
Abstract
The C-terminal globular endostatin domain of collagen type XVIII is anti-angiogenic in a variety of experimental tumor models, and clinical trials to test it as an anti-tumor agent are already under way. In contrast, many of its cell biological properties are still unknown. We systematically localized the mRNA of collagen type XVIII with the help of in situ hybridization (ISH) and detected it in epithelial and mesenchymal cells of almost all organ systems throughout mouse development. Light and electron microscopic immunohistochemistry (IHC) revealed that the endostatin domain is a widespread component of almost all epithelial basement membranes in all major developing organs, and in all basement membranes of capillaries and blood vessels. Furthermore, quantitative immunogold double labeling demonstrated a co-localization of 50% of the detected endostatin domain together with perlecan in basement membranes in vivo. We conclude that the endostatin domain of collagen type XVIII plays a role, even in early stages of mouse development, other than regulating angiogenesis. In the adult, the endostatin domain could well be involved in connecting collagen type XVIII to the basement membrane scaffolds. At least in part, perlecan appears to be an adaptor molecule for the endostatin domain in basement membranes in vivo.
Keywords
A
Collagen type XVIII protein was found in basement membrane zones of embryonic chick and adult murine blood vessels (Muragaki et al. 1995; Halfter et al. 1998). In humans, the collagen type XVIII mRNA was seen in several tissue extracts, including heart, kidney, liver, and intestine (Saarela et al. 1998b). The corresponding protein was seen with the help of IHC to be a ubiquitous component of basement membrane zones of many adult organs and especially of blood vessels (Saarela et al. 1998a), and in fibrotic human liver (Musso et al. 1998). Recently mutations in the α1-chain of collagen type XVIII have been identified in families suffering from the Knobloch syndrome, a disease involving retinal detachment (Sertie et al. 2000). Lack of the endostatin domain of collagen type XVIII also results in eye abnormalities in mice (Fukai et al. 2002). The progression of human hepatocellular carcinoma is associated with a decrease in the expression of the endostatin precursor of collagen type XVIII (Musso et al. 2001).
The α1-chain of collagen type XVIII is homologous with that of collagen type XV (Myers et al. 1992; Rehn and Pihlajaniemi 1994). Large amounts of murine collagen type XV mRNA were seen in heart and skeletal muscle, and collagen type XV knockout mice exhibit cardiovascular defects and a skeletal myopathy (Eklund et al. 2001). Collagen type XV mRNA was also found in a variety of human tissues, especially in endothelial cells (Kivirikko et al. 1995). Accordingly, the protein was mainly seen in basement membrane zones of capillaries and blood vessels (Hagg et al. 1997).
Endostatin is part of the larger C-terminal globular NC1 domain of collagen type XVIII (Oh et al. 1994; Rehn et al. 1994) and was originally isolated as an inhibitor of endothelial cell proliferation from hemangioendothelioma medium (O'Reilly et al. 1997). X-ray crystallography revealed a compact globular structure that resembles C-type lectins (Hohenester et al. 1998). The NC1 domain binds to perlecan, laminin-1, nidogen-2 and fibulin-2, whereas the endostatin domain itself shows higher affinity for fibulin-2, nidogen-2, and heparin in vitro. It was detected in adult mouse brain, skeletal muscle, heart, kidney, and liver by Western blotting. This analysis revealed tissue forms usually related in size to the NC1 domain, whereas the circulating form corresponds more closely to endostatin (Sasaki et al. 1998,2002). Investigation of blood vessels revealed that the elastic sheets of mouse aorta contain the greatest amount of endostatin thus far extracted from tissue, which seems to be associated through ternary complexes via fibulin-2 or nidogen-2 with elastin (Miosge et al. 1999).
The involvement of the endostatin domain of collagen type XVIII (endostatin-XVIII) in mouse development is still not quite clear. We localized endostatin-XVIII at the light and ultrastructural level and found it to be a true basement membrane component even early during development. To test binding partners of endostatin-XVIII demonstrated by solid-phase assays in vitro (Sasaki et al. 1998) for their in vivo relevance, we performed immunogold double labeling and show that endostatin-XVIII is co-localized with perlecan in basement membranes in vivo.
Materials and Methods
Animals
NMRI mice were kept on a normal day/night cycle. Day 0 of gestation was defined as starting at 1100 hr on the day on which a vaginal plug was detected after a mating period of 3 hr. On the respective days of gestation, beginning with day 8, pregnant mice were anesthetized with ether and sacrificed by cervical dislocation. After dissection of the uterine horns, the embryos (days 8, 10, 12) or fetuses (days 14, 16, 18) were removed. Three embryos or fetuses of each developmental stage were investigated. For light microscopic analysis of adult tissues, three kidneys from different 3-month-old NMRI mice were taken.
Fixation and Preparation of Tissues
All specimens were fixed by immersion in 4% paraformaldehyde in PBS, pH 7.2, at 4C. They were then dehydrated in an ascending series of ethanol from 30% to 100% and embedded in paraffin. Serial sections of 5 μm were cut with a Reichert's microtome. Every fifth section was stained with hematoxylin for topological orientation and staging was achieved by comparison with the appropriate Theiler stages. Tissue pieces (∼1 mm3) of adult and fetal NMRI mouse kidneys, fetal skin, and lung were applied for immunogold staining and were fixed in 4% paraformaldehyde and 0.5% glutaraldehyde for 15 min, dehydrated, and embedded in the acrylic resin LR Gold (London Resin Company; Reading, UK). Ultrathin sections were cut with a Reichert's ultramicrotome and collected on formvar-coated nickel grids for immunoelectron microscopy as described previously (Miosge et al. 2000).
Antibodies
Affinity-purified rabbit antibody (2.26 mg/ml) against the endostatin domain from mouse collagen type XVIII (Sasaki et al. 1998) was generated recombinantly from transfected mammalian cells and was a kind gift from Drs. T. Sasaki and R. Timpl (MPI for Biochemistry; Martinsried, Germany). No significant crossreactivity with the corresponding domain of collagen type XV was detected (Sasaki et al. 2000). Affinity-purified rabbit antibodies (1.53 mg/ml) against domain V of perlecan (Brown et al. 1997), 1.11 mg/ ml against nidogen-1 (Fox et al. 1991), and 3.4 mg/ml against fibulin-2 (Pan et al. 1993) were used. The antibodies against perlecan, nidogen-1, and fibulin-2 were used to coat 16-nm colloidal gold particles according to standard protocols (Miosge et al. 2000). Furthermore, an affinity-purified goat anti-rabbit IgG (1.0 mg/ml) was obtained from Medac (Hamburg, Germany) and applied to coat 8-nm colloidal gold particles.
Light Microscopic Immunohistochemistry
Sections were deparaffinized, rehydrated, and rinsed for 10 min in PBS. Endogenous peroxidase was blocked by incubation in 3% H2O2 in methanol for 45 min in the dark. Each of the reaction steps was followed by rinsing for 10 min in PBS. Sections were pretreated for 5 min with 10 μg/ml protease XXIV (Sigma; Deisenhofen, Germany). The anti-endostatin-XVIII antibody was used at a dilution of 1:100 for 1 hr at 37C. The anti-perlecan antibody was applied at a dilution of 1:100 for 24 hr at 37C and the anti-nidogen-1 and anti-fibulin-2 antibodies at a dilution of 1:100, all in PBS, for 1 hr at RT. A peroxidase-anti-peroxidase method followed the previously described procedures (Miosge et al. 2000). As negative controls, normal rabbit IgGs and the corresponding preimmune sera were used instead of the primary antibodies, at similar concentrations. No immunostaining was observed.
Electron Microscopic Immunohistochemistry
For single labeling, the tissue sections were incubated for 1 hr at 4C with the antibodies against endostatin-XVIII. Sections were rinsed in PBS and incubated with the 8-nm gold-coupled goat anti-rabbit IgG diluted 1:200 in PBS for 20 min at RT. Further sections for single labeling were incubated for 1 hr at 4C with the antibodies against perlecan (1:50), nidogen-1 (1:100), or fibulin-2 (1:50) directly coupled to 16-nm colloidal gold particles. Thereafter, all sections were rinsed with water and stained with uranyl acetate (10 min) and lead citrate (8 min). Sections were examined with a LEO 906E electron microscope. For double labeling, all sections were incubated for 5 min at RT with 1% bovine serum albumin (BSA) in PBS and then rinsed in PBS. Thereafter, anti-endostatin-XVIII antibody diluted 1:50 in PBS was applied for 1 hr at 4C. After being rinsed with PBS, the gold-coated (8 nm) goat anti-rabbit antibody diluted 1:200 was applied for 20 min at RT. The sections were thoroughly rinsed with PBS. The gold-coated (16 nm) anti-perlecan antibody diluted 1:50 in PBS or the anti-nidogen-1 antibody diluted 1:100 in PBS was then incubated. All sections were finally rinsed with water and stained with uranyl acetate (10 min) and lead citrate (8 min). As control for the double labeling experiments we also applied a monoclonal (rat) endostatin-XVIII antibody (Tomono et al. 2002) as shown in Miosge et al. (1999). This approach avoids possible crosslinks between the two rabbit IgGs used in the double labeling described above and yielded identical results (data not shown).
Statistical Analyses
The numbers of gold particles, shown as means± SEM, were counted on 10 randomly chosen micrographs of identical size and magnification. Each micrograph included a basement membrane of approximately 120 nm in width and 6.5 μm in length. The number of gold particles representing individual labeling and co-localizations, defined as different gold particles less than 30 nm apart, was determined for proximal tubule basement membranes after double labeling. With the immunogold method, a correlation between the number of gold particles and the relative protein amount can be established (Miosge et al. 2002). Statistically significant differences with p values <0.001 (Wilcoxon-Mann-Whitney test for unpaired samples) were noted between all reactions and their appropriate controls.
In Situ Hybridization
A 205-bp sac/hinc II fragment corresponding to the C-terminal endostatin domain of collagen type XVIII was used for hybridization (Sasaki et al. 1998). Nonradioactively-labeled sense and anti-sense RNAs were produced in vitro with the digoxigenin (DIG) labeling kit (Roche Diagnostics; Mannheim, Germany). Paraffin sections were deparaffinized, re-hydrated, and pretreated with proteinase K, then incubated with the hybridization solution (50% formamide, 5 × SSC, 0.1 mg/μl yeast tRNA) for 16 hr at 50C. The RNA concentration for collagen type XVIII was 100 ng digoxigenin-labeled antisense probe in 100 μl hybridization solution per section. In control sections, an equivalently labeled amount of sense RNA was used instead of anti-sense RNA. Posthybridization treatment included washes with 1 × SSC (twice for 15 min at 50C), 1 × SSC (twice for 15 min), 0.1 × SSC (four times for 15 min), both at 60C, and finally PBS for 15 min at RT. Specimens were subsequently incubated with a 1:60 dilution of anti-DIG phosphatase-labeled antibody (Roche Diagnostics) in PBS for 1 hr at RT (Miosge et al. 1998). As controls, the procedure described above was carried out either without any probes or with omission of the antibody.
Results
Light Microscopic Localization of Collagen Type XVIII mRNA During Mouse Development
ISH revealed the mRNA of collagen type XVIII on days 8, 10, 12, 14, 16, and 18 of mouse development. On day 8 the mRNA was detected only in the mesoderm, not in the ectoderm or endoderm. In addition, the ectoplacental conus was positive for the mRNA (data not shown). On day 10 the mRNA was located widespread in the cytoplasm of epithelial and mesenchymal cells, e.g., the structures of the developing head mesenchyme, the gut, the neural tube and ganglia, or the heart and blood vessels (Figure 1A). From day 12 onwards, the collagen type XVIII mRNA was seen in the mesenchymal and epithelial cells of all major organ anlagen, e.g., the kidney, liver, or, as shown here, the lung (Figure 1B). In the vascular system of all developmental stages investigated, all endothelial cells of the developing capillaries stained for the mRNA, as seen here in the lung mesenchyme on day 14 (Figure 1B). On day 16 the mRNA of collagen type XVIII was detected, e.g., in the epithelial cells of the developing skin, whereas the mesenchymal cells of the developing dermis remained unstained (Figure 1C). None of the control ISHs with the corresponding sense probes (Figures 1D-1F) showed any staining.
Light Microscopic Localization of Endostatin-XVIII During Mouse Development
Day 8 mouse embryos are still surrounded by Reichert's membrane, a thick and multilayered basement membrane, which stained positive for endostatin-XVIII. In the early embryo the three germ layers, endoderm, ectoderm, and mesoderm, are separated by basement membranes exhibiting staining for the protein (Figure 2A). From day 10 to day 18, staining for endostatin-XVIII was seen in almost all epithelial basement membrane zones in all consecutive stages of development. In addition, the endothelial basement membrane zone of capillaries and larger blood vessels revealed staining for the protein (Table 1). In the brain, the neuroectodermal cells themselves were not stained. In contrast, the basement membrane zones of the neuroectoderm and the adjacent leptomeningeal anlage were positive for endostatin-XVIII (Figure 2B). In the peripheral nervous system, the basement membrane zones of the ganglia exhibited staining, as did the endoneurium of peripheral nerves (Figure 2C). No staining was seen in any developmental stages of bone formation, either in the condensed mesenchyme or the chondrocytes or at the stage of onset of calcification. The same was true for the myocytes of the developing skeletal muscles (Figure 2D). The basement membrane zones of the skin stained positive for endostatin-XVIII, but neither the fibroblasts, nor the keratinocytes showed any staining (Figure 2E). In the lung anlage, neither the mesenchyme nor the developing epithelial cells were stained, but the protein was found in the basement membrane zones of the branching bronchi (Figure 2F). The cells of the developing myocardium, endocardium, and pericardium were never stained, but their corresponding basement membrane zones were positive for endostatin-XVIII. In the liver anlage, the protein was found in the sinusoids and underneath the mesothelium. The hepatocytes and the hematopoietic cells showed no staining (Figure 2G). The basement membrane zones of the developing exocrine glands in the pancreas anlage and the salivary glands in the head mesenchyme were positive for endostatin-XVIII. In the small and large intestine the epithelial cells were not stained, whereas the basement membrane zones underlying the developing epithelium were stained for endostatin-XVIII. In all stages of kidney organogenesis, only basement membrane zones of the consecutive stages of glomeruli development (comma, S-shaped, early glomeruli) and those of the tubules were positive for endostatin-XVIII. The surrounding mesenchyme was never stained (Figure 2H).
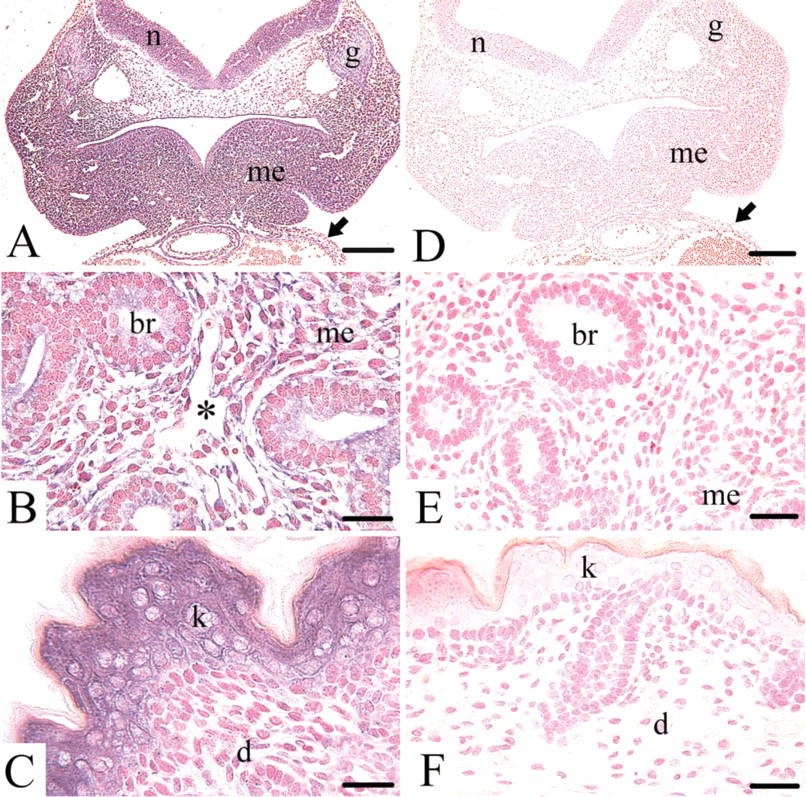
Summary of the ISH for collagen type XVIII mRNA during mouse development. (
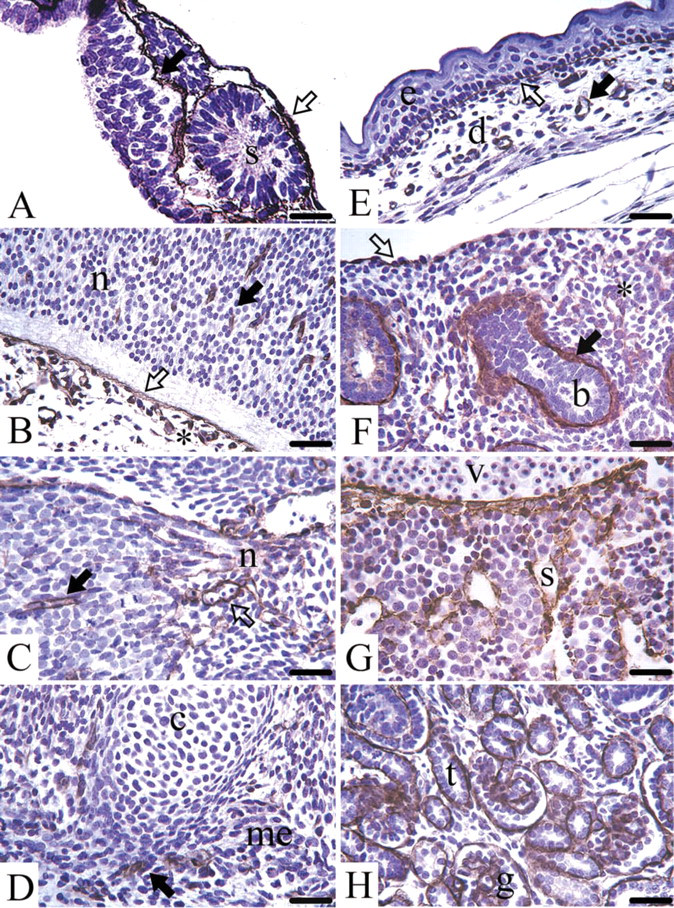
Light microscopic localization of endostatin-XVIII during mouse development. (
Localization of endostatin-XVIII during mouse development
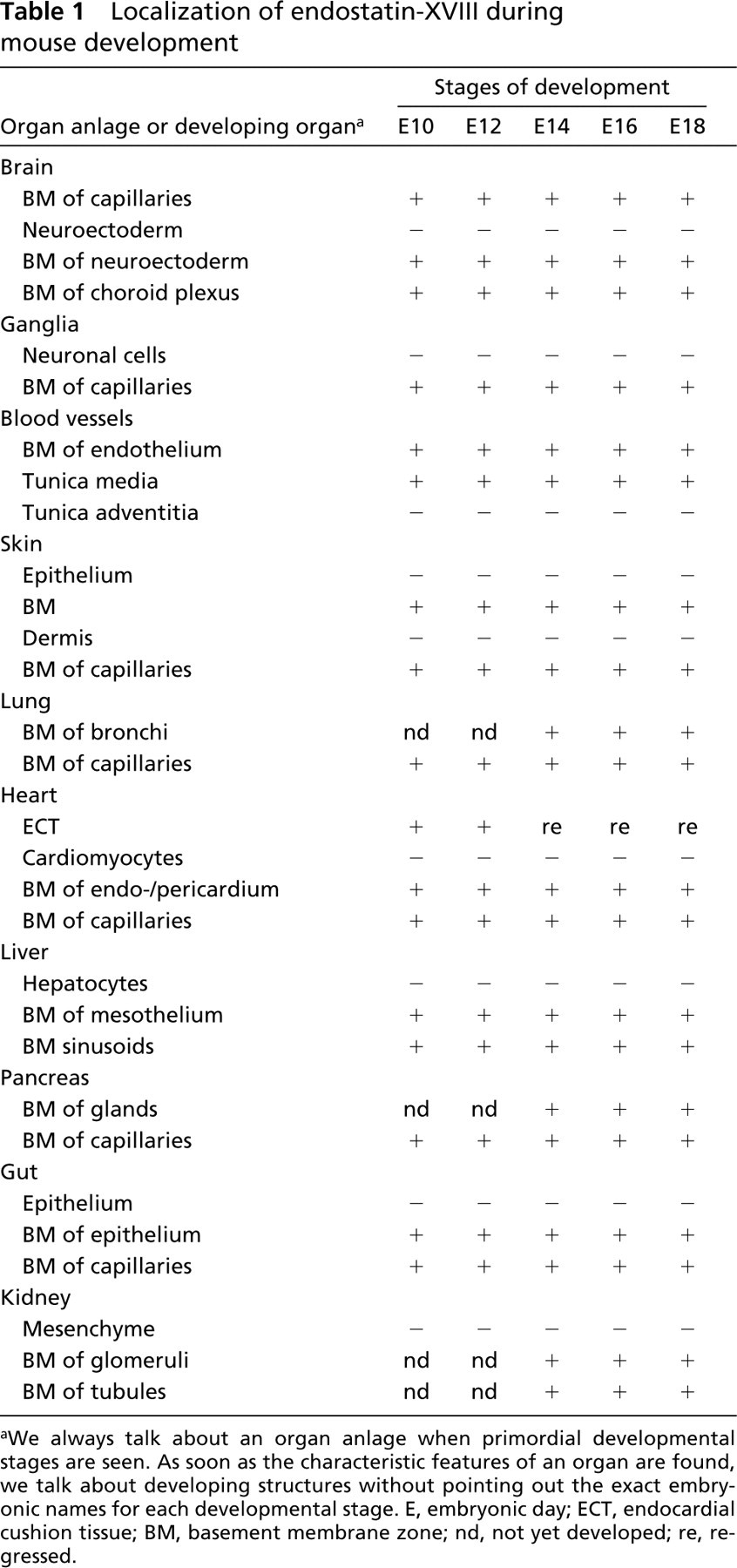
aWe always talk about an organ anlage when primordial developmental stages are seen. As soon as the characteristic features of an organ are found, we talk about developing structures without pointing out the exact embryonic names for each developmental stage. E, embryonic day; ECT, endocardial cushion tissue; BM, basement membrane zone; nd, not yet developed; re, regressed.
Ultrastructural Localization of Endostatin-XVIII During Mouse Development
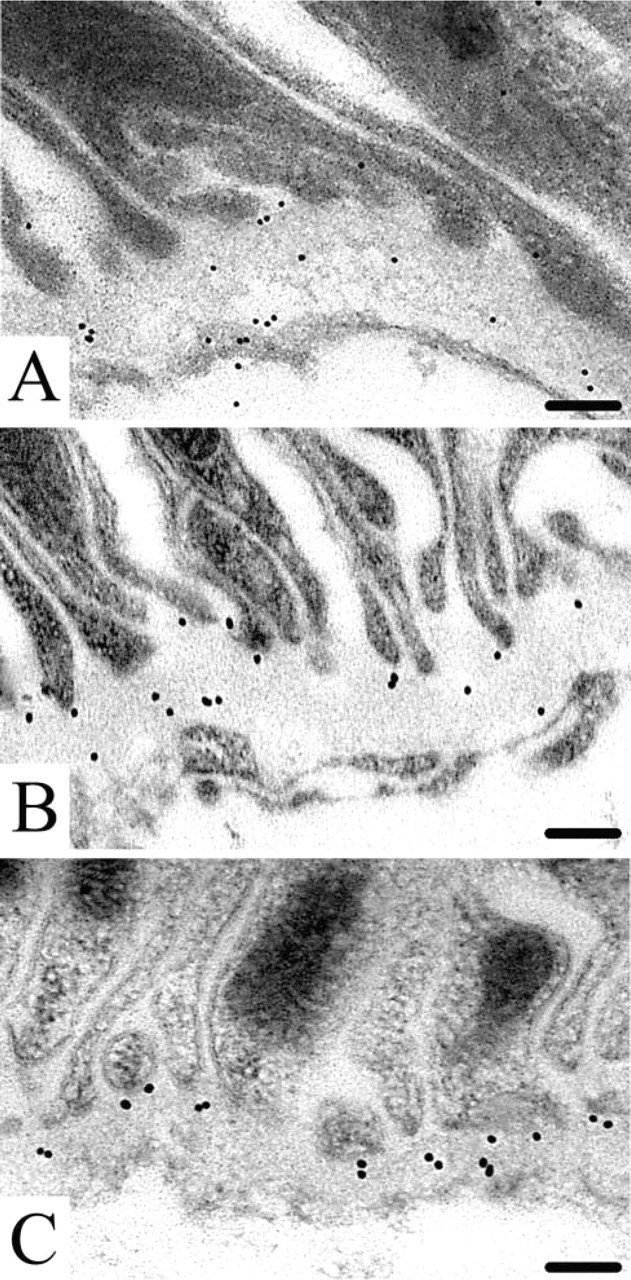
Immunogold histochemistry at the ultrastructural level revealed that the endostatin domain of collagen type XVIII is indeed a true component of developing basement membranes in the mouse embryo and fetus. Endostatin-XVIII was found in the basement membranes of the developing epithelial cells in the lung anlage (Figure 3A). In the kidney, endostatin-XVIII was detected in the basement membranes of developing tubules (Figure 3B). The same was true for the basement membranes underneath endothelial cells, as shown here for a capillary in the dermis. The extracellular matrix, adjacent to the capillary, also stained positive (Figure 3C).
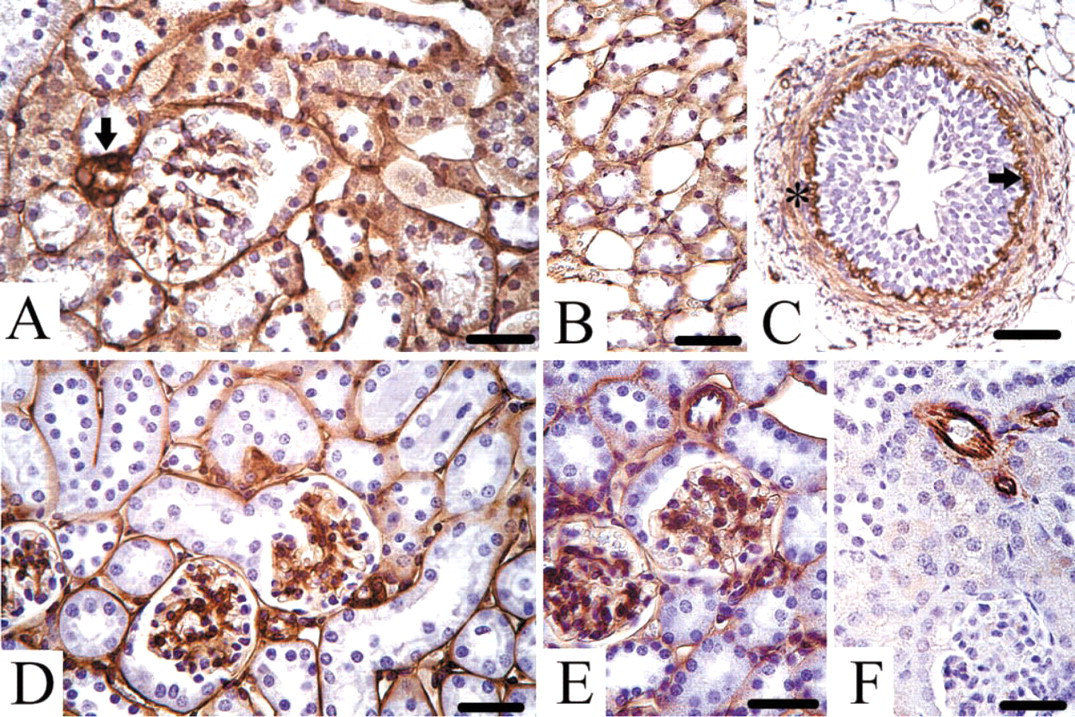
Light Microscopic Localization of Endostatin-XVIII, Perlecan, Nidogen-1, and Fibulin-2 in Adult Mouse Kidney
Endostatin-XVIII was found in adult mouse kidney, especially in the basement membrane zones of the Bowman's capsule and the proximal and distal tubules. The protein was also detected in connective tissue surrounding arterioles (Figure 4A). The basement membrane zones of the collecting ducts, loops of Henle (Figure 4B), and the ureter (Figure 4C) stained positive. The urothelium itself was free of any staining, but the lamina propria and lamina muscularis contained endostatin-XVIII next to elastic fibers. Perlecan (Figure 4D) and nidogen-1 (Figure 4E) were found in all basement membrane zones in the mouse kidney, including tubules and glomeruli as well as Bowman's capsule. Fibulin-2 was detected only in the walls of arterioles in connection with elastic fibers. In contrast, no tubular or glomerular basement membrane zones stained for fibulin-2 (Figure 4F).
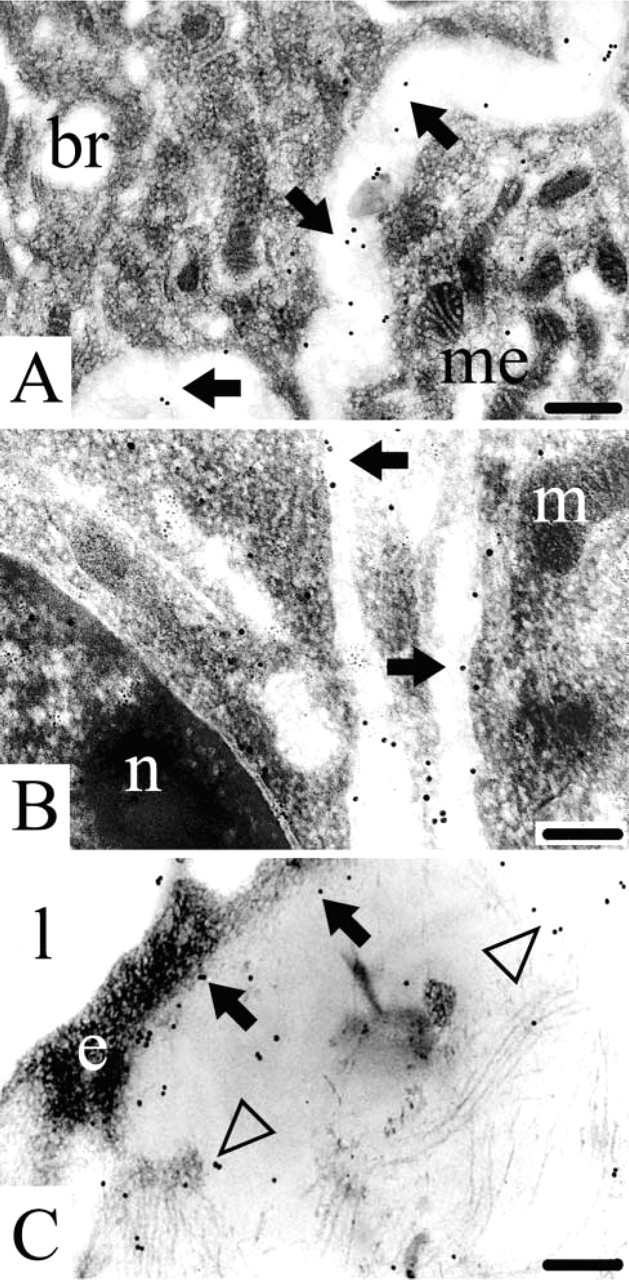
Electron microscopic localization of endostatin-XVIII during mouse development. (
Ultrastructural Localization of Endostatin-XVIII, Nidogen-1, and Perlecan in Adult Mouse Kidney
We found endostatin-XVIII in the proximal (Figure 5A) and distal tubules as well as in the Bowman's capsule. Less labeling was seen in the basement membranes of the glomeruli and some intracellular labeling was seen. Free endostatin-XVIII was also detectable in the serum and urine (Sasaki et al. 1998; John et al. 1999). Perlecan (Figure 5B) and nidogen-1 (Figure 5C) were detected in all of the basement membranes of the kidney, as shown here for the adult proximal tubule.
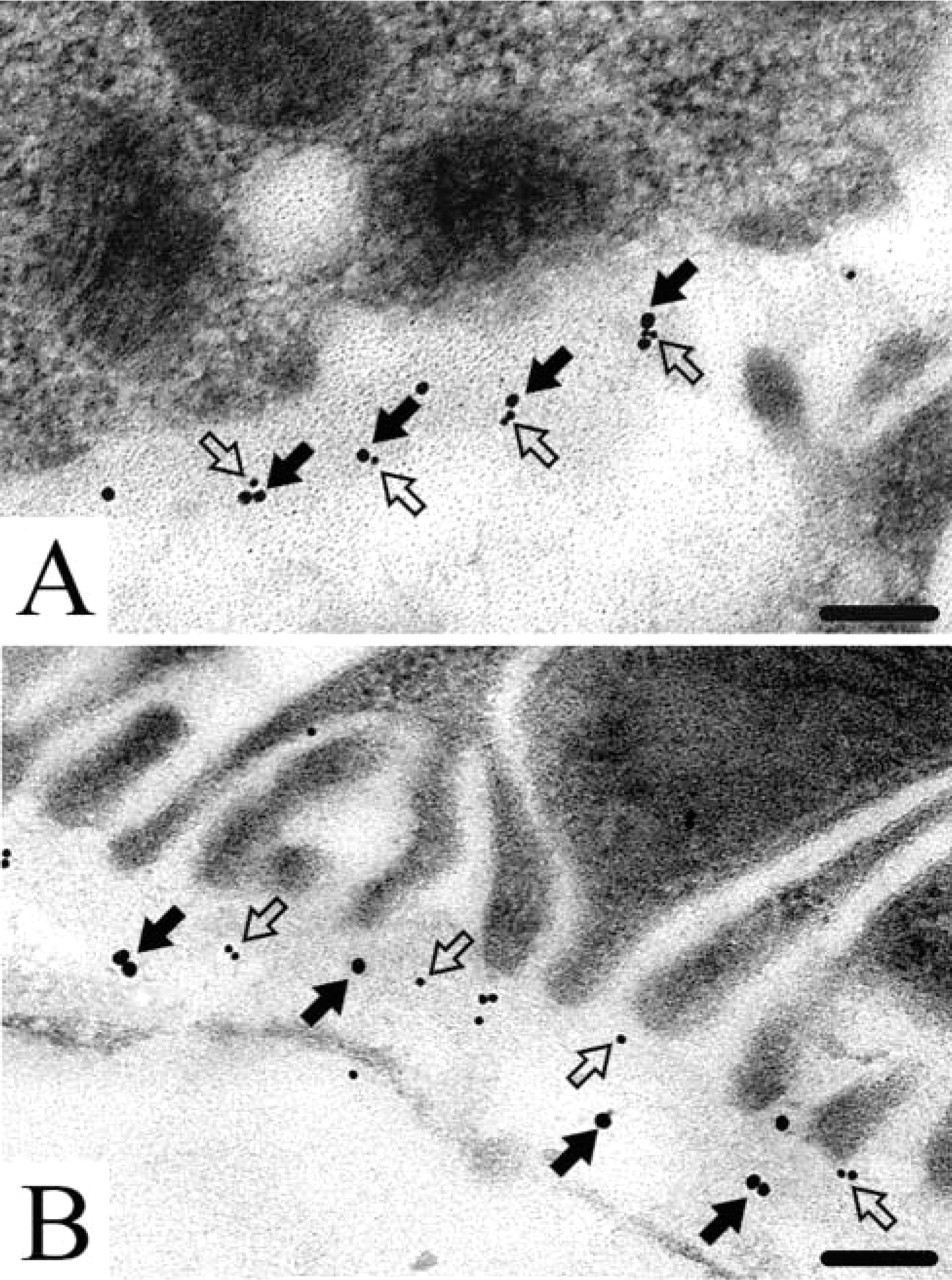
Immunogold Double Labeling of Endostatin-XVIII with Nidogen-1 and Perlecan
Because the endostatin domain of collagen type XVIII exhibits various binding activities in solid-phase assays (Sasaki et al. 1998), we performed double labeling for potential binding partners. Perlecan was co-localized with endostatin-XVIII in the basement membranes of the proximal tubules in the adult mouse kidney (Figure 6A). A co-localization was classified as a potential molecular contact if the distance between the different sizes of gold label (8 nm or 16 nm) was less than 30 nm (Miosge et al. 1999). In contrast, nidogen-1 showed no co-localization with endostatin-XVIII. Labeling was always seen at some distance away from labeling for nidogen-1 (Figure 6B).
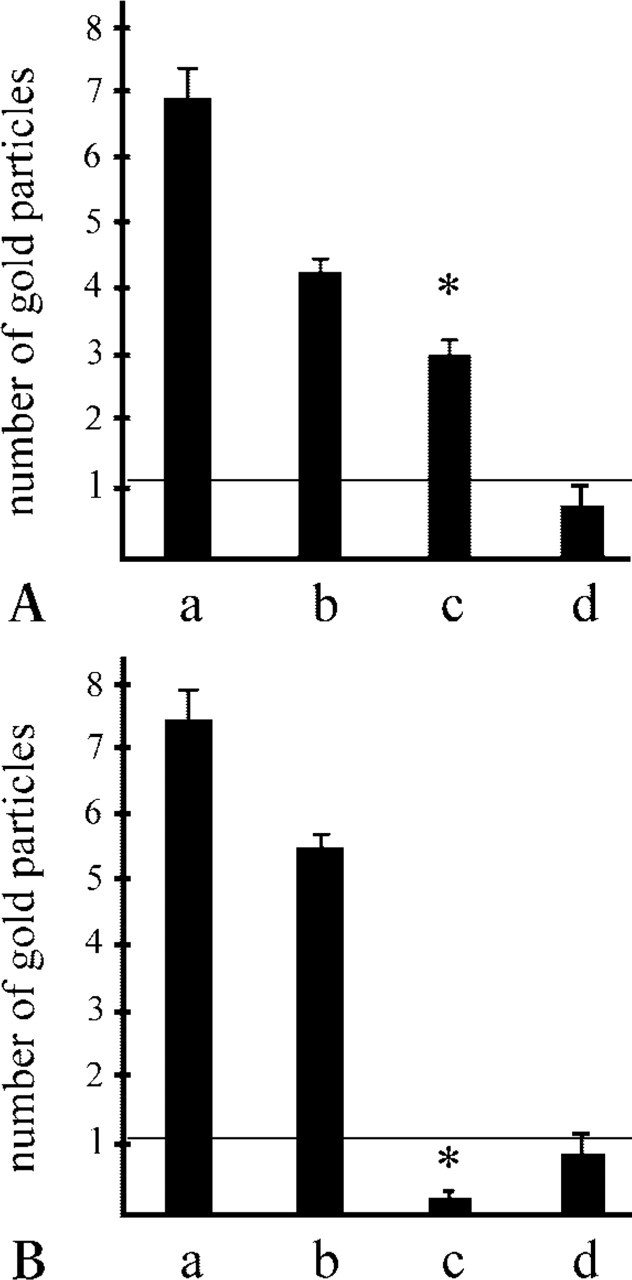
Statistical Analysis of the Immunogold Results
The amount of endostatin-XVIII detected in the two co-localization experiments was almost identical (Figures 7A and 7B, Lanes a). Approximately 50% of the detectable endostatin-XVIII (Figure 7A, Lane a) was co-localized with perlecan (Figure 7A, Lane c). In contrast to the amount of nidogen-1 detected (Figure 7B, Lane b), the co-localization rate with nidogen-1 (Figure 7B, Lane c) was below the background level (Figure 7B, Lane d). The difference between the co-localization rates of nidogen-1 and perlecan with endostatin-XVIII was statistically significant with p values <0.001 (Figures 7A and 7B, Lanes c, asterisks).
Discussion
The C-terminal globular endostatin domain of collagen type XVIII is of special interest because of its putative value as an anti-angiogenic agent in human antitumor therapy (Nelson 1998). It is known that the endostatin domain needs proteolytic liberation from collagen type XVIII for its anti-angiogenic action (Marneros and Olson 2001; Sasaki et al. 2002). The anti-angiogenic effect of endostatin-XVIII is believed to depend on heparin binding to antagonize FGF-induced endothelial cell proliferation (Sasaki et al. 1999) and/ or a mechanism involving blockage of VEGF-induced endothelial cell migration (Yamaguchi et al. 1999). The activation of apoptosis of endothelial cells by endostatin-XVIII is also under discussion (Dhanabal et al. 1999; Dixelius et al. 2000). Recently it has been demonstrated that its anti-angiogenic activity involves the modulation of focal adhesions (Wickstrom et al. 2001) and the inhibition of αv and α5 integrin-dependent endothelial cell migration (Rehn et al. 2001). Glypicans were shown to act as low-affinity receptors for endostatin-XVIII (Karumanchi et al. 2001).
Light microscopic location of endostatin-XVIII (
As one of the first steps to further elucidate the physiological role of collagen type XVIII during mouse development, we localized its mRNA. Expression is seen in the mesoderm, but not in the ecto- or endoderm, at early stages of development and is later found in the mesenchyme. From day 12 onwards, the epithelial cell compartments also express collagen type XVIII mRNA. Interestingly, we found the mRNA in both cell compartments, epithelial and mesenchymal, throughout kidney and lung development. Collagen type XVIII is implicated in kidney and lung patterning processes during epithelial-mesenchymal tissue interaction in an in vitro culture system (Lin et al. 2001). Our results indicate that the switch in collagen type XVIII localization during the branching process seen by Lin et al. (2001) could be achieved by translating either the mRNA produced by the mesenchyme or the epithelium during organogenesis of kidney and lung. Schuppan et al. (1998) described the adult human hepatocyte as a source of endostatin-XVIII. We can now demonstrate that the murine hepatocytes produce the mRNA from the earliest developmental stages onwards. Endostatin-XVIII interferes with blood vessel maturation during cutaneous wound healing (Bloch et al. 2000). During mouse development, collagen type XVIII mRNA is produced by the keratinocytes. Whether this is the source of endostatin-XVIII involved in the wound healing process or whether endostatin-XVIII from the blood circulation is used remains to be established.
Immunogold histochemistry for endostatin-XVIII (
In agreement with the wide distribution of the collagen type XVIII mRNA early on during mouse development, the protein is also seen from day 8 onwards. The antibody applied detects the C-terminal domain of collagen type XVIII, which means that we cannot distinguish between free and bound endostatin. Therefore, it remains to be elucidated whether the whole molecule plays a role during development or whether the endostatin domain is cleaved from the molecule but remains in the same tissue location. Endostatin-XVIII was seen in all epithelial and endothelial basement membranes from day 8 onwards in all developing organs investigated, and might therefore be involved in early developmental processes. It appears to be a basement membrane component as abundant as collagen type IV. This was also recently demonstrated for adult human tissues (Tomono et al. 2002). Therefore, endostatin-XVIII could be involved in developmental processes other then regulating angiogenesis, e.g., kidney and lung patterning processes, as intriguingly demonstrated by Lin et al. (2001). Interestingly, no expression was seen during bone formation. This process in particular should require an anti-angiogenic molecule to hinder the vascularization of the cartilage, but here endostatin-XVIII appears not to be involved. Ackley et al. (2001) showed that the deletion or over-expression of endostatin-XVIII in C. elegans causes neuronal guidance defects. During mouse development, endostatin-XVIII is present in basement membrane zones of the central and peripheral nervous systems. However, its role in vertebrate neuronal development remains to be elucidated.
Statistical analysis. Bars represent the mean values with SEM of the number of gold particles in the two double-labeling experiments. (
As determined by immunogold histochemistry, endostatin-XVIII is a true basement membrane component even early during mouse development. Collagen type XVIII might be an integral basement membrane component involved in maintaining its structural network integrity. On the other hand, cleavage of the endostatin domain from the collagen type XVIII molecule is one of the main features of its anti-angiogenic function (Marneros and Olson 2001; Sasaki et al. 2002). It is tempting to speculate that basement membrane collagen type XVIII serves as a reservoir for the endostatin domain to counteract pro-angiogenic growth factors also bound to the extracellular matrix. Previously, we found that endostatin-XVIII is bound via nidogen-2 and fibulin-2 to elastic sheets of larger blood vessels, where it appears to be stored as its free form (Miosge et al. 1999).
To investigate the association of endostatin-XVIII with the known basement membrane components in vivo, we performed immunogold double labeling of endostatin-XVIII together with nidogen-1 or perlecan. These were chosen because their binding affinities were demonstrated in vitro (Sasaki et al. 1998). In preparation for the immunogold double labeling, we localized endostatin-XVIII, nidogen-1, perlecan, and fibulin-2 at the light and electron microscopic level and found endostatin-XVIII, nidogen-1, and perlecan within basement membranes at the ultrastructural level. In contrast, fibulin-2 is not a basement membrane component of adult mouse kidney. Quantitative immunogold double labeling revealed that approximately 50% of the detectable endostatin-XVIII is co-localized with perlecan in basement membranes in vivo. The level of co-localization of endostatin-XVIII with nidogen-1 was nearly zero. This could well mean that perlecan is one adaptor molecule for the endostatin domain in vivo to connect collagen type XVIII to the basement membrane, at least in adult mouse kidney basement membranes. At this time, one can only speculate about the biological relevance of this binding of the endostatin domain to perlecan, which might hinder its proteolytic activation in certain basement membranes or cell biological processes.
In summary, endostatin-XVIII is already found in the early mouse embryo and has a widespread distribution, mainly in basement membranes of epithelial and endothelial tissues. This supports a physiological role during organogenesis, not only for the regulation of angiogenesis. Expression of the mRNA is seen in epithelial as well as mesenchymal cells. In the adult mouse kidney, approximately 50% of the detectable endostatin domain is co-localized with perlecan in basement membranes. This seems to be at least one mechanism to connect collagen type XVIII to the basement membrane scaffold in vivo.
Footnotes
Acknowledgements
Supported by a grant from the German Research Council (DGF) to N. Miosge (Mi 573/2–1). Parts of this work were taken from the doctoral thesis of O. Abdul-Malak.
We would like to thank Drs R. Timpl and T. Sasaki for providing antibodies and cDNA, and Cyrilla Maelicke, BSc, for editing the manuscript.