
Abstract
Gap junctions are intercellular channels composed of connexin subunits that mediate cell-cell communication. The functions of gap junctions are believed to be associated with cell proliferation and differentiation and to be important in maintaining tissue homeostasis. We therefore investigated the expression of connexins (Cx)26 and 43, the two major connexins in human epidermis, and examined the formation of gap junctions during human fetal epidermal development. By immunofluorescence, Cx26 expression was observed between 49 and 96 days’ estimated gestational age (EGA) but was not present from 108 days’ EGA onwards. Conversely, Cx43 expression was observed from 88 days’ EGA onwards. Using electron microscopy, the typical structure of gap junctions was observed from 120 days’ EGA. The number of gap junctions increased over time and they were more common in the upper layers, within the periderm and intermediate keratinocyte layers rather than the basal layer. Immunoelectron microscopy revealed Cx43 labeling on the gap junction structures after 105 days’ EGA. Formation of gap junctions increased as skin developed, suggesting that gap junctions may play an important role in fetal skin development. Furthermore, the changing patterns of connexin expression suggest that Cx26 is important for early fetal epidermal development.
Keywords
T
Although the Cx family has more than 20 members in vertebrates, the major Cxs of the human epidermis are Cx26 and Cx43 (Salomon et al. 1994). Cx43 is expressed in the cells in the suprabasal layers and is believed to be the major gap junction subunit in the interfollicular epidermis. The expression of Cx26 is very weak in the interfollicular epidermis, but strong expression is observed in the hair follicle and sweat gland. In addition, Cx26 expression has been reported to be upregulated in the epidermis of tape-stripped skin, in hyperproliferative disorders such as psoriasis, erythrokeratodermia variabilis, verruca vulgaris (Richard et al. 1997; Labarthe et al. 1998: Lucke et al. 1999), and in cultured keratinocytes during stratification (Wiszniewski et al. 2000). Therefore, Cx26 appears to maintain keratinocyte homeostasis during rapid growth and differentiation.
It has been hypothesized that gap junctions play an important role in skin development, especially during epidermal morphogenesis. In the mouse and rat, the formation of gap junctions and the changing patterns of Cx expression have been studied in detail (Goliger and Paul 1994; Risek et al. 1994; Choudhry et al. 1997). However, little is known about Cx expression and gap junction formation during human epidermal development, although an immunofluorescent study of Cx43 expression in human fetal skin was reported recently (Hentula et al. 2001). We have examined the expression of Cx26 and Cx43 in human fetal skin using immunofluorescence and postembedding immunoelectron microscopy and have studied the ultrastructural morphology of developing human epidermal gap junctions to clarify the contribution of gap junctions during epidermal morphogensis.
Materials and Methods
Tissue
Human embryonic and fetal skin specimens were obtained from several institutes with the approval of the appropriate ethical boards and in accordance with their policies. The ages and the autopsy sites of fetuses included in the present study were as follows: 49 days’ EGA, trunk; 57 days’ EGA, trunk; 88 days’ EGA, scalp; 96 days’ EGA, trunk; 108 days’ EGA, scalp; 135 days’ EGA, scalp; 163 days’ EGA, scalp (all above for immunofluorescence); 102 days’ EGA, trunk; 108 days’ EGA, scalp; 120 days’ EGA, scalp; 152 days’ EGA, scalp (for transmission electron microscopy); 42 days’ EGA, digit and hand; 105 days’ EGA, digit and arm; 154 days’ EGA, digit and arm [for immunoelectron microscopy (McMillan and Eady 1996)]. These samples were taken from extensor or dorsal aspect of formerly mentioned regions). EGA was determined from maternal histories, fetal measurements (crown-rump and foot length), and comparative histological appearance of the epidermis (Trolle 1948; Shepard 1975; Holbrook 1979; Mercer et al. 1987).
Antibodies
The primary antibodies used in the present study were rabbit polyclonal anti-Cx26 antibody (Zymed Laboratories; San Francisco, CA) and mouse anti-Cx43 monoclonal antibody, clone 4E6.2 (Chemicon International; Temecula, CA). The secondary linker antibodies used in immunoelectron microscopy were rabbit anti-mouse immunoglobulins (DAKO; Glostrup, Denmark) and mouse anti-rabbit immunoglobulin (Jackson Immunoresearch Laboratories; West Grove, PA).
Immunofluorescent Labeling
Fetal skin was snap-frozen in isopentane and 6-μm-thick sections were cut with a cryostat. The sections were washed with 0.01 M PBS for 10 min and then incubated in primary antibody solution for 1 hr at 37C. Antibody dilutions were 1:10 for anti-Cx26 antibody and 1:100 for anti-Cx43 antibody. The sections were then incubated in fluorescein (FITC)-conjugated goat anti-mouse immunoglobulins (Jackson Immunoresearch) or in FITC conjugated to goat anti-rabbit immunoglobulins (Jackson Immunoresearch) for 30 min at room temperature, followed by 10 μ/ml propidium iodide as a nuclear counterstain (Sigma Chemical; St Louis, MO) for 10 sec. The sections were extensively washed with 0.01 M PBS between incubations. The stained sections were mounted with a glycerol-based mounting medium (Permafluor; Shandon, Pittsburgh, PA) and stored in the refrigerator. Photos were taken with an Olympus IX70 confocal laser scanning microscope. Immunostaining was detected as green (FITC) and nuclear stain was observed as red (propidium iodide). Overlap of both FITC and propidium iodide was demonstrated as a yellowish color. Image collection was performed by software Fluoview version 2.0 (Olympus America; Melville, NY). As negative control, non-immunize normal rabbit serum or mouse myeloma supernatant instead of primary antibodies was used.
Quantitative Analysis of Cx Expression in Human Fetal Epidermis
Single-channel FITC immunofluorescence images of fetal epidermis were digitized as TIFF images (using Photoshop v6.0; Adobe, CA) and subsequently analyzed quantitatively with the MCID/M2 software image analysis program (Imaging Research; Ontario, Canada). Briefly, rectangular areas containing linear epidermal basal cells up to the periderm or stratum corneum (10 cell widths on average) were selected. Image analysis was then performed to determine the overall image intensity at various levels from the base of the rectangle. Propidium iodide (PI) nuclear counterstain that was removed using Photoshop from the original image was used to determine the boundary of various layers on the MCID image analysis program.
Electron Microscopy
Fetal skin was fixed in half-strength Karnovsky's fixative or 2% glutaraldehyde solution, postfixed in 1% OsO4, dehydrated, and embedded in Epon 812 (Perry et al. 1987). All the samples were ultrathin-sectioned at a thickness of 70 nm and stained with uranyl acetate and lead citrate (Reynolds 1963). Photographs were taken with a Hitachi H-7100 or H-800 transmission electron microscope.
Cryofixed and Cryosubstituted Postembedding Immunoelectron Microscopy Without Chemical Fixatives
Postembedding immunoelectron microscopy using cryofixed and cryosubstituted skin specimens was carried out with slight modifications to the previously described method (Shimizu et al. 1989). Briefly, skin specimens were cryofixed by plunging them into liquid propane cooled to −196C, followed by cryosubstitution. They were then embedded in Lowicryl K11M (Chemische Werke Lowi; Waldkraiburg, Germany) at −60C. The specimens were polymerized by UV irradiation. Ultrathin sections were incubated for 1 hr at 37C with a primary antibody diluted 1:2 in 0.01 M PBSα. After being washed, each section was incubated for 1 hr at 37C with a secondary linker antibody. After another wash, each section was placed on a drop of 5-nm gold-labeled goat anti-mouse or rabbit IgG (British Biocell International; Cardiff, UK) diluted 1:200 at room temperature for 1 hr, and was then washed with distilled water. The sections were counterstained with 15% uranyl acetate for 2 min.
Quantitative Analysis of Gap Junctions in Fetal Epidermal Keratinocytes
Quantitative analysis of morphologically apparent gap junctions was performed within the periderm, intermediate, and basal cells. Gap junctions were counted using fetal specimens of ultrathin sections for standard high-power transmission electron microscopy. We defined a gap junction on a standard electron micrograph as a plasma membrane area in close apposition to another cell plasma membrane that exhibited a central electron-dense line between each plasma membrane lipid bilayer of adjacent cells. Gap junction counts were done on at least 26 cells in each specimen. In addition, the number of immunogold-labeled gap junctions was counted on sections stained for immunoelectron microscopy. We calculated the numbers of gap junctions per epidermal cell to evaluate the number of gap junctions in the three epidermal cell populations, i.e., basal cells, intermediate cells, and periderm cells.
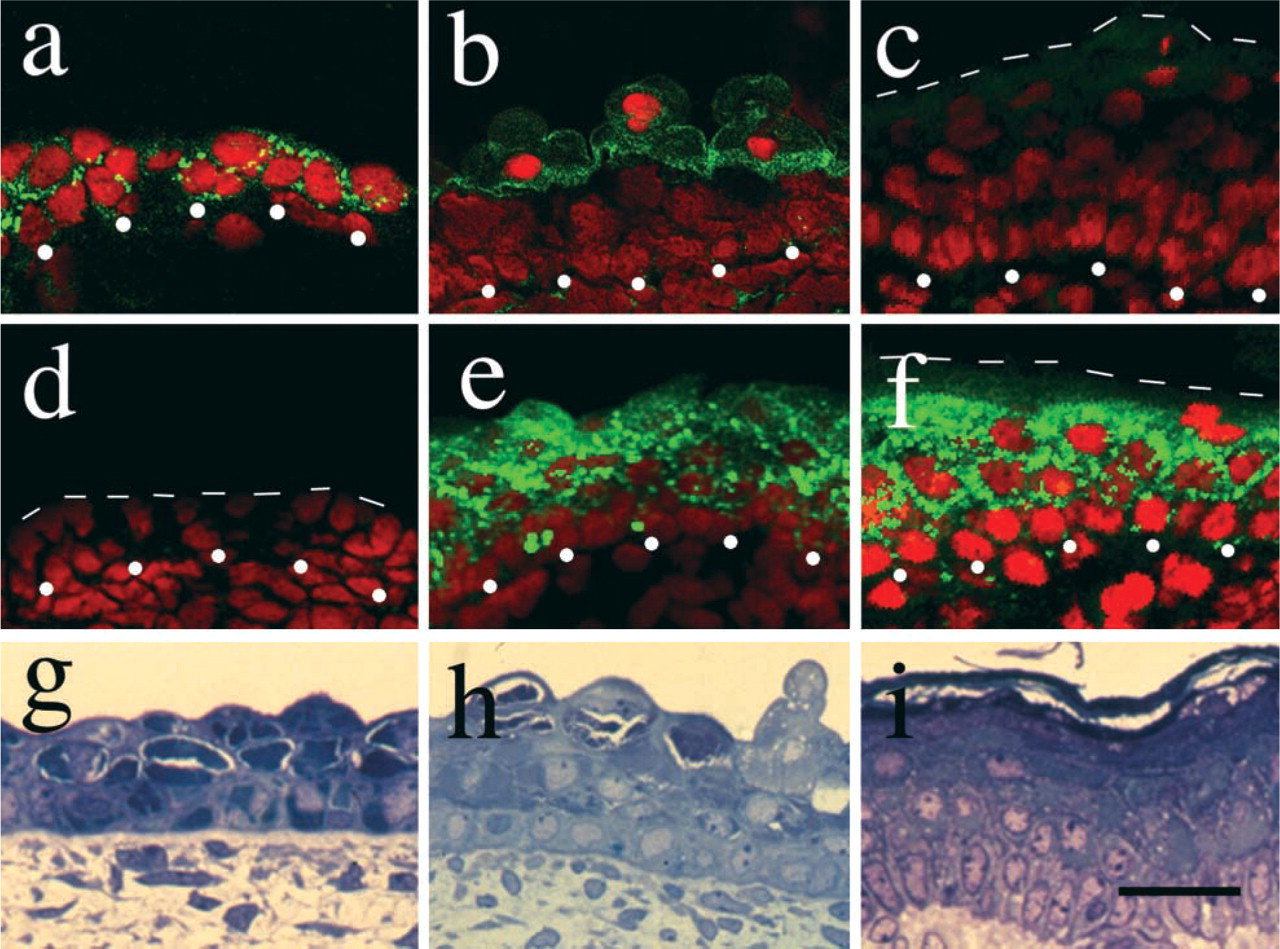
Connexin 26 was expressed in the early stages of human fetal epidermal development and Cx43 in the later stages. Cx26 was expressed in the early stages of development, but gradually disappeared as the epidermis developed. (
Results
Cx26 Is Expressed in Early Fetal Stages, Whereas Cx43 Is Expressed in Later Stages of Epidermal Development
Cx26 was expressed in the basal cells and periderm at 49 days’ EGA (corresponding to the two-layered stage) (Figure 1a), but this expression gradually became restricted to the periderm and became weaker during fetal skin development (Figure 1b). From 108 days’ EGA (four or more layered stage), expression of Cx26 was not detected in the epidermis (Figure 1c).
Conversely, the expression of Cx43 was not detected at 49 days’ EGA nor at 57 days’ EGA (Figure 1d), but from 88 days’ EGA (four or more layered stage), expression of the Cx43 molecule was detected in the intermediate and periderm layers, although the expression was very weak in the basal layer (Figure 1e). At 163 days’ EGA (the stage of interfollicular keratinization), expression of Cx43 was apparent in all the intermediate cells. Again, weak expression was seen in the basal cells and no expression was detected in the horny layer (Figure 1f). Periderm had been already been lost at this stage.
Quantitative analysis for Cx26 immunofluorescence revealed high FITC levels at the intermediate cell-periderm interface site at 96 days’ EGA (Figure 2a). However, intense FITC levels for Cx43 were noted at the intermediate cell layer at the same time point (Figure 2b).
Formation of Ultrastructurally Mature Intact Gap Junctions Is Observed After the Four or More Layered Stage of Epidermal Development
Morphologically apparent gap junction structures were not observed in the epidermis at 108 days’ EGA or younger. The intact gap junction structures were observed from 120 days’ EGA (four or more layered stage) (Figures 3a and 3b). In the epidermis at 120 days’ EGA, most gap junctions were located on the sites of periderm-periderm or periderm-intermediate cell attachment and a small number of gap junctions were seen on the intermediate cell-intermediate cell attachment region. There were no obvious gap junctions between the basal cells. At this stage of epidermal development, the numbers of ultrastructurally apparent gap junctions per periderm cell and intermediate cell were 1.8 (gap junction counts/number of periderm cells examined) and 0.15 (gap junction counts/number of intermediate cells examined), respectively (Figure 5a).
At 152 days’ EGA, most gap junctions were located between intermediate cell attachment sites and fewer numbers of gap junctions were found on any of the borders of the basal cell attachment sites. At this stage of epidermal development, the periderm had already senescenced and become detached. The numbers of apparent gap junctions per intermediate cell and basal cell were 1.0 and 0.13, respectively (Figure 5a).
Mature Intact Gap Junction Structures in Intermediate Cells of Fetal Epidermis Contained Cx43
In the epidermis at 42 days’ EGA, there were no obvious gap junction structures but sparse immunogold labeling for Cx26 was observed on the cell membrane, forming small clusters of gold particles between both basal cells and periderm cells (data not shown). Immunogold labeling for Cx43 was not seen.
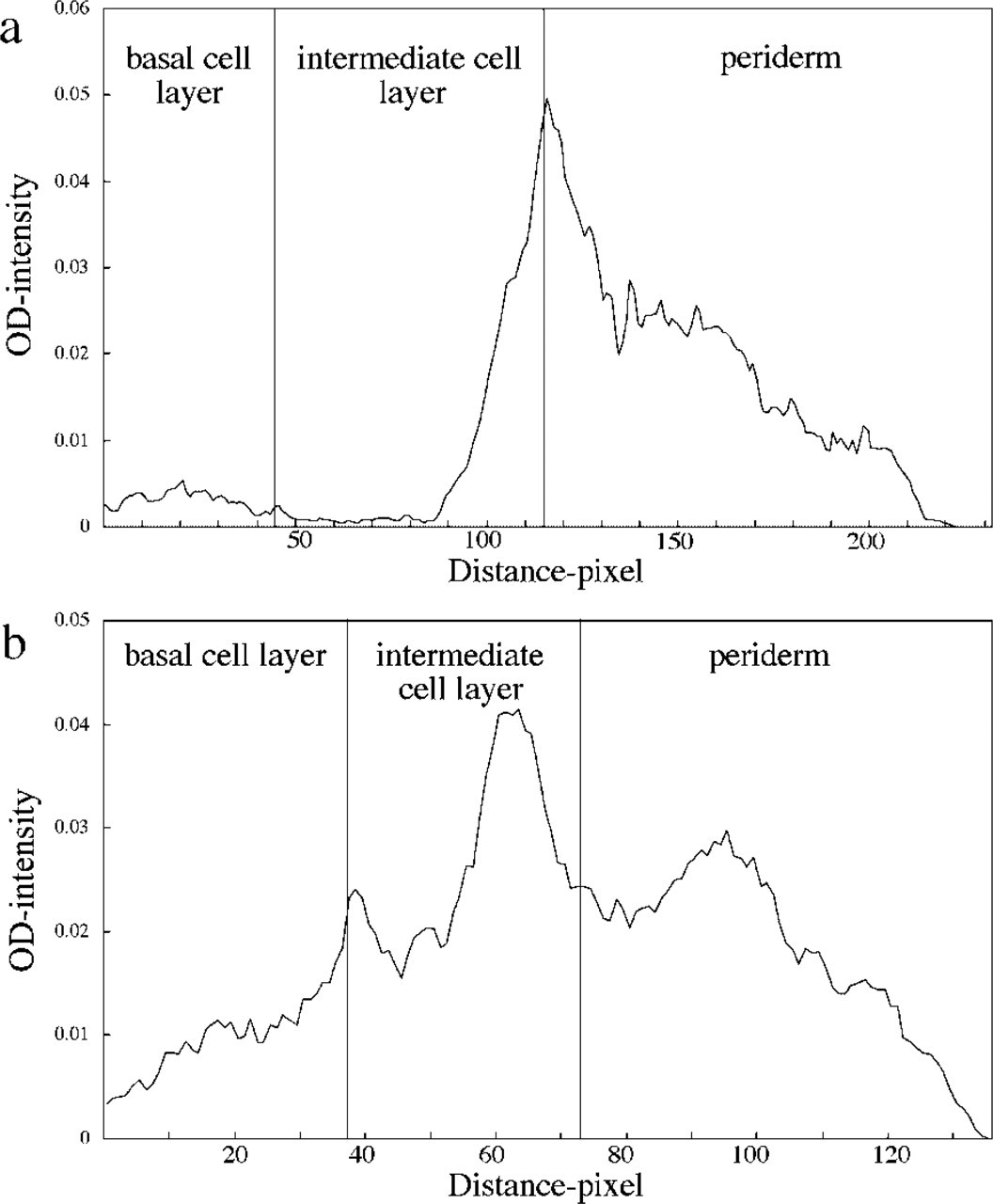
Cx26 was detected mainly within the periderm and Cx43 between the intermediate cells at 96 days’ EGA. Immunofluorescence data were analyzed with a computer to measure the distribution of Cxs in human fetal epidermis. Horizontal axis represents the distance from the basement membrane; vertical axis represents the fluorescent intensity levels. (
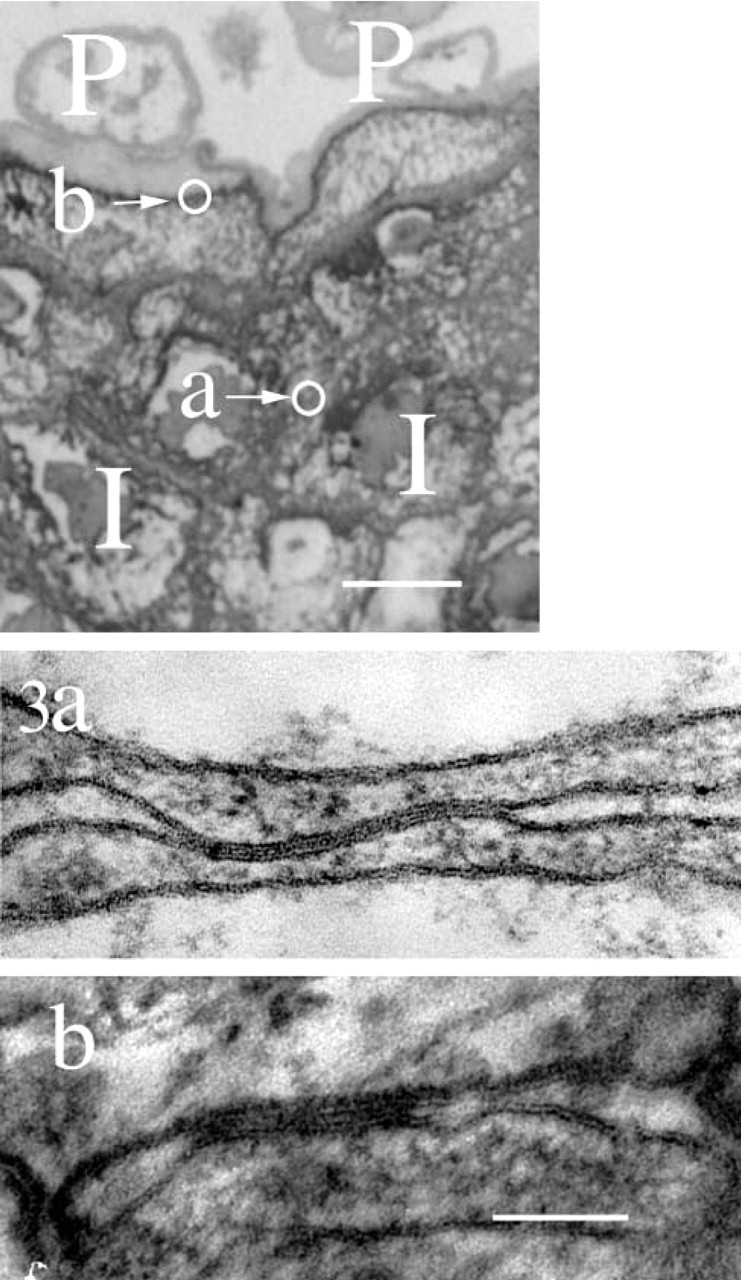
Morphologically typical gap junctions were observed from 120 days’ EGA onwards. The typical pentalaminar structures of gap junctions were observed by electron microscopy (120 days’ EGA). Highlighted on the semithin section (top) are the positions of the gap junctions shown in
At 105 days’ EGA, immunogold labeling for Cx43 was observed at immature gap junction-like structures that comprised closely apposed cell membranes of the intermediate cells (Figure 4a). The number of labeled immature gap junction-like structures per intermediate cell was 0.051 (Figure 5b). No Cx43 labeling was seen in the basal cells or the periderm. Cx26 immunogold labeling was not detected in any part of the epidermis at this stage of development.
At 154 days’ EGA, immunogold labeling for Cx43 was observed on the mature intact gap junctions on the intermediate cells (Figure 4b), and the number of labeled gap junctions per intermediate cell was 0.17 (Figure 5b). At this stage of epidermal development, no Cx43 labeling was seen in the basal cells, and the periderm had already been lost. Cx26 immunogold labeling was not detected in any layer of the epidermis.
Discussion
In the developing human fetal epidermis before interfollicular keratinization, three major populations of epidermal cells are identified: the basal cells, the intermediate cells, and the periderm. Before 36 days’ EGA, there is only a single-layered epithelium. Between 36 and 55 days’ EGA, two layers, the basal cell layer and the periderm, are observed in the primitive epidermis. Between 55 and 75 days’ EGA, the intermediate cell layer appears between the basal cell layer and the periderm and, as the fetal skin develops, the intermediate cell layer forms into two and then three or more layers. At about 160 days’ EGA, the periderm is lost and interfollicular keratinization begins (Holbrook and Odland 1975).
Cell-cell junction proteins are identifiable from the early stage of human epidermal development. For example, desmosomal proteins, the desmoplakin, desmocollin and desmoglein, and adherens junction components E-cadherin, α-catenin, and vinculin were reported to be expressed in the epidermis at 56 days’ EGA (Hentula et al. 2001). However, the exact timing of gap junction formation has not yet been clarified in developing human epidermis.
By immunofluorescence, Cx26 was expressed at 49 days’ EGA within the basal cells and periderm. However, Cx26 expression gradually disappeared and was not observed at 108 days’ EGA. Cx43 expression was observed from 88 days’ EGA. Switching of the expression pattern of the Cx family subunits was also observed during mouse embryonic development (Choudhry et al. 1997). In mouse, Cx26 and Cx43 were uniformly expressed in all epidermal layers in the early phase of development, but Cx43 expression became localized to the middle layer and Cx26 expression became restricted to the outermost differentiated epidermal layers in the later stages of development (Choudhry et al. 1997). In addition, in primary cultures of mouse keratinocytes, the induction of terminal differentiation by high calcium switching resulted in altered gap junction protein expression from Cx26 and Cx43 to Cx31 and Cx31.1 (Brissette et al. 1994). These results suggest that changing the patterns of Cxs might be closely associated with the differentiation states of epidermal keratinocytes. In human epidermis, Cx26 is upregulated in hyperproliferative conditions (Richard et al. 1997; Labarthe et al. 1998: Lucke et al. 1999). It is believed that, in early fetal development, expansion of the skin surface may need a higher proliferation rate in epidermal cells than that in the later developmental stage. The expression of Cx26 in early fetal skin may be associated with such an active epidermal proliferative status during this period.
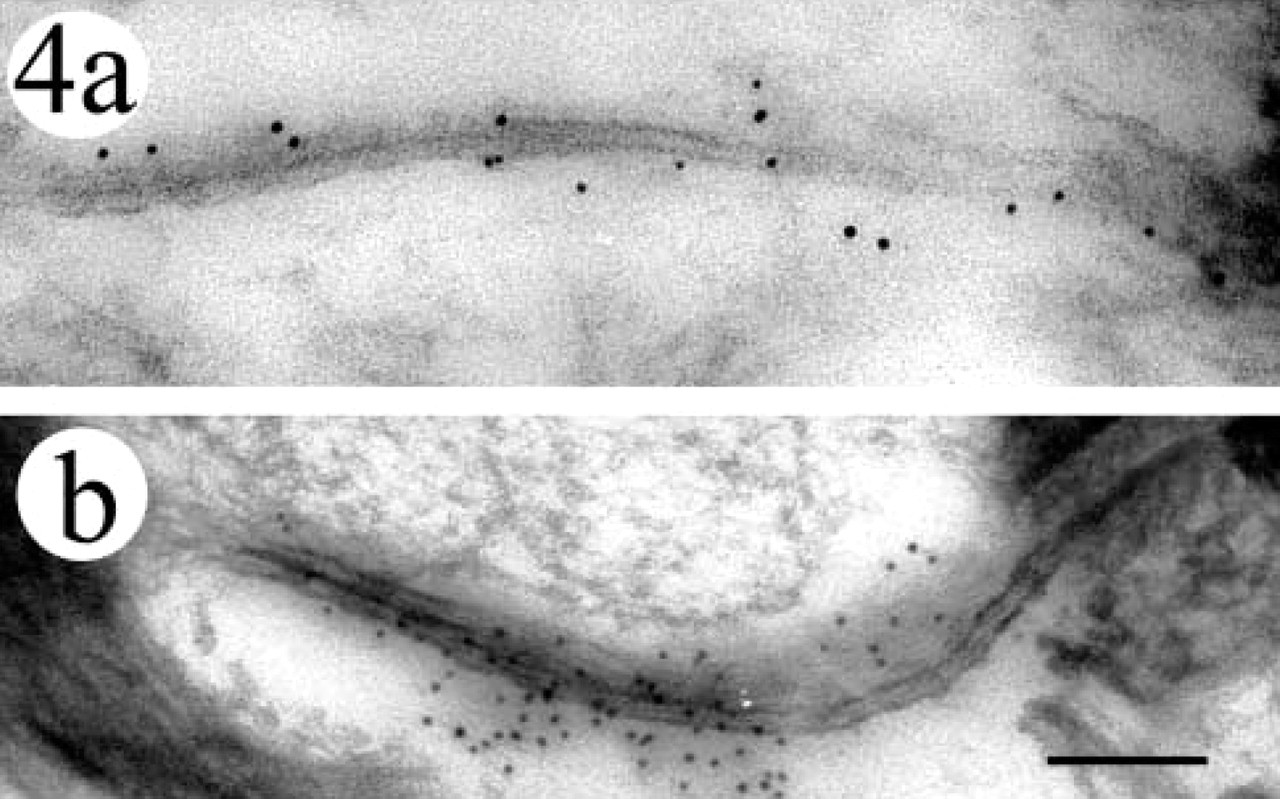
Immunogold labeling for Cx43 was observed at gap junctions in human fetal skin. (a) At 105 days’ EGA, immunogold deposition was seen at sites of immature gap junctions between the intermediate cell membranes. (b) At 154 days’ EGA, mature gap junctions labeled with Cx43 were observed at the periphery of intermediate cells. Bar = 100 nm.
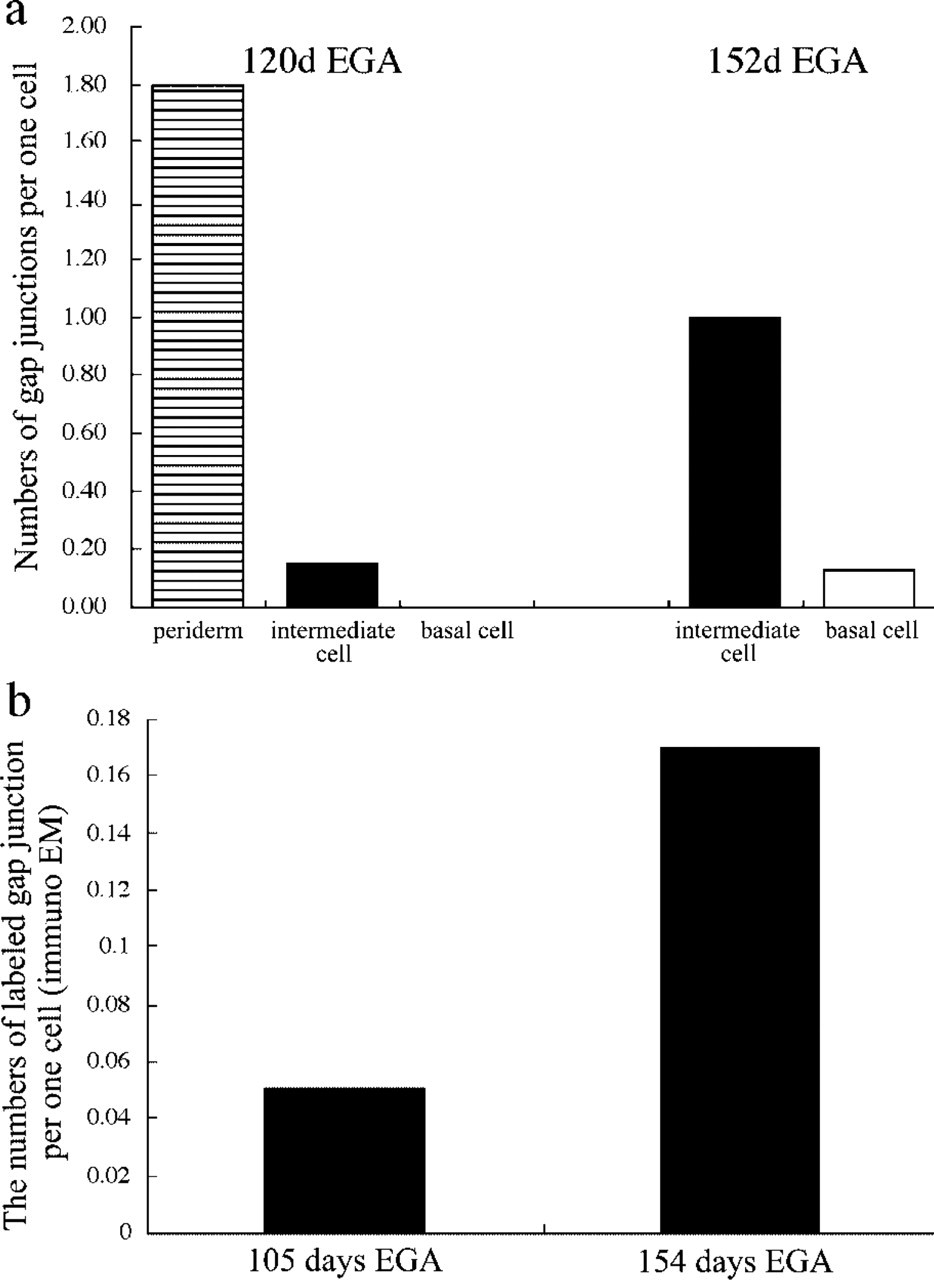
The numbers of gap junctions increased with gestational age, and gap junctions became more common in the upper layers. (a) The number of gap junctions (as observed by electron microscopy) was calculated per cell. At 120 days’ EGA, gap junctions were more common in the periderm and fewer gap junctions were seen between intermediate cells. There were no gap junctions between basal cells. At 152 days’ EGA, the periderm had already degenerated and the majority of gap junctions were between the intermediate cells. A few gap junctions were observed within the basal cells. The numbers of gap junctions in the intermediate cell and basal cell increased with gestational age. (
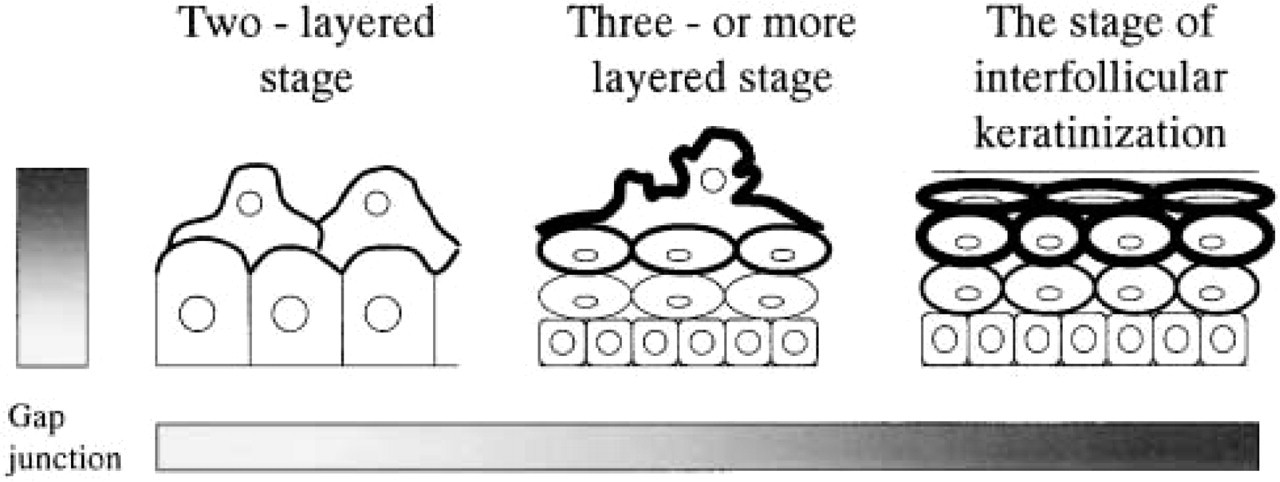
Schematic showing the formation and distribution of gap junctions during human epidermal development. More gap junctions are observed as the gestational age increases (increasing towards right), and more gap junctions are observed in the upper layer (increasing towards upper).
At 42 days’ EGA, we failed to observe mature gap junction structures, but the presence of clusters of gold particles at the cell membrane, forming immature-looking gap junction-like structures, was observed by immunoelectron microscopy. In this stage of epidermal development, Cx26 was expressed, but Cx26 molecules might only form immature gap junctions. From the four or more layered stage of epidermal development (105 days’ EGA), Cx43 staining was observed at the gap junctional structures by immunoelectron microscopy. This finding, together with the results of immunofluorescence data and electron microscopic observations, further confirms that, from the four or more layered stage, gap junctions mainly comprise Cx43. Previous studies from another group showed that the gap junction protein Cx43 was expressed on a few locations in the cell membrane of basal cells at 56 days’ EGA (Hentula et al. 2001). In our study, the Cx43 expression was observed from 88 days’ EGA. This discrepancy may be due to the different antibodies for Cx43 used and/or to the different sites of fetal epidermis sampled.
Quantitative study revealed that the number of gap junctions increased according to the maturation of the fetal epidermis. This was also highlighted by the differentiation process, in which gap junctions were more frequently observed in the intermediate cell layer than in the basal cell layer (Figure 6).
The number of gap junctions per cell was counted on ultrathin sections approximately 70 nm thick. The diameter of the keratinocyte is about 20–40 μm according to semithin section measurement. Therefore, the total number of gap junctions per single whole cell is estimated to be 300–500-fold of our data. The number of Cx43-labeled gap junctions detected in immunoelectron microscopy was about 1/6 for the data of transmission electron microscopy, as expected from the limitations of the postembedding immunoelectron microscopic technique. This is caused by immunogold particles binding only to the antigens that are exposed at the very surface of the section. The increase in Cx43-labeled gap junctions on epidermal maturation was also demonstrated by this technique. Thus, our results were confirmed not only at the ultrastructural level but also at the protein expression level. From the quantitative analysis, we can conclude that gap junctions are formed more frequently in the intermediate cells than in basal cells in accordance with keratinocyte differentiation. In contrast, in developing rat epidermis, the gap junctions were observed with the highest frequency in the basal layer at 20 days of gestation, corresponding to the period of interfollicular keratinization in rat epidermis (Risek et al. 1992). This difference may be explained by the strong expression of Cx43 in basal cells in rat fetal epidermis, which was conversely very weak in human fetal skin.
In our study, many gap junctions were observed in the periderm. The existence of gap junctions in the periderm layer was also shown in rat fetal epidermis (Risek et al. 1994). The presence of gap junctions between the periderm and the intermediate cell layer suggests active intercellular communication between these layers. The role of periderm has yet to be fully elucidated, but the presence of intercellular communication with the intermediate layer cells suggests that periderm might participate in developmental regulation and in the maintenance of homeostasis in the lower fetal epidermal layers. The periderm has abundant microvilli, which are similar to those seen in cells of the intestinal mucosa. Those morphological characteristics suggested that the periderm might have function in the uptake of carbohydrates from the amniotic fluid (Holbrook and Odland 1975). We speculate that carbohydrates are transported from the periderm to basal and/or intermediate cells via gap junctions, although experimental evidence has yet to be provided.
In conclusion, our results clearly demonstrate that Cx26 is expressed in the early stages of fetal epidermal development and Cx43 in the later stages. In addition, gap junctions are more frequently assembled as keratinocytes mature from the basal cell layer to upper epidermal layers. These facts add further evidence that the gap junctions are essential structures involved in human epidermal cell proliferation, differentiation, and morphogenesis, and suggest that Cx26 plays a crucial role in early fetal epidermal morphogenesis.
Footnotes
Acknowledgements
Supported in part by grants-in-aid for Scientific Research (nos. 13357008 and 12470175) to HS, and by no. 12670839 to MA from the Japan Society for the Promotion of Science.
We thank Prof John E. Olerud, Ms Marcia L. Usui, Mr Robert A. Underwood, Dr Lynne T. Smith, Megumi Sato, and Hideki Nakamura for their generous help in this project.