
Abstract
Recent studies have suggested that the soluble N-ethylmaleimide-sensitive factor attached protein (SNAP) receptor (SNARE)-mediated membrane fusion system is involved in vesicle fusion with the surface plasma membrane, which leads to neurite elongation. There have been several reports analyzing the effects of neurite outgrowth by inhibition of SNAREs. We studied this mechanism by overexpressing GFP-fusion SNAREs including VAMP-2, SNAP-25A, and syntaxin1A in PC12 cells to investigate the role of SNAREs in neurite outgrowth. When overexpressed in PC12 cells, VAMP-2 promoted neurite elongation, whereas SNAP-25A stimulated neurite sprouting. On the other hand, overexpression of syntaxin1A neither promoted nor inhibited neurite outgrowth. Thus, VAMP-2 and SNAP-25A play different roles in neurite elongation and sprouting.
D
On the other hand, the SNARE hypothesis (Söllner et al. 1993b) has been proposed to explain the vesicle fusion mechanism at the synapse. The SNARE system consists of proteins on the vesicular membrane, called v-SNARE, and proteins on the target membrane, called t-SNARE. VAMP (Trimble et al. 1988) is a v-SNARE protein, whereas SNAP-25 (Oyler et al. 1989) and syntaxin (Bennett et al. 1992) are t-SNARE proteins. These three proteins form a complex with proteins of the general fusion machinery (Söllner et al. 1993a).
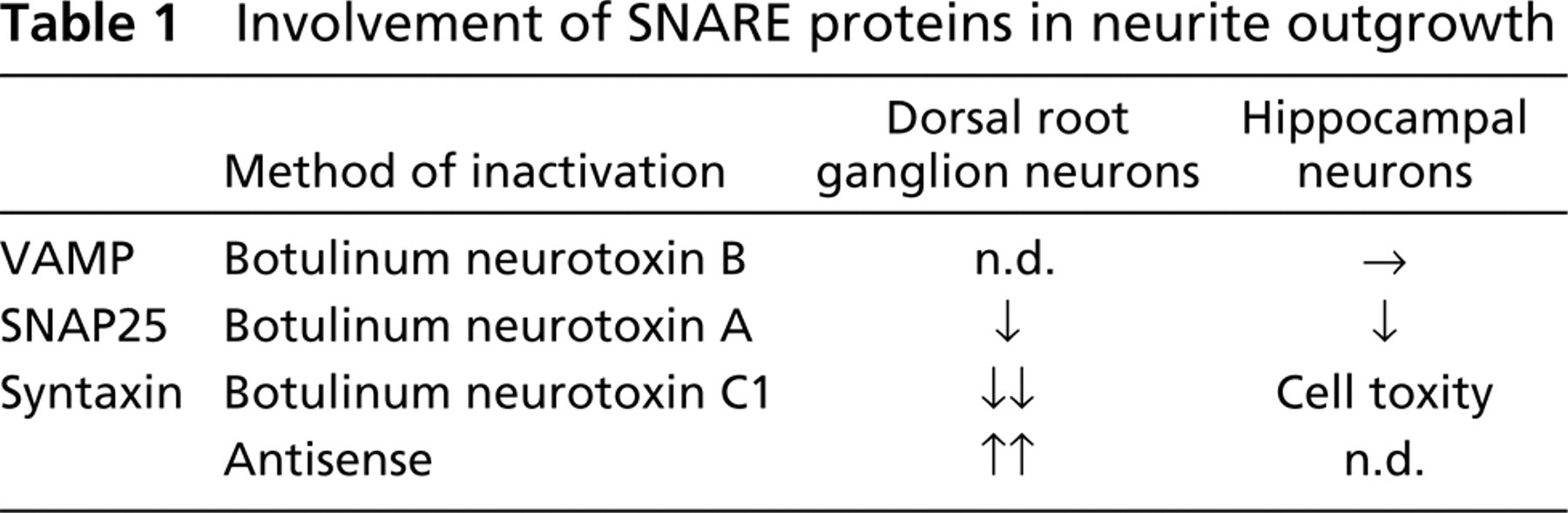
The SNARE system has been suggested to be involved in neurite outgrowth (Table 1) (Hepp and Langley 2001; Tang 2001). VAMP-2 (Elferink et al. 1989) was shown to participate in neurotransmitter release, although it had no effect on neurite extension when cleaved with botulinum neurotoxin B or tetanus neurotoxin (Ahnert-Hilger et al. 1996; Osen-Sand et al. 1996). Inhibition of SNAP-25 expression by antisense oligonucleotides prevented neurite elongation in rat cortical neurons and PC12 cells in vitro (Osen-Sand et al. 1993). Cleavage of SNAP-25 with botulinum neurotoxin A inhibited axon growth (Osen-Sand et al. 1996; Morihara et al. 1999). In contrast, inhibition of syntaxin1A expression or activity with antisense oligonucleotides or antibodies increased neurite sprouting and neurite length of both rat dorsal root ganglion neurons and retinal ganglion neurons (Yamaguchi et al. 1996). On the other hand, cleavage of syntaxin by botulinum neurotoxin C1 inhibited axon growth (Igarashi et al. 1996). Furthermore, the possible cytotoxic effect of neurotoxin C1 cannot be overlooked. Other reports also indicated that the SNARE mechanism operating in the growth cone is involved in membrane expansion for axon growth (Igarashi et al. 1997).
Involvement of SNARE proteins in neurite outgrowth
Several analyses using inhibition of SNAREs were performed as described above, indicating that SNAREs have an important role in membrane expansion for neurite outgrowth. However, there are contradictory reports concerning the effect of syntaxin inhibition on neurite outgrowth described above. Furthermore, a cytotoxic effect when toxins are used must be taken into consideration. Here we approached the role of SNARE proteins for neurite outgrowth by overexpressing green fluorescent protein (GFP)-fusion SNAREs (VAMP-2, SNAP-25A, syntaxin1A) in PC12 cells.
PC12 cells were seeded in dishes and transfected with plasmids expressing GFP-SNAREs, after which the medium was replaced with a growth medium containing nerve growth factor. These exogenous GFP-SNAREs are known to form a complex with endogenous SNAREs in transfected PC12 cells (GFP-VAMP-2 with SNAP-25 and syntaxin1A; GFP-SNAP-25 with VAMP-2 and syntaxin1A; GFP-syntaxin1A with VAMP-2 and SNAP-25) (Shirasu et al. 2000). The distribution patterns of the overexpressed SNARE proteins were similar to those of the endogenous SNARE proteins: GFP-VAMP-2 on the membranes of vesicles, GFP-SNAP-25A on the cytoplasmic surface of the plasma membrane, and GFP-syntaxin1A on the surface plasma membrane (Figure 1). These findings suggest that overexpressed exogenous VAMP-2, SNAP-25A, and syntaxin1A could function in the same manner as endogenous SNAREs.
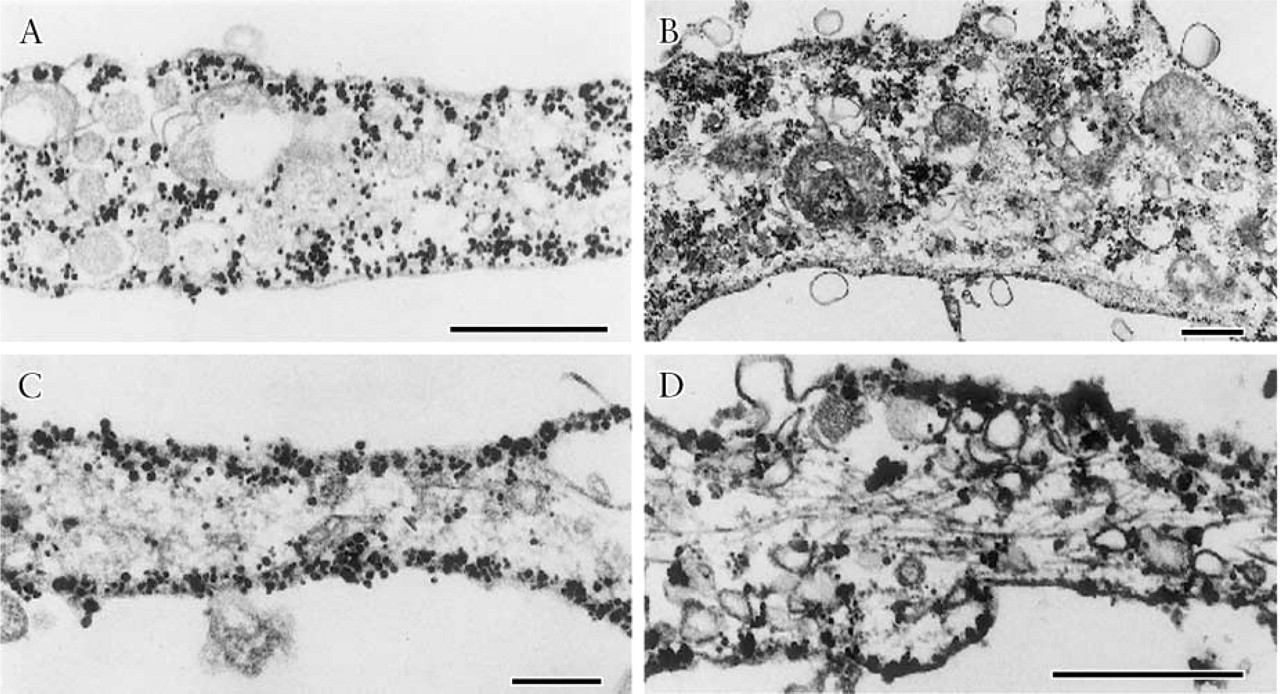
Immunogold electron micrographs showing the localization of GFP-labeled SNAREs detected by the anti-GFP antibody in neurites of PC12 cells that overexpressed GFP-SNAREs (Shirasu et al. 2000). (
Figure 2 shows fluorescence photomicrographs of PC12 cells 72 hr after transfection. The neurites of GFP-VAMP-2-expressing cells were longer than those of GFP-expressing cells. After transfection with the GFP-SNAP-25A, the PC12 cells had more neurites per cell than the GFP-expressing cells. The pattern of neurite outgrowth of cells transfected with the GFP-syntaxin1A was the same as that of GFP-expressing cells. To analyze in more detail these morphological changes due to transfection with GFP-SNAREs, we performed quantitative analysis on PC12 cells, focusing on the following parameters: neurite length per individual neurite, total neurite length per cell, and number of neurites per cell (Figure 3). The mean individual neurite length significantly increased for the GFP-VAMP-2-expressing cells. The total neurite length per cell significantly increased not only in cells expressing GFP-VAMP-2 but also in GFP-SNAP-25A-expressing cells. The average number of neurites per cell was significantly higher in the cells expressing GFP-SNAP-25A than in the control GFP-expressing cells. These findings indicate that overexpression of GFP-VAMP-2 increased the total neurite length per cell through an increase in the length of individual neurites and that overexpression of GFP-SNAP-25A increased the total neurite length through an increase in the number of neurites. No significant changes were observed for these parameters in the cells expressing GFP-syntaxin1A.
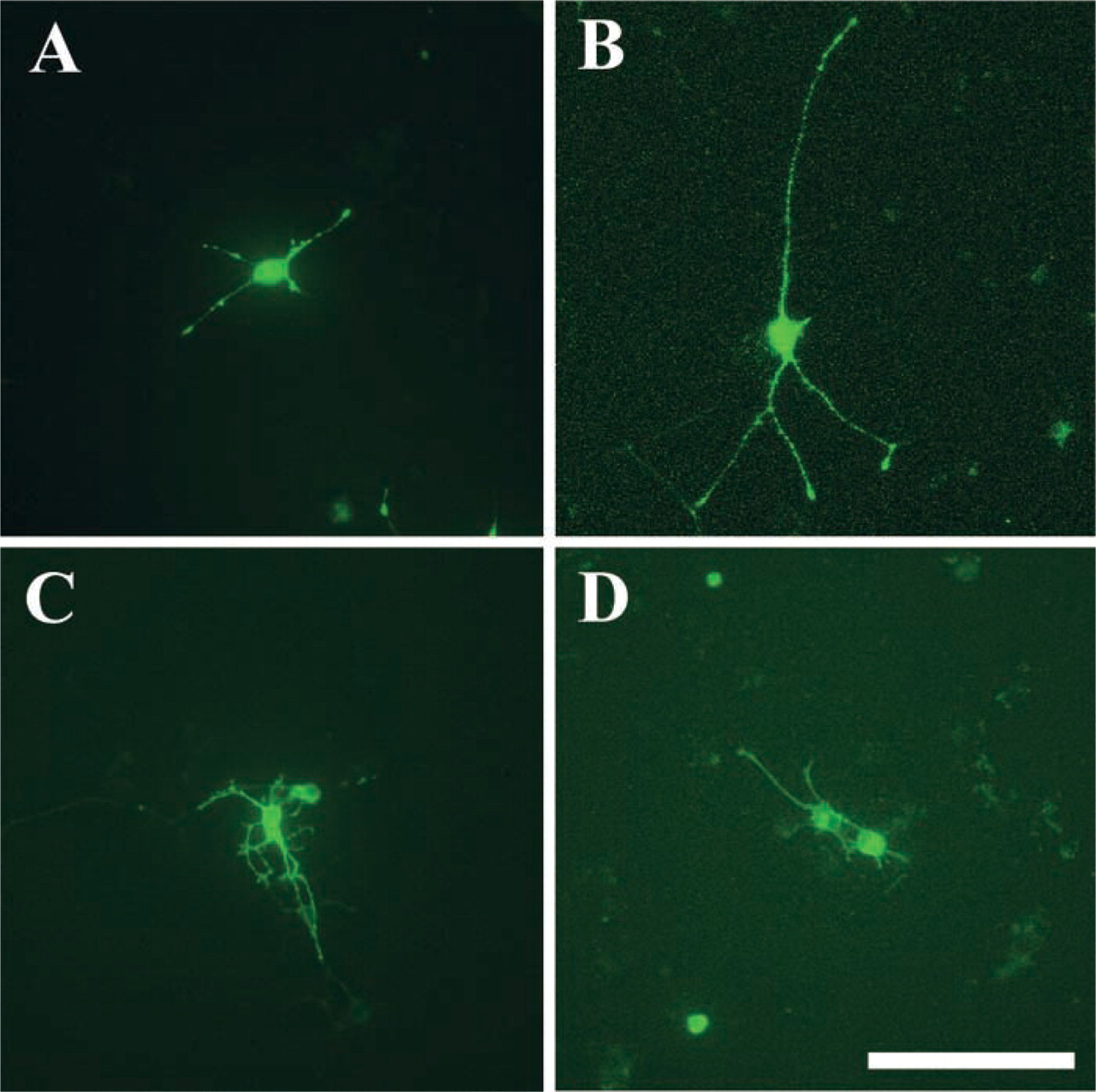
Fluorescence photomicrographs showing PC12 cells 72 hr after transfection and replacement with growth medium containing nerve growth factor (Shirasu et al. 2000). Living cells were examined by confocal laser scanning microscopy. These micrographs show the representative features of cells in which GFP (
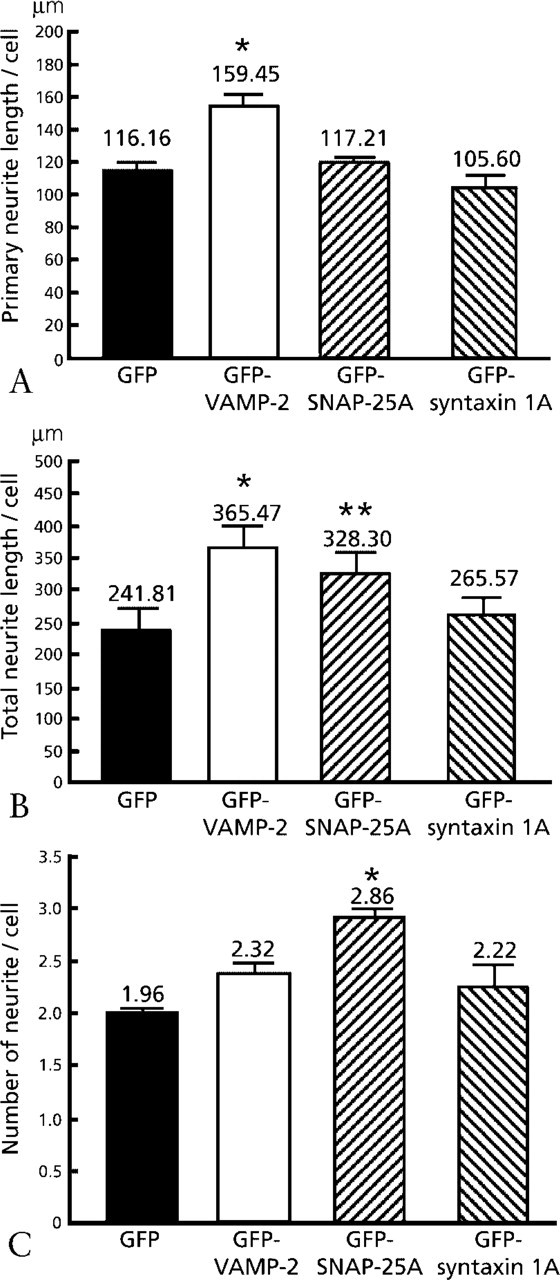
Effect of overexpression of GFP-SNARE proteins on neurite out growth from PC12 cells (Shirasu et al. 2000). (
In the study just described, we demonstrated that, when overexpressed in PC12 cells, VAMP-2 promoted neurite elongation, whereas SNAP-25A stimulated neurite sprouting. On the other hand, overexpression of syntaxin1A neither promoted nor inhibited neurite outgrowth. Thus, VAMP-2 and SNAP-25A play different roles, the former in neurite elongation and the latter in sprouting (Table 2). Although it was reported that cleavage of VAMP, with tetanus neurotoxin or botulinum neurotoxin B, had no effect on neurite elongation in hippocampal neurons (Osen-Sand et al. 1996), our findings indicate that VAMP-2 has a strong positive effect on neurite elongation in PC12 cells. This discrepancy is probably due to the existence of toxin-resistant VAMP (TI-VAMP) (Galli et al. 1998; Martinez-Arca et al. 2000) in PC12 cells. The finding that VAMP-2, a v-SNARE, promoted the elongation of individual neurites is in accordance with the hypothesis that the SNARE complex is involved in vesicle fusion for neurite elongation. The overexpression of VAMP-2 can therefore be considered to activate the fusion of vesicles with the plasma membrane for membrane expansion. Alternatively, the overexpression of VAMP-2 could support vigorous neurite initiation within 72 hr. Because we measured neurite length only 72 hr after transfection, over a certain time period the neurite growth rate might be not different.
Effect of overexpression of SNARE proteins on neurite outgrowth
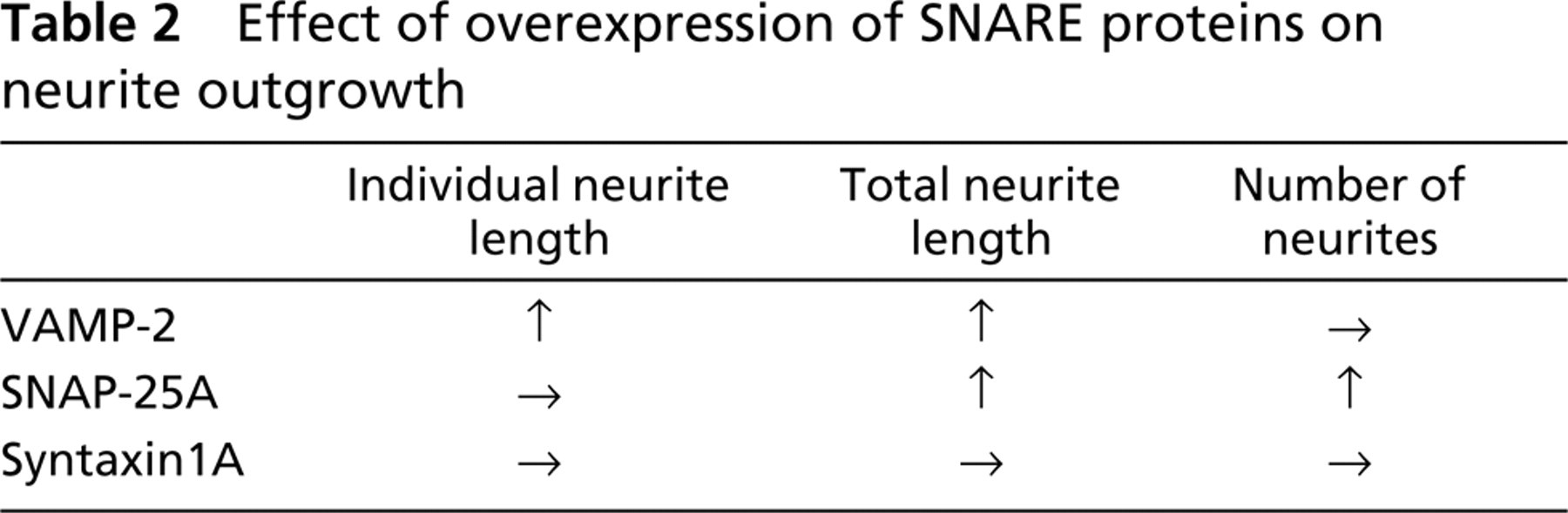
SNAP-25A promoted neurite sprouting while having no clear effect on neurite elongation. It is likely that the overexpression of SNAP-25A increased the amount of surface plasma membrane-associated SNAP-25A, leading to an increase in the number of sites for vesicle fusion and thus to enhancement of neurite sprouting. Inhibition of SNAP-25 expression by antisense oligonucleotides prevented neurite extension in rat cortical neurons and PC12 cells in vitro (Osen-Sand et al. 1993). Individual neurite length was also decreased by cleavage of SNAP-25 with botulinum neurotoxin A (Morihara et al. 1999). On the other hand, it was reported that total neurite length was decreased by the cleavage of SNAP-25 with botulinum neurotoxin A (Osen-Sand et al. 1996) and via inhibition of SNAP-25 expression by antisense oligonucleotides (Osen-Sand et al. 1993). These studies are compatible with our study, which showed that the total neurite length per cell increased in SNAP-25A-overexpressing cells, indicating that SNAP-25A plays a positive role not only in elongation but also in sprouting of neurites.
Overexpression of GFP-syntaxin1A had no effect on neurite elongation or neurite sprouting in our study described above. The precise role of syntaxin in neurite outgrowth remains unclear. There are contradictory reports concerning the effect of syntaxin inhibition on neurite outgrowth. Inhibition of syntaxin1A by antisense oligonucleotides increased neurite sprouting and neurite elongation in cultured rat dorsal root ganglion neurons (Yamaguchi et al. 1996). Similarly, the application of antibodies against syntaxin1A caused excessive sprouting from the severed ends of neurites of cultured chick retinal ganglion neurons (Yamaguchi et al. 1996). In contrast, it was demonstrated that cleavage of syntaxin with botulinum neurotoxin C1 inhibited neurite outgrowth in cultured chick dorsal ganglion neurons (Igarashi et al. 1996). On the other hand, it was reported that botulinum neurotoxin C1 cleaved not only syntaxin but also SNAP-25 in cultured spinal cord neurons (Williamson et al. 1996), thus having a severe cytotoxic effect on neurons (Williamson and Neale 1998). Overexpression of syntaxin1A by adenovirus inhibited neurite extension in PC12 cells (Zhou et al. 2000). Because the overexpression of syntaxin1A had no significant effect on neurite sprouting or elongation, it is conceivable that, in PC12 cells, interaction with some other members of SNARE family prevents syntaxin 1A from functioning to accelerate neurite sprouting or elongation. Further studies will be needed to clarify the detailed mechanism of the membrane expansion and the regulation of the cytoskeleton in neurite outgrowth and axon guidance.
Footnotes
Acknowledgements
Supported in part by a grant-in-aid for Scientific Research from the Japanese Ministry of Education, Culture, Sports, Science and Technology, and by Health Sciences Research Grants for Research on Brain Science.
We thank Dr Masami Takahashi (Kitazato University) for fruitful collaborations.