
Abstract
Using RNA in situ hybridization to reveal cytoplasmic localization patterns of mRNAs in cultured cells, we noted unexpected staining of a cytoplasmic component in telophase cells. Control experiments revealed that the anti-digoxin-specific antibody was responsible for this staining. Because the staining was observed only at a position where both daughter cells are still connected, we identified the stained component as the midbody. This was confirmed by double staining of cells with anti-digoxin and antia -tubulin antibodies. We concluded that anti-digoxin-specific antibody shows crossreactivity with a component present in the midbody.
I
For many years we have successfully used digoxigenin-labeled probes in conjunction with anti-digoxigenin and anti-digoxin antibodies in our studies to specifically detect mRNA molecules at the light and electron microscopic levels (reviewed in Dirks 1996). However, as shown here, we noted an unexpected staining pattern in a subpopulation of cultured cells when we studied cytoplasmic localization patterns of mRNAs using an anti-digoxin-specific antibody. Control experiments, leaving out the specific probe, revealed staining of a discrete cytoplasmic component in late telophase cells, which also showed positive immunocytochemical staining for α-tubulin. These results suggest a nonspecific binding of the anti-digoxin antibody to one of the midbody components.
Materials and Methods
Cell Lines
X1 cells (HeLa cells containing a luciferase gene construct), human foreskin fibroblasts, and a rat fibroblast cell line (rat 9G) were grown on glass microscopic object slides at 37C in Dulbecco's minimal essential medium without phenol red (Gibco; Gaithersburg, MD), supplemented with 10% fetal calf serum, antibiotics, and glutamine (also from Gibco).
In Situ Hybridization
The in situ hybridization procedure was performed as described previously (Dirks et al. 1993). Briefly, in vitro cultured cells were washed in PBS and fixed for 20 min with 3.7% formaldehyde in PBS either with or without 5% acetic acid. Cells were then washed in PBS and stored in 70% ethanol at 4C. Before hybridization, cells were rehydrated in deionized water, incubated with 0.1% (w/v) pepsin (Sigma; St Louis, MO) in 0.01 M HCl for 1 min at 37C, briefly washed in water, dehydrated in a graded ethanol series, and air-dried. A 50-mer oligonucleotide (dT) and a 30-mer oligonucleotide 28S rRNA probe were labeled with Cy3-dUTP using terminal transferase. After labeling, these probes were precipitated in ethanol and dissolved in 20% formamide, 2 X SSC (0.3 M NaCl, 0.03 M sodium citrate) at a concentration of 1 ng/μl. A plasmid probe containing the cDNA of β-actin mRNA was labeled with digoxigenin-dUTP by nick translation, precipitated in ethanol, and dissolved in 60% formamide, 2 X SSC at a concentration of 5 ng/μl. Probes were applied to the pretreated cells, covered with a coverslip, and denatured for 3 min on an 80C metal plate. Hybridizations were performed for 3 hr in a moist chamber at room temperature (RT). After hybridization, cells were washed three times in 2 X SSC at RT when hybridized with an oligonucleotide probe and three times in 50% formamide/2 X SSC at 37C when hybridized with a nick-translated probe.
Immunocytochemical Detection
Antibodies used in this study were MAbs anti-digoxigenin (Boehringer Mannheim; Mannheim, Germany), FITC-conjugated anti-digoxin (clone no. DI-22; Sigma), anti-α-tubulin (Amersham; Poole, UK), and polyclonal antibody sheep antidigoxigenin-FITC (Boehringer Mannheim). Cells were incubated with 1:500 dilutions of these antibodies in TBS (150 mM NaCl, 100 mM Tris-HCl, pH 7.2) containing 0.5% (w/v) blocking reagent (Boehringer Mannheim) for 30 min. Next, cells were washed three times in TBS for 5 min each and incubated for 30 min with rabbit anti-mouse-FITC (Sigma) diluted 1:500. After three washes in TBS, slides were embedded in Vectashield (Vector Labs; Burlingame, CA) containing 4′, 6′-diamidino-2-phenyl indole (DAPI). For double staining, cells were first incubated with anti-tubulin followed by an incubation with goat anti-mouse-Cy3 (Jackson Immunoresearch; West Grove, PA). Then cells were incubated with normal mouse serum followed by an incubation with mouse anti-digoxin-FITC.
Cells were examined with a DM epifluorescence microscope (Leica) equipped with appropriate filter sets for red, green, and blue single excitation and double or triple excitation filters for multicolor exposures. Digital images were captured with a cooled CCD camera (Photometrics) and image analysis was performed on a Macintosh computer using SCIL image (Multihouse, The Netherlands).
Results and Discussion
ISH experiments designed to visualize mRNA distribution patterns in different cultured cell lines (see Materials and Methods) using digoxigenin-labeled probes and MAb anti-digoxin revealed an unexpected “specific” staining of a cytoplasmic component exclusively present in telophase cells. Figure 1 shows an example of such staining in X1 cells hybridized with a probe for β-actin mRNA. In addition to the small fluorescent spots indicating β-actin mRNA, strong staining is present at the sites where both cells are still attached to each other (Figure 1). Control experiments omitting the digoxigenin-labeled probe revealed that this strong staining was derived from the anti-digoxin-specific antibody. Furthermore, this staining pattern was not observed when the β-actin probe was labeled with biotin and detected with a streptavidin conjugate. Although anti-digoxin shows 100% crossreactivity with digoxigenin, the anti-digoxigenin-specific antibodies tested in this study did not reveal any staining of cytoplasmic components in telophase cells. When cells were hybridized first with a Cy3-labeled oligonucleotide probe specific for poly (A)+ tails or 28S rRNA sequences and then incubated with FITC-labeled anti-digoxin MAb, it appeared that the cellular component stained by this antibody did not contain RNA sequences (Figures 2a and 2b).
Because staining with anti-digoxin MAb was observed only in telophase cells at a position where both daughter cells were still connected to each other, it is most likely that the stained component is the midbody. The midbody is identified at the EM level as amorphous electron-dense matrix material and consists of microtubules and at least 35 proteins (McIntosh and Landis 1971; Mullins and McIntosh 1982). Although the midbody was discovered quite some years ago and several specific proteins have been characterized (e.g., Todorov et al. 1992; Paul and Quaroni 1993; Breckler and Burnside 1994; Baldin and Ducommun 1995; Williams et al. 1995), its function has not yet been fully elucidated. However, there are strong indications that it plays a role in cytokinesis (Williams et al. 1995) and possibly also in cell-to-cell communication (Jimenez and Goday 1993). To prove that it is indeed the midbody that stains with antidigoxin, cells were double stained with anti-digoxin and anti-α-tubulin. α-Tubulin has been identified as one of the major components of the midbody (Mullins and McIntosh 1982). Figures 3a and 3b show that the structure stained by anti-digoxin MAb is also stained intensely with anti-α-tubulin. Spindle figures in metaphase and anaphase cells also stained positively for α-tubulin but did not stain with anti-digoxin MAb, precluding crossreactivity with α-tubulin (result not shown). Apparently, this antibody shows crossreactivity with one of the other midbody's proteins.
Apart from being used in hybridization studies to detect digoxigenin-labeled probes (e.g., Huang et al. 1994; Dirks et al. 1995; Hamada et al. 1995), antidigoxin antibodies are used to measure digoxin concentrations in serum and urine of patients with cardiopathies during treatment with digitalis preparations. These measurements proved not always reliable because of the presence of endogenous digoxin-like immunoreactive factors. Some of these factors have been identified as Na+ pump inhibitors (Lichtstein et al. 1993; Tamura et al. 1994) and, in addition, immuno-cytochemical studies have demonstrated digoxin-like immunoreactivity in adrenal cortex (Ghione et al. 1993). Thus far, however, digoxin-like immunoreactivity in midbodies has not been reported.
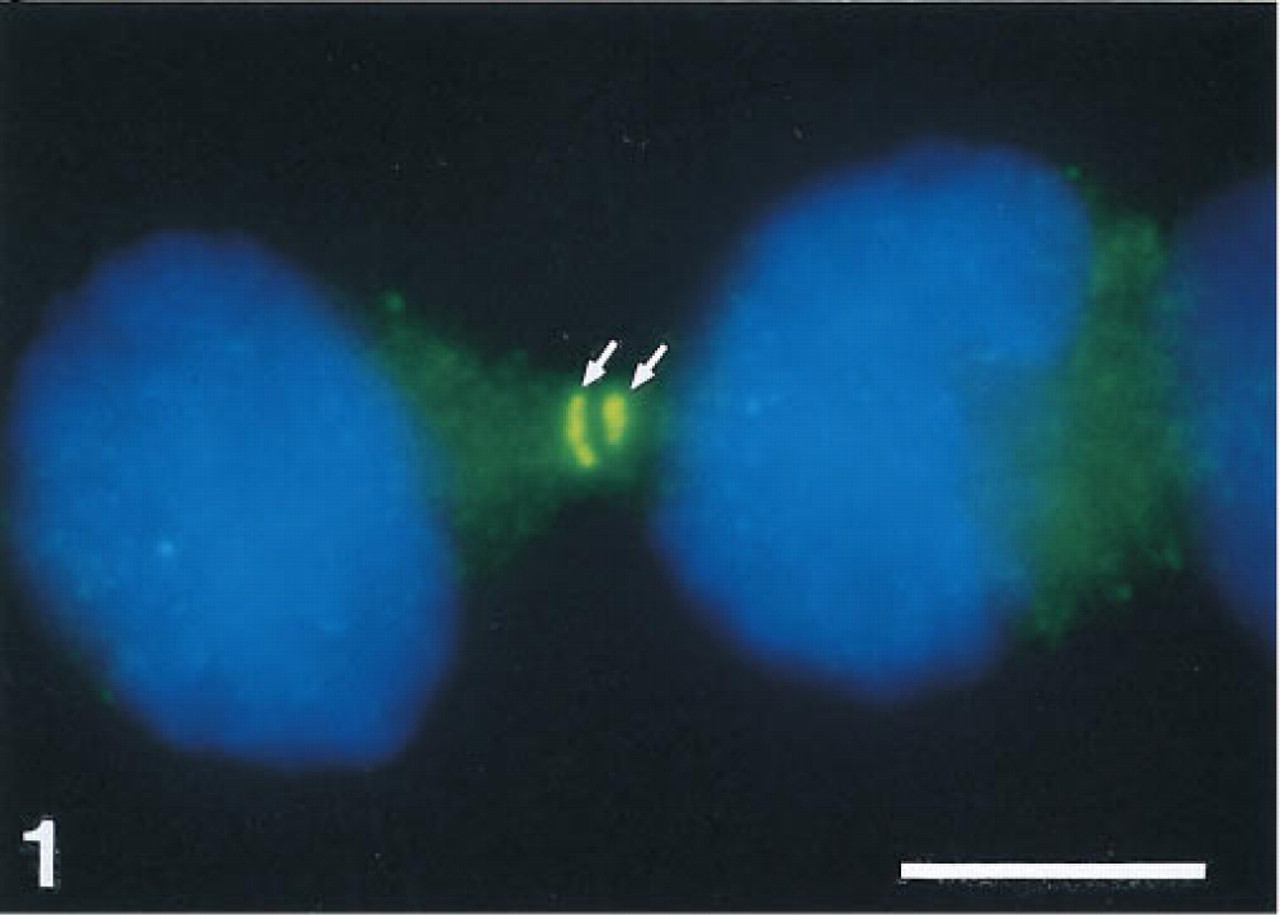
Staining of a cytoplasmic component (green, arrows) in telophase cells after RNA FISH for detection of β-actin mRNA. The small fluorescent spots throughout the cytoplasm, which are partly out of focus, indicate the presence of b-actin mRNA. X1 cells were fixed in formaldehyde/acetic acid, pretreated with pepsin, and hybridized for 3 hr with a digoxigenin-labeled plasmid probe for b-actin. Hybridized probe was detected with MAb anti-digoxin-FITC. Nuclei are counterstained with DAPI (blue). Bar 5 = μm.
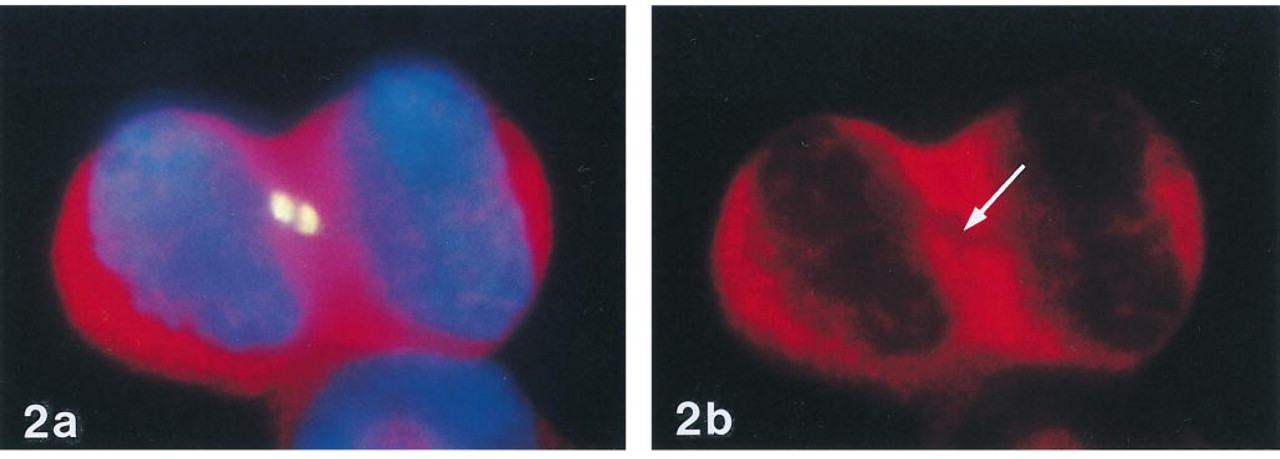
(a) Combined detection of poly (A)1 RNA by RNA FISH (red) and a cytoplasmic component by anti-digoxin-FITC (yellow) in telophase cells. (b) Absence of RNA staining at the site of anti-digoxin staining (arrow). X1 cells were first hybridized for 3 hr with Cy3-labeled oligo dT and then incubated with MAb anti-digoxin-FITC.
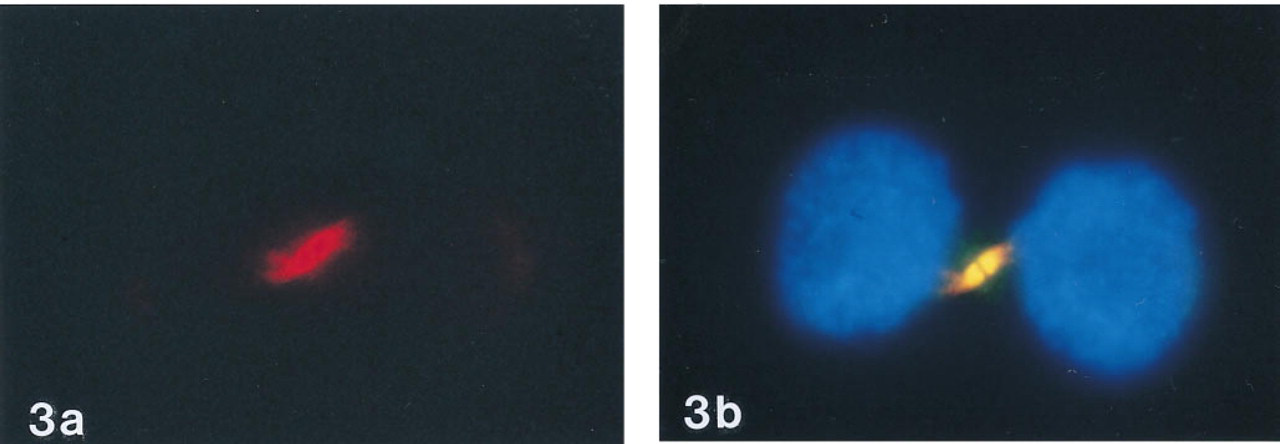
Double staining of cells with anti-α-tubulin and antidigoxin.
In conclusion, anti-digoxin MAb shows crossreactivity with one of the midbody's components. Because this crossreactivity is confined to a defined body present in telophase cells only, it will in general not trouble interpretation of ISH results. Furthermore, this crossreactivity is not observed with anti-digoxigenin-specific antibodies. Digoxigenin will therefore remain one of the most useful haptens to be employed in ISH studies.
Footnotes
Acknowledgements
Supported in part by Boehringer Mannheim.