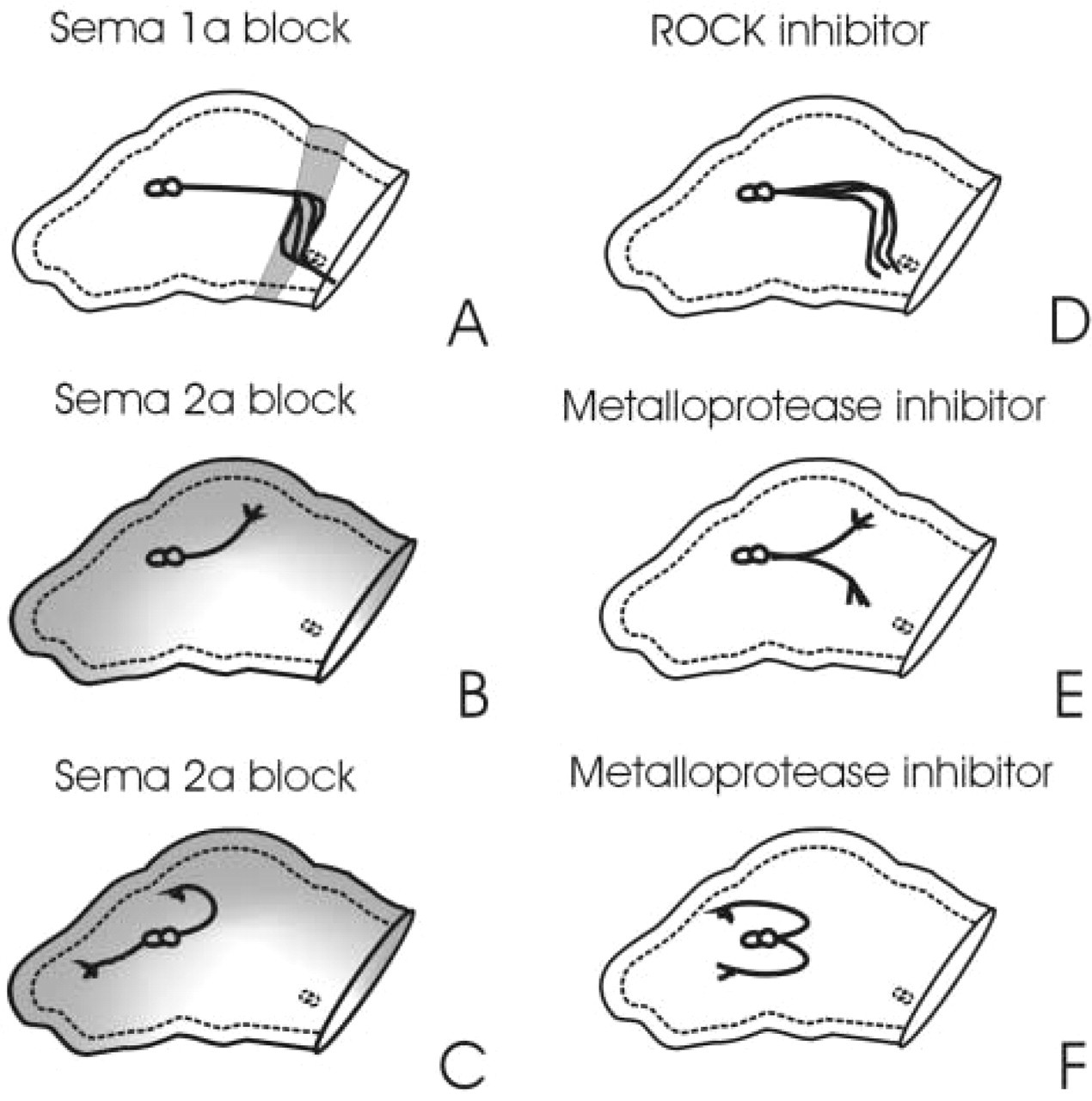
Schematic illustration comparing errors from semaphorin blocking experiments (
Two Exponential Gradients of Sema-2a
Detailed analysis of Sema-2a distribution demonstrates that it is expressed as two overlapping, yet perpendicular gradients in the developing limb (Isbister et al. 1999; Isbister et al. in press). Immunocytochemical analysis with antibodies against Sema-2a clearly indicates that the distal-proximal expression gradient encloses the Ti1 cell bodies. Curve fit analysis of the gradient demonstrates it is described by two separate equations, an exponential gradient from the distal limb to a point proximal to the Ti cell bodies, followed by a proximal linear gradient (Isbister et al. in press). A second exponentially distributed gradient of Sema-2a occurs along the dorsal-ventral axis within the trochanter, but with a significantly shallower slope (Isbister et al. 1999). These two gradients converge near the Tr guidepost cell, which consequently coincides with a critical decision point in the pathway when the Ti1 neurons reorient themselves ventrally. Despite the different steepness of the slopes of the gradients, they do not differ significantly in their peak magnitude (Isbister et al. in press).
When axonogenesis initiates at approximately 30% of development, the Ti1 growth cones interact with the steeper distal-proximal gradient of Sema-2a (Figures 1A and 1B). The two cell bodies can differ in their initial direction of growth cone projection. The proximal Ti1 cell body ordinarily projects its axon from the proximal pole, whereas the distal Ti1 cell body may initiate its axon from either the distal or proximal pole (Lefcort and Bentley 1989). Growth cones emerging from the distal pole immediately reorient themselves and extend proximally. Axons then continue to extend down the gradient of Sema-2a toward the trochanter. A second reorientation of the growth cones occurs at approximately 33% development when they reach the trochanter segment and reorient themselves ventrally to extend down the shallower dorsal-ventral gradient of Sema-2a (Figures 1D and 1E). Although growth cones orient themselves down the repulsive gradient of Sema-2a in both instances, analysis of growth cone behaviors in the two gradients reveals striking differences. In the distal limb bud, growth cones do not sample much of their environment before migrating proximally. In contrast, at the trochanter, the growth cones pause and typically extend processes into much of the surrounding environment, including dorsally up the gradient of Sema-2a expression. In addition, growth cones often extend branches dorsally up the Sema-2a gradient before extending branches in the correct ventral direction. Studies from other systems suggest that growth cones undergo changes in morphology and behavior, depending on their surroundings and location along their developmental pathway. In tracts and pathways, growth cones are smaller and relatively simpler in nature but in decision regions where they change direction and approach their targets, growth cones have a more complex morphology (Caudy and Bentley 1986; Holt 1989; Norris and Kalil 1991,1992). This phenomenon occurs at the trochanter when Ti1 neurons make a ventral turn. As neurons extend proximally before the ventral turn, axons travel rapidly down the gradient and are associated with smaller growth cones (Isbister et al. in press). On contacting the Tr cell and epithelial expression of Sema-1a, growth cone morphology changes, leading to an increase in spatial sampling and branch formation in the incorrect direction. Do differences in the nature of Sema-2a gradients help to facilitate these behavioral changes?
Pathfinding Errors Associated with Sema-2a Gradients and Ectopic Expression
On comparison of the two gradients, we found that the slope of the distal-proximal exponential gradient in the distal limb was consistently significantly steeper than that of the dorsal-ventral gradient at the trochanter (Isbister et al. in press). We hypothesized that the steepness of the gradient might confer important guidance information, explaining the differences in growth cone behaviors at the two choice points (Isbister et al. in press). To examine this hypothesis, we analyzed the frequencies of errors at both choice points in normally developing embryos. Normally, in a wild-type population of grasshopper embryos, approximately 10–15% of the Ti1 pathways display errors (Kolodkin et al. 1992; Isbister et al. 1999). When untreated embryos are analyzed for errors at 36% of development, after the Ti1 pathway is completed, we found that the overwhelming majority of errors occurred during growth cone interactions with the shallower dorsal-ventral gradient (Isbister et al. in press). These errors were typified by an extension of axons into dorsal regions at the trochanter (Figure 3A). The few errors associated with the distal-proximal gradient were typically an extension of the axon distally into the limb bud or a lack of a single axon (Figure 3B). Because fewer errors occurred in the steeper distal gradient, we hypothesized that the steepness of the gradient prevented the guidance errors. To ensure that differences in errors were not due to sampling at a stage well beyond the initiation of axon outgrowth (30–31%), we examined growth cone error rate at approximately 32.5%. Similar to the above results, we found that very few errors were observed in the distal limb bud where the Ti1 neurons initiate axonogenesis, and that the majority of errors observed were of axons extending into the dorsal limb compartment. Therefore, in both cases the Ti1 neurons made significantly more errors along the Sema-2a gradient with the shallower slope. Surprisingly, for the neurons to make dorsal errors at the trochanter they had to extend axons into regions of Sema-2a expression that were similar in concentration to Sema-2a expression in the distal limb, further confirming that the shape of the gradient, rather than the absolute concentration, is the important feature for accurate neuron growth (see below).
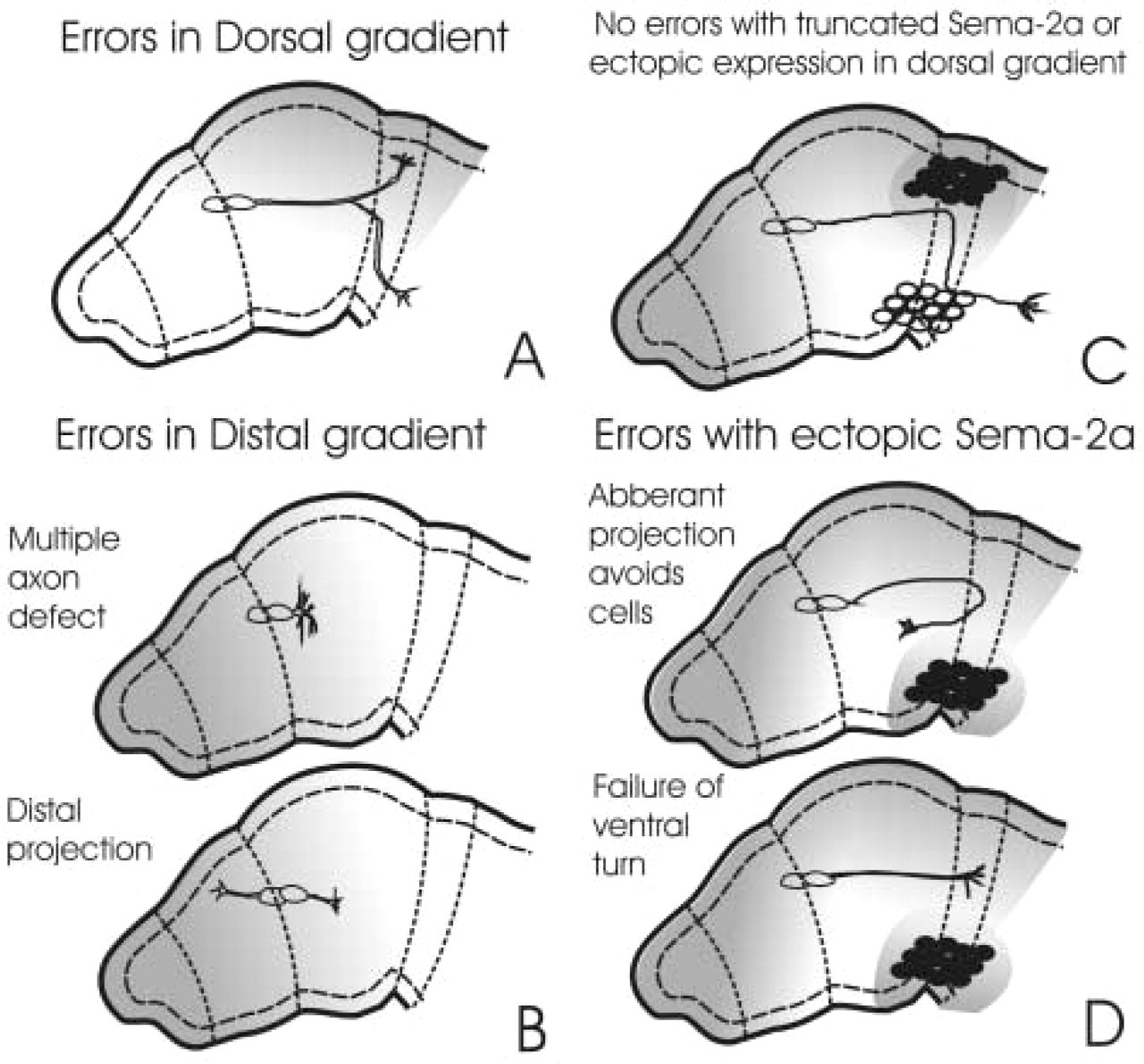
Schematic representation of the errors associated with the two gradients of Sema-2a in the wild-type and antibody-blocked limbs. (
To confirm that these steering decisions were the result of the Sema-2a gradients rather than other guidance cues, we examined the frequencies of errors made by Ti1 growth cones in the presence of Sema-2a function-blocking antibodies (Isbister et al. 1999). If alternative cues direct correct Ti1 axon initiation and/or correct ventral turning, then antibodies to Sema-2a should have little effect on the steering decisions made in these locations. Alternatively, if the Sema-2a gradients are important for these steering decisions at these locations, then an increase in errors should occur in each region and it would be expected that there would be no bias in the frequency of errors. In the presence of the antibodies we observed a significant increase in the total number of errors and found that the frequencies of errors in the distal limb and trochanter were the same (Isbister et al. in press). These results show that, when the function of Sema-2a is blocked, the Ti1 growth cones are as equally likely to err in the steep gradient as in the shallow, supporting the notion that the Sema-2a gradients are important for correct steering decisions at these locations.
To further substantiate that Sema-2a played a central role in guiding Ti1 axons, Sema-2a-expressing S2 cells were transplanted into the ventral compartment of developing limbs (Isbister et al. in press). When Sema-2a-expressing cells were transplanted into the ventral compartment of the limb at 32–33% of development, the Ti1 axons frequently did not make a correct ventral turn (Figure 3D). This often resulted in axons not turning ventrally or reorienting their axons distally or dorsally to avoid the Sema-2a-expressing cells (Figure 3D). In contrast, placing cells expressing Sema-2a in dorsal regions of the limb had no effect on Ti1 pathfinding, suggesting that the increased levels of Sema-2a did not affect Ti1 outgrowth per se, yet disrupting the dorsal to ventral gradient does. Placing control S2 cells expressing truncated Sema-2a in any regions within the limb had no effect on Ti neuron pathfinding. These observations further support the importance of Sema-2a as a guidance cue required for proper Ti1 pathfinding.
The Shape of the Gradient, Not the Absolute Concentration, Confers Guidance Information to the Ti1 Growth Cones
As described above, the Ti1 growth cones err more frequently when turning ventrally in the trochanter (where a shallow gradient of Sema-2a is found) compared to when they initiate proximal growth in the limb (where a steeper gradient of Sema-2a is found). One explanation for these observations is that growth cones in the distal-proximal gradient are exposed to a higher absolute concentration of Sema-2a and therefore make fewer steering errors. However, when examining the total amounts of Sema-2a encountered by Ti1 growth cones that extend aberrantly into the dorsal trochanter, we found that the levels of Sema-2a were the same (Isbister et al. in press). Therefore, the absolute levels of Sema-2a do not appear to be responsible for the better fidelity of pathfinding in the distal limb. However, growth cone morphology differed markedly in these two regions and, on further analysis, it was found that the absolute concentrations of Sema-2a may regulate growth cone size (Isbister et al. in press). That is, axons in the higher concentration distal gradient have smaller growth cones in comparison to growth cones in regions of lower Sema-2a. It therefore appears that the concentration of chemorepellent may be more important in determining the morphology and, more specifically, the size of the growth cone than in directing a specific steering response.
For the Ti1 growth cones to acquire guidance information from a gradient, they must be able to detect subtle differences of concentration across the growth cones. Two models that predict how growth cones detect small changes in substrate concentration differ in whether growth cones utilize the absolute or the fractional change in concentration across their extent (Walter et al. 1990; Goodhill 1998; Goodhill and Baier 1998). The absolute model predicts that the growth cone detects a change in the absolute concentration of a ligand across its spatial extent, whereas the fractional change model predicts that the growth cone measures changes in the concentration as a fraction of the average concentration detected by growth cone. In the limb, absolute changes in concentration of Sema-2a protein across the growth cones are not significantly different between the two gradients, as the differences in the size of the growth cones at each decision point result in similar absolute changes across the growth cone (Isbister et al. in press). In contrast, there is a significant difference in the fractional changes across the growth cones at these points in the pathway (Isbister et al. in press). These results suggest that growth cones measure the relative changes in protein concentration across the growth cone rather than absolute changes. Although the models would predict this behavior of growth cones in an exponential gradient, a demonstration of this in vivo has not been shown before (Goodhill and Urbach 1999).
Precedence for the importance of gradient shape and steepness has come from a number of elegant studies in vitro. Studies on retinal ganglion neurons from embryonic chick demonstrate that growth cones from the temporal half of the retina are sensitive to small concentration changes of the guidance cues across their spatial extent (Baier and Bonhoeffer 1992). The results show that retinal growth cone response to a repulsive cue varied depending on the steepness of the gradient in a stripe assay. In a separate study, cortical axon growth cones responded to the direction and sign of in vitro gradients of Sema3A and Sema3C (Bagnard et al. 2000). Interestingly, cortical axon length was independent of the absolute concentration or steepness of the gradient. In a contrasting study, Rosentreter et al. (1998) demonstrated that retinal axons extend further up shallower gradients of tectal membrane repellent to a specific point of avoidance. This point of avoidance does not correlate with a fixed absolute concentration but rather with a similar increment of concentration relative to the basal level. Unfortunately, these studies did not address the effects of gradients on growth cone turning, but rather focused on the length and rate of outgrowth in a gradient in vitro.
Cytoskeletal Aspects of Turning
Alhough it is clear that Ti1 growth cones sense subtle changes in concentrations of guidance cues, it is not clear how the growth cones modify their cytoskeleton in response to these cues. Initial studies on growth cone behavior and morphology implicate the dynamic actin and microtubule cytoskeleton in underlying growth cone motility and guidance (Bentley and Toroian-Raymond 1986; Letourneau et al. 1987; Forscher and Smith 1988). For the guidance information supplied by the Sema-2a gradient to be effective, the growth cones must be able to signal to the underlying cytoskeleton. If this is indeed the case, then an analysis of actin dynamics in the context of a gradient of Sema-2a must demonstrate changes in the underlying cytoskeleton that reflect the nature of the gradient. Observations of filopodia from Ti1 growth cones in the process of turning at the trochanter reveal that individual filopodia travel up to threefold faster in the ventral direction compared to dorsally projecting filopodia (Isbister and O'Connor 1999). This phenomenon holds true even when ventrally extending filopodia from one Ti1 growth cone overlap with dorsally extending filopodia from the other growth cone. Even though both filopodia in this overlapping region are exposed to the same guidance information, they differ in direction of outgrowth. It is quite possible that filopodia traveling dorsally respond to the shallow dorsal gradient and slow outgrowth as they travel up the concentration gradient, whereas filopodia traveling in the ventral direction down the gradient of Sema-2a would continue to grow without slowing in response to Sema-2a. Indeed, this biased reorganization of the actin cytoskeleton towards the correct ventral target correlates well with the gradient of Sema-2a at the trochanter, suggesting that the information supplied by the gradient is translated into the proper cytoskeletal response. Interestingly, these observations also suggest that perhaps even individual filopodia are capable of sensing concentration differences. This observation is consistent with studies on Xenopus spinal neurons demonstrating that individual filopodia generate local bursts of calcium influx in response to a variety of guidance cues (Gomez et al. 2001). These bursts of calcium influx propagate back to the growth cone, leading to a global increase in calcium levels, a slowing of the individual filopodia extension rate, and turning when preferentially stimulated on one side of the growth cone (Gomez et al. 2001).
Signaling
How the cytoskeleton responds to extracellular guidance cues is important for understanding the mechanisms of axon guidance, but there is still the question of what links these two processes together. Many recent studies have sought to identify the signaling molecules used to transmit the information provided by guidance cues to the cytoskeleton. Recent results have lead to a complicated catalogue of mechanisms underlying growth cone motility and dynamics. These include such signaling events as localized protein synthesis (Campbell and Holt 2001), calcium influxes (Gomez et al. 2001), proteolytic activity (Galko and Tessier-Lavigne 2000), and a number of secondary signaling molecules, including cyclic nucleotides (Ming et al. 1997; Song et al. 1998), the Rho family of small GTPases (Grunwald and Klein 2002), and phospholipids (Ming et al. 1999). Because most of these studies are conducted in an in vitro environment, the role and significance of process in directing growth cone behavior in vivo is unclear. The grasshopper provides an excellent model in which to study these processes in vivo. Recent results from our lab have demonstrated that inhibition of both the Rho family of small GTPases and the proteolytic activity conferred by metalloproteases is important for correct Ti1 pathfinding.
The Rho Family of Small GTPases
Initial research focusing on Rho GTPases implicated these proteins as important players in outgrowth and retraction of axons (Jalink et al. 1994; Luo et al. 1994). Rho is thought to mediate axon retraction, whereas Rac1 and Cdc42 have been implicated in neurite extension (Hu et al. 2001). In neuronal cell lines, activation of Rho leads to axon retraction, whereas blocking Rho function prevents retraction induced by an external stimulus (Jalink et al. 1994). Rho GTPases also appear to be important for neuronal growth and pathfinding in vivo. For example, in Drosophila, over-expression of dominant negative Rac causes guidance errors in motor axons at specific choice points in vivo (Kaufmann et al. 1998). Recent studies have shown that cofilin/actin depolymerizing factor, a protein with actin-depolymerizing activity, is inactivated during Rho-mediated axon retraction (Maekawa et al. 1999). Evidence from a number of studies strongly implicates Rac in semaphorin signaling (Rohm et al. 2000; Vikis et al. 2000; Driessens et al. 2001; Hu et al. 2001). In addition to Rac activity, recent work by Aurandt et al. (2002) has implicated the activation of Rho by a Rho-specific nucleotide exchange factor LARG, in response to stimulation by another semaphorin, Sema4D. Therefore, many studies have demonstrated the importance of the Rho family of small GTPases during axon guidance and, more particularly, as downstream molecules of semaphorin signaling (Dickson 2001; Patel and Van Vactor 2002).
To determine whether the Rho family of small GTPases is important for proper axon pathfinding in the grasshopper, the activity of a downstream Rho kinase (ROCK) was inhibited in vivo. Grasshopper embryos cultured in the presence of a ROCK inhibitor displayed a significant number of Ti1 pathfinding errors (Figure 2D). Surprisingly, the aberrant projections were confined to regions of Sema-1a expression and appeared to phenocopy the anti-Sema-1a antibody blocking experiments, suggesting a role for Rho in Sema-1a signaling (Figure 2D). In contrast, inhibition of ROCK activity had little effect on a growth cone response to Sema-2a gradients.
Metalloprotease Activity in the Limb
In addition to the intracellular signaling in response to guidance cues, there is evidence that proteolytic activity associated with growth cones may be important for proper migration (Krystosek and Seeds 1981a,b). In Drosophila, flies containing mutations in the kuzbanian gene display a number of stalled axon guidance defects (Fambrough et al. 1996). The kuzbanian gene encodes a member of the ADAM (a disintegrin and metalloprotease) class of metalloproteases. Additional studies have demonstrated that metalloprotease activity influences netrin activity, because inhibition of metalloprotease activity potentiates netrin-mediated axon outgrowth in embryonic rat dorsal spinal cord explants (Galko and Tessier-Lavigne 2000). The receptor for netrin, DCC, is a substrate for metalloprotease activity and may require this proteolytic modification for proper recycling. Previous studies have implicated proteolytic processing for the proper activity of some semaphorins, including the secreted semaphorin Sema3A and the transmembrane semaphorin Sema4D (Adams et al. 1997; Elhabazi et al. 2001). To test whether metalloprotease activity is important for proper Ti1 neuron pathfinding in the grasshopper limb, embryos were cultured in medium with an inhibitor of metalloprotease activity. Inhibition of metalloproteases resulted in a number of guidance errors (see Figure 2). The majority of errors were characterized by defasciculation of the Ti1 axons, with one axon extending dorsally and one extending ventrally (Figure 2E). Other guidance errors included the distal turning of Ti1 neurons and premature stalls (Figure 2F). The rate of guidance errors increased significantly as the concentration of inhibitor increased. Unlike the guidance errors associated with Rho signaling inhibition that are similar to Sema-1a antibody-blocking defects, the errors from these experiments phenocopied the errors associated with Sema-2a antibody-blocking experiments (Figures 2B and 2C). These results suggest that proper signaling through Sema-2a may require proteolytic processing, although where this occurs, at the level of Sema-2a or its receptors, is unclear.
Summary
A central question of developmental neurobiology is how growth cones are able to interpret changes in the external concentration of guidance cues and to transfer this information internally to alter their direction and correctly steer in vivo. Addressing this question has been hampered, in part, by the lack of guidance gradients described in vivo. In the grasshopper limb bud a known guidance molecule, Sema-2a, is expressed in a gradient that is amenable to analysis and perturbation. Based on the results from studying the grasshopper Ti1 pathway, the nature of the distribution of guidance cues appears crucial for correct path-finding. Indeed, growth cones appear to detect fractional changes across their extent, with steeper slopes conferring greater repulsive information and therefore fewer guidance errors. How these guidance cues signal to the cytoskeleton is still not well understood. However, preliminary results suggest that specific semaphorins may signal down different pathways. This may suggest that the different semaphorins function through distinct receptor systems, a hypothesis now being tested.