
Abstract
Myotonic dystrophy (DM) is one of the most prevalent muscular diseases in adults. The molecular basis of this autosomal disorder has been identified as the expansion of a CTG repeat in the 3′ untranslated region of a gene encoding a protein kinase (DMPK). The pathophysiology of the disease and the role of DMPK are still obscure. It has been previously demonstrated that DMPK is localized at neuromuscular junctions, myotendinous junctions, and terminal cisternae of the sarcoplasmic reticulum (SR), in the skeletal muscle, and at intercalated discs in the cardiac muscle. We report here new findings about specific localization of DMPK in the heart. Polyclonal antibodies raised against a peptide sequence of the human DMPK were used to analyze the subcellular distribution of the protein in rat papillary muscles. Confocal laser microscopy revealed a strong although discontinuous reactivity at intercalated discs, together with transverse banding on the sarcoplasm. At higher resolution with immunogold electron microscopy, we observed that DMPK is localized at the cytoplasmic surface of junctional and extended junctional sarcoplasmic reticulum, suggesting that DMPK is involved in the regulation of excitation-contraction coupling. Along the intercalated disc, DMPK was found associated with gap junctions, whereas it was absent in the two other kinds of junctional complexes (fasciae adherentes and desmosomes). Immunogold labeling of gap junction purified fractions showed that DMPK co-localized with connexin 43, the major component of this type of intercellular junctions, suggesting that DMPK plays a regulatory role in the transmission of signals between myocytes.
Keywords
M
The disease mechanism and the role of DMPK are obscure at present. In the past few years, a number of studies have been devoted to the identification of DM gene transcripts in the muscle. We and others have found that DMPK is associated with the sarcoplasmic reticulum (Salvatori et al. 1994,1997; Shimokawa et al. 1997) and is also localized to specialized cell structures in both heart and skeletal muscle. In particular, a marked concentration of DMPK has been observed at intercalated discs in cardiac muscle (van der Ven et al. 1993; Whiting et al. 1995; Salvatori et al. 1997). Intercalated discs (IDs) of mammalian heart are specialized and complex structures, the function of which is to connect the myocardial cells to each other while allowing the passage of ions and other small solutes. Over the past decade, the physiological role of ID has received much attention because morphological observations, biochemical data and, more recently, molecular biology-based approaches have made possible a comprehensive model for the ID (for review see Severs 1995).
The microscopic anatomy of IDs is complex, in that different specialized regions can be distinguished, i.e., desmosomes, fasciae adherentes, and gap junctions. Each of these regions accomplishes a different function. Fasciae adherentes and desmosomes are mainly involved in end-to-end joining and lateral adhesion of myocytes (Sommer and Jennings 1986). Gap junctions (GJs) allow an action potential to spread quickly from one cell to the other, leading to synchronized contractions of myocytes.
GJs are constituted of clusters of connexons which, in turn, are composed of transmembrane proteins called connexins. Connexins are encoded by at least 13 different genes (for review see Goodenough et al. 1996) and their distribution is largely dependent on the animal tissue and species. In the rat heart, the most represented forms are connexin 43 (Cx43) and connexin 40 (Cx40).
GJ abnormalities, such as alteration in GJ distribution and reduced synthesis of Cx43, have been reported in association with ischemic heart disease (Severs 1994). This author also hypothesizes that a more generalized Cx43 decrease per se could explain the proarrhythmic state seen, e.g., in cardiac hypertrophy. Among the several causes of arrhythmia, myotonic dystrophy (DM) must also be considered (for reviews see Cox and Kunkel 1997; Phillips and Harper 1997).
To aid in characterizing the precise sub-cellular localization of DMPK and thus contribute to the identification of its substrate(s) in tissues relevant to the disease, we focused our attention on cardiac myofibers and further analyzed the subcellular distribution of the protein by confocal laser scanning microscopy and immunoelectron microscopy. This article presents ultrastructural evidence that, in rat cardiac muscle, DMPK is distributed both at the cytoplasmic surface of sarcoplasmic reticulum elements and at gap junctions. Some of the results have been partially reported elsewhere (Mussini et al. 1996).
Materials and Methods
Animal Tissues and Gap Junction Preparation
Young adult male Wistar rats were used as a source of cardiac and skeletal muscle for morphological and biochemical studies. In particular, papillary muscles were used for ultrastructural observations. Isolated GJs were prepared from whole hearts. After tissue homogenization, Kensler and Goodenough's four-step isolation procedure (1980) was exe cuted as follows: (a) removal of cytoplasmic soluble elements and fragments of sarcoplasmic reticulum by differential centrifugation in the presence of 1 mM NaHCO3, pH 8.2; (b) removal of myofibrillar proteins by treatment with 0.6 M KI; (c) fractionation of the KI-resistant fraction and removal of fragments derived from adhesive junctions by sucrose density gradient centrifugation; and (d) solubilization of the great majority of nonjunctional membranes with Sarkosyl and Tween 20. In some experiments, partially purified GJs were further treated with 0.3% DOC (w/v) and fractioned by sucrose density gradient centrifugation. For electrophoresis and Western blotting analyses, after Sarkosyl/ Tween treatment aliquots were taken and used from resuspended pellets and from fractions either layered at the 43%/30% sucrose interface or floated at the 30% sucrose layer. Fractions layered at the 43%/30% sucrose interface both after Sarkosyl/Tween and after 0.3% DOC treatment were used for ultrastructural studies. Small aliquots were taken and centrifuged for 20 min at 105,000 X g in a high-speed Airfuge centrifuge. The pellets were then processed for immunoelectron microscopy.
Protein concentration of samples was measured by the method of Lowry et al. (1951), using bovine serum albumin (BSA) as a standard.
Antibodies
Primary antibodies used were polyclonal antibodies developed in a young female New Zealand rabbit using a synthetic 20-mer peptide chosen within the predicted DMPK sequence (IREGAPLGVHLPFVGYSYSC). Preparation of antisera and affinity purification of peptide-specific IgG were carried out according to previously reported procedures (Salvatori et al. 1997). A monoclonal antibody (MAb) specific for the cytoplasmic domain of Cx43 was also used in this study (Chemicon International; Temecula, CA).
Electrophoresis and Western Blotting
To examine the immunoreactive protein pattern of skeletal and cardiac muscles, homogenates were prepared as previously described (Salvatori et al. 1997). In brief, rat soleus and cardiac muscles were rapidly minced and homogenized in 3% SDS, 0.1 mM EGTA, pH 7.0, then boiled for 5 min before processing for gel electrophoresis (SDS-PAGE). Proteins of both homogenates and fractions of partially purified GJs were resolved by SDS-PAGE carried out according to Laemmli (1970). After electrophoretic transfer onto nitrocellulose (Towbin et al. 1979), blots were blocked in low-fat milk diluted (1:10) with TBS (10 mM Tris-HCl, pH 7.5, 0.9% NaCl) and the proteins were allowed to react with anti-DMPK antibodies (5 μg/ml) or anti-Cx43 antibodies (1 μg/ml) in TBS. The immune complexes on the blots were detected either with alkaline phosphatase-conjugated protein A followed by nitroblue tetrazolium (NBT) and 5-bromo-4-chloro-3-indolylphosphate (BCIP) for the anti-DMPK antibody or with horseradish peroxidase-conjugated anti-IgGs followed by BM chemiluminescence Western blotting reagent (Boehringer; Mannheim, Germany) for anti-Cx43 antibodies.
Immunocytochemistry
Immunofluorescence by confocal laser microscopy was performed on small bundles of fibers that were prepared from unfixed papillary muscles as previously described (Salviati et al. 1982; Salvatori et al. 1997). After preincubation with 5% goat nonimmune serum in PBS, the samples were incubated with primary antibodies (50 μg/ml in PBS), thoroughly washed in PBS, and incubated with FITC-tagged secondary antibodies. In double labeling experiments the second primary antibodies were revealed by rhodamine-labeled secondary antibodies. Nonimmune goat serum was used instead of primary antibodies in controls. The samples illuminated with an argon ion laser using the appropriate bands were observed in a Nikon RCM 8000 Real Time confocal microscope.
Electron Microscopy
Gold immunolabeling was carried out either on thin bundles of fibers taken from papillary muscles or on high-speed pellets of partially purified GJs. After a 30-min treatment with cold 0.3 M ethylacetimidate in 0.1 M phosphate buffer, pH 7.2, the samples were lightly fixed in cold 3% p-formaldehyde in the same buffer. Then the samples were thoroughly washed in PBS with the addition of 1% BSA and a cocktail of antiproteases made up of 0.2 mM 4-(2-aminoethyl)benzenesulfonil fluoride (Calbiochem; La Jolla, CA), 1 mM benzamidine, 1 mM EGTA, and 5 mM leupeptin (all obtained from Sigma; St Louis, MO). This medium was also used for dilution of antibodies. After a preincubation with 5% nonimmune goat serum, the samples were overlain (overnight at 4C) with primary antibodies which were detected with anti-rabbit or with anti-mouse IgG-gold conjugates (EM Grade; BioCell, Cardiff, UK). All incubations were followed by a number of washes (PBS with added BSA and anti-proteases) which lasted from 4 to 7 hr. In controls, primary antibodies were omitted. Double labeling experiments were carried out on high-speed pellets of partially purified GJs. After incubation with primary antibodies, the samples were incubated with biotinylated secondary antibodies which were detected with 15-nm streptavidin-gold or anti-biotin-gold conjugates (EM grade; BioCell). Then the samples were overlain (overnight at 4C) with second primary antibodies which were revealed using 5-nm anti-rabbit or anti-mouse-gold conjugates (EM grade; BioCell).
Finally, the samples were fixed with 2% glutaraldehyde in phosphate buffer (0.1 M, pH 7.2), postfixed in 1% osmium tetroxide in the same buffer, dehydrated, and embedded in Epon. Ultrathin plastic sections stained with uranyl acetate and lead citrate were observed in a Philips EM 301 electron microscope.
Results
Immunofluorescence Studies
Thin fascicles of fibers obtained from rat papillary muscles were incubated with anti-DMPK antibodies and studied by confocal laser scanning microscopy. As already shown in a previous report (Salvatori et al. 1997), in the sarcoplasm a transverse pattern of immu nofluorescence was visible, regularly spaced, and punctate in appearance. The most intense immunostaining, however, was found at the intercalated discs, as expected (van der Ven et al. 1993; Whiting et al. 1995), whereas no reactivity was present at the periphery of the fibers. When the laser source intensity was decreased to low levels (Figure 1A) the strong and continuous reactivity at intercalated discs became fragmented and appeared as discrete patches very similar to those observed after treatment with anti-Cx43 antibodies (Figure 1B). Simultaneous immunostaining of the two proteins demonstrated that they exactly co-localize at intercalated discs (Figure 1C).
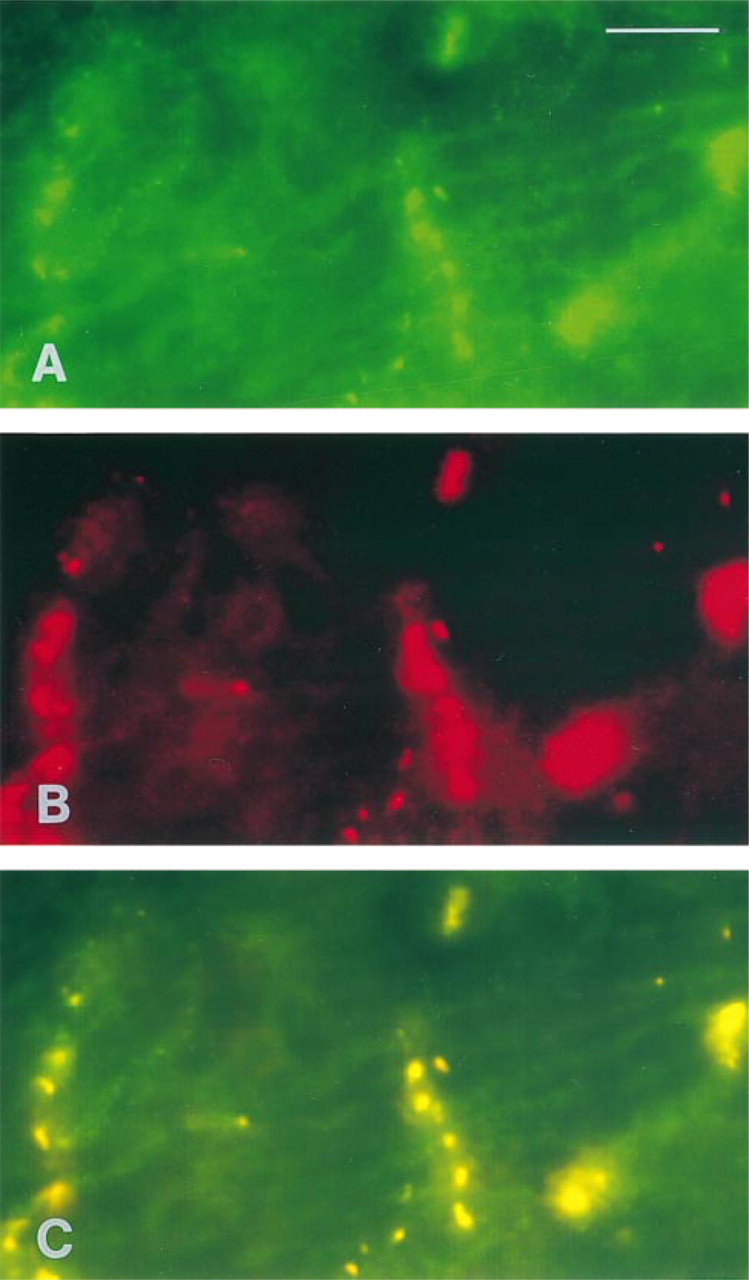
Double immunofluorescence labeling of DMPK (A) and connexin 43 (B) on adult rat papillary muscles. (C) Simultaneous double immunofluorescence picture of DMPK/Cx43 from the same area shown in A and B. The most intense immunostaining both in A and B corresponds to intercalated discs. The picture in C demonstrates that the DMPK and Cx43 co-localize. Bar = 10 μm.
Immunoelectron Microscopic Localization of DMPK in Cardiac Tissue
Immunogold electron microscopy allowed a high-resolution analysis of DMPK distribution. In papillary muscles, a large amount of protein was found associated with sarcoplasmic reticulum elements, whereas the plasma membrane and the nuclear membrane were devoid of protein. In mammalian ventricular muscle, the organization of the sarcoplasmic reticulum (SR) is characterized by the presence of a single network of tubules surrounding the myofibrils and in register with the sarcomere, whereas T-tubules mostly run transversely to the longitudinal axis of the fiber at the Z-line level. DMPK immunogold particles were recognized around the network of thin longitudinal tubules but they were particularly abundant and regularly distributed around expanded portions of SR containing electron-dense material (Figure 2). Such vesicular structures were numerous and were observed either in connection with longitudinal tubules (Figure 2A) or in close proximity to and/or apposition with T-tubules (Figures 2B and 2C), but also at the periphery of the fibers in subsarcolemmal accumulations of membranous structures (Figure 2D).
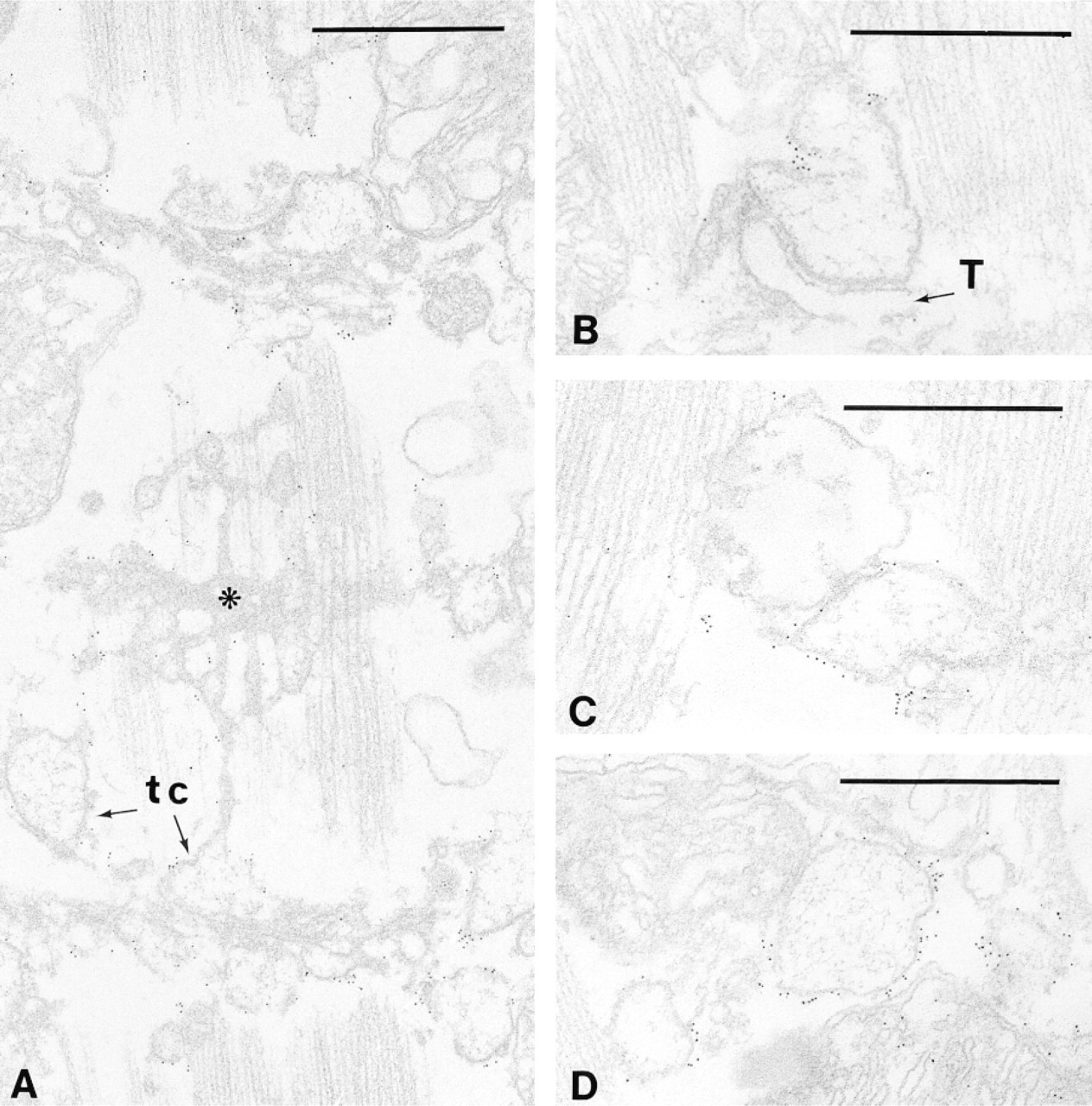
Immunoelectron localization of DMPK in papillary muscles. (A) A face view of the sarcoplasmic reticulum system is shown in which longitudinal tubules, terminal cisternae (tc), and M-rete (asterisk) are recognizable. Immunogold particles are distributed all along the surface of the SR and are mainly concentrated at junctional (B,C) and peripheral SR (D). T, T-tubules. Bars = 0.5 μm.
In the cytoplasm of the myocardiocytes, the only other localization of DMPK was found at GJs (Figures 3A and 3B) in the intercalated disc, where Cx43 is known to be present (Figure 3C). The distribution of the protein kinase was strictly limited to the GJs and was never observed at either fasciae adherentes or desmosomes (Figure 3D).
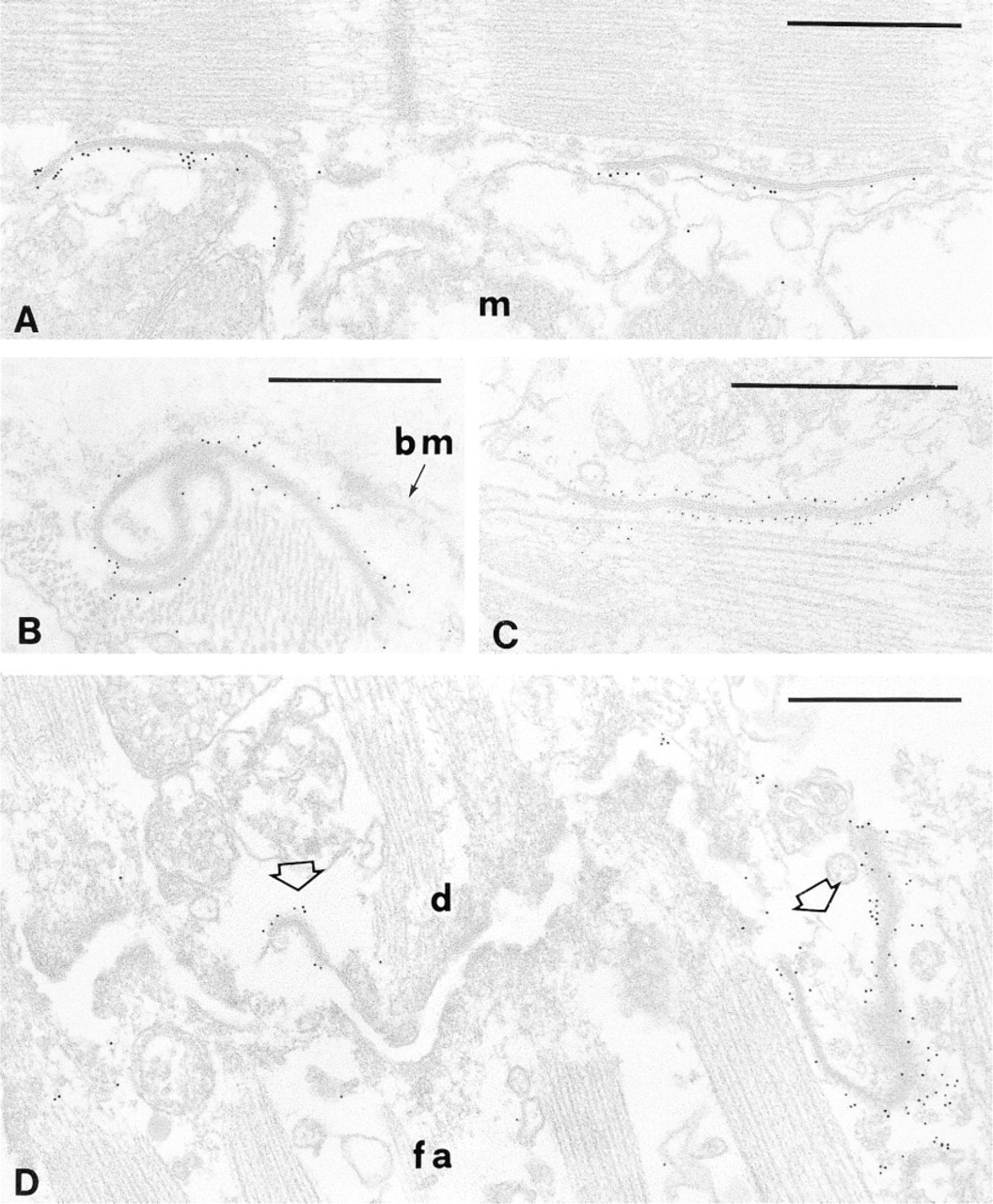
Immunoelectron localization of DMPK in papillary muscles. In ultrathin sections gap junctions are easily identifiable as double-membrane profiles (A-C). (D) The characteristic step-like structure of the intercalated disc is illustrated. Two gap junctions (open arrows) are separated by a long fascia adherens (fa); a desmosome (d) is also present. Anti-DMPK antibodies demonstrate that only gap junctions are strongly labeled with colloidal gold particles (A,B,D). The presence of Cx43 at gap junctions is shown in C. bm, basement membrane; m, mitochondrion. Bars = 0.5 μm.
To better analyze such a specific localization, we studied the presence of DMPK in partially purified fractions of GJs isolated from whole rat hearts. Immunogold staining confirmed that the protein exists at GJs (Figures 4A and 4B), where it also co-localizes with Cx43 (Figures 4D and 4E). Moreover, DMPK was still demonstrable in the gap junction after treatment with 0.3% DOC, pH 10 (Figures 4G and 4H), and its distribution remained comparable to that of Cx43 (Figure 4F).
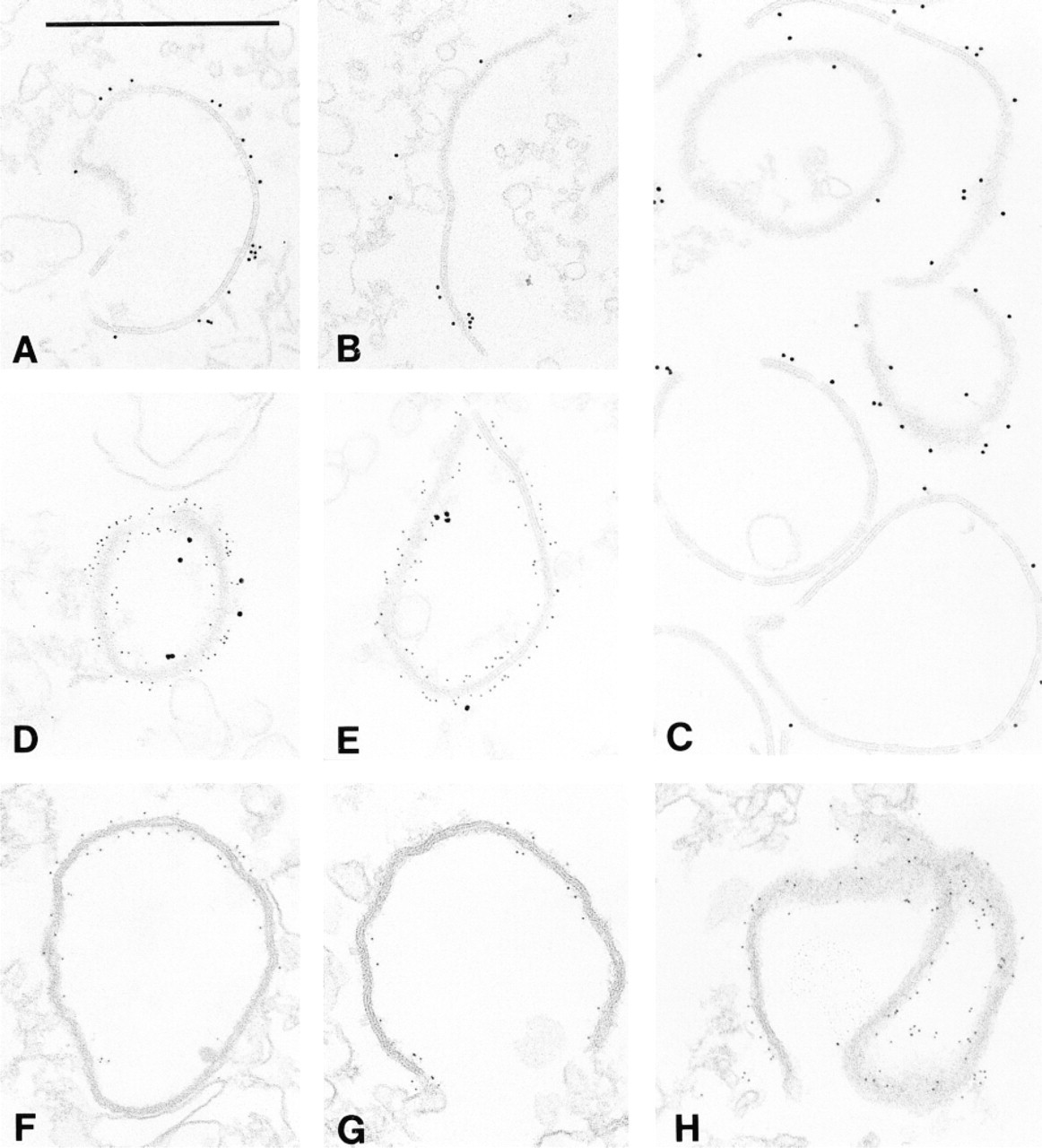
Immunogold labeling of cardiac gap junctions demonstrating the co-presence of DMPK and Cx43. Electron micrographs were taken from ultrathin sections of high-speed pellets of partially purified junctions before (A-E) and after treatment with 0.3% DOC (F-H). Gap junction preparations were treated with anti-DMPK (A,B,G,H) or with anti-Cx43 (C,F) antibodies. By double staining, DMPK (large gold particles) and Cx43 (small particles) are simultaneously demonstrated on gap junctions (D,E). Bar = 0.5 μm.
The amount of protein kinase revealed by immunogold staining along single junctions varied from large (Figure 4A) to small, or was seen in a limited part of the structure (Figure 4B). Very likely such a variable distribution of DMPK was mainly due to technical reasons or to steric hindrance of gold immunoconjugates. The same variability was seen when anti-Cx43 antibodies were used (Figure 4C). On the other hand, the density of particles was much higher when 5-nm immunogold (Figures 4G and 4H) was used but was very low with 15-nm immunogold conjugates (Figures 4D and 4E).
Western Immunoblot Analyses
Muscle homogenates obtained both from soleus and cardiac muscles were analyzed by Western blotting. As already shown (Salvatori et al. 1994,1997), in slow skeletal and in cardiac muscle two proteins of 85 kD and 54 kD, respectively, were immunostained by our polyclonal anti-DMPK antibodies, the latter being more abundant in cardiac than in skeletal muscle (Figure 5, Lanes 1 and 2).
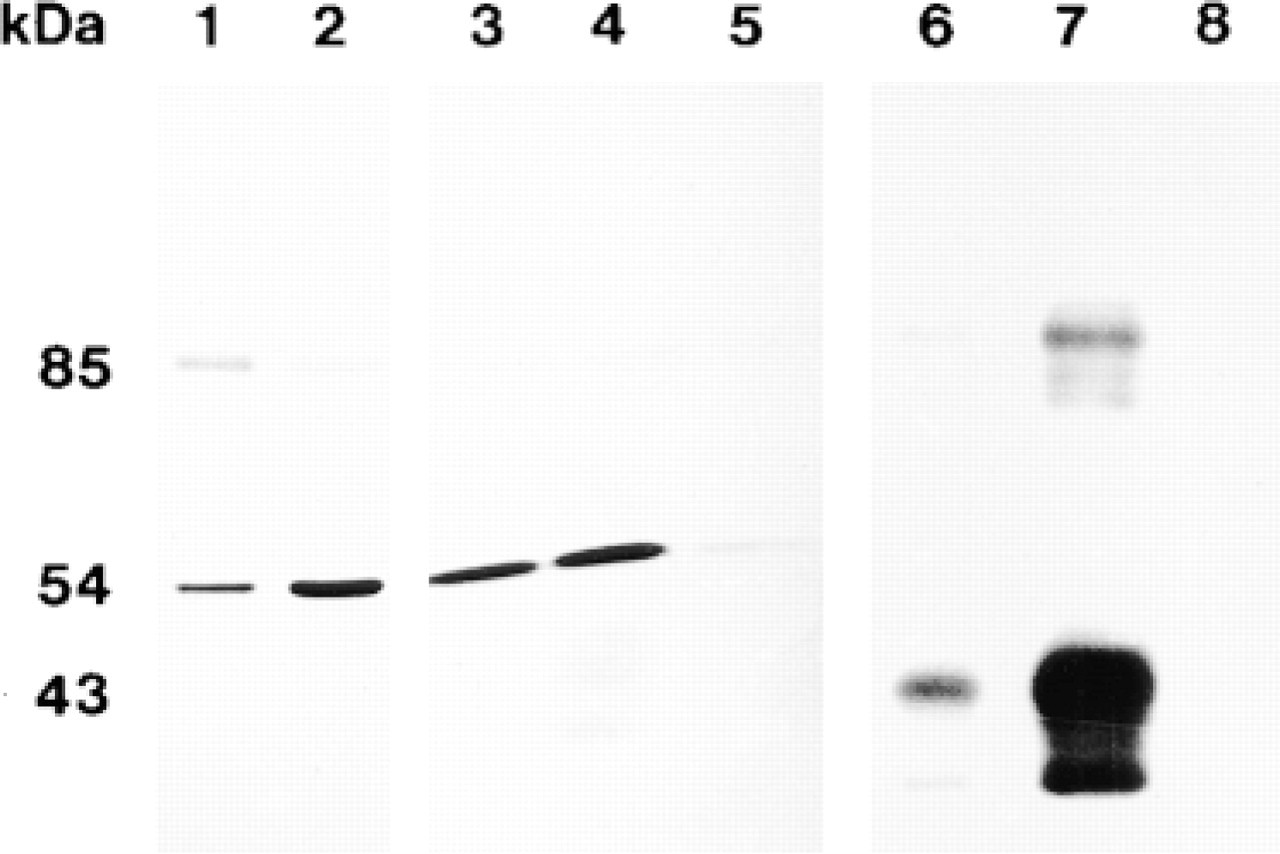
Western immunoblot analysis of rat muscle tissue homogenates and fractions enriched in cardiac gap junctions. Muscle homogenates (Lanes 1 and 2) were prepared as previously described (Salvatori et al. 1997). Detergent-resistant gap junctions (Lanes 3-5 and 6-8) were isolated according to Kensler and Goodenough (1980). Lane 1, soleus muscle homogenate; Lane 2, cardiac muscle homogenate; lanes 3, and 6, pellet after Sarkosyl/Tween treatment; Lanes 4 and 7, fractions layered at the 43%/30% sucrose interface; Lanes 5 and 8, fractions floating at the 30% sucrose layer. Samples were loaded as follows: Lanes 1 and 2, 50 μg; Lanes 3-5, 40 μg; Lanes 6-8, 10 μg. Samples in Lanes 1-5 were allowed to react with anti-DMPK antibodies (5 μg/ml), incubated with alkaline phosphatase-conjugated protein A; the immunoreactive proteins were then revealed with NBT/BCIP developing system (Sigma). Samples in Lanes 6-8 were reacted with anti-Cx43 (Chemicon) (1 μg/ml), incubated with horseradish peroxidase-conjugated antimouse IgGs, and the immunocomplexes were revealed with BM chemiluminescence blotting substrate (Boehringer).
Partially purified GJs from ventricular muscle exhibited strong immunoreactivity corresponding to the 54-kD band (Figure 5, Lanes 3 and 4). In the same fractions, Cx43 was revealed by monoclonal anti-Cx43 antibodies (Figure 5, Lanes 6 and 7). In contrast, a DMPK content was barely visible and Cx43 was completely absent in fractions floating at the 30% sucrose layer (Figure 5, Lanes 5 and 8), where only empty vescicular structures were found (results not shown). These biochemical results confirmed that Cx43 and DMPK co-localize in the same structures, i.e., cardiac gap junctions, and that such an association is resistant to detergent treatment.
Discussion
The data presented here demonstrate that DMPK, the product of the DM gene, in rat cardiac muscle is associated with precise and highly specialized subcellular structures, i.e., junctional SR and GJs.
The immunofluorescence observations parallel the findings of Whiting et al. (1995), who described intense staining at intercalated discs. On the other hand, Salvatori et al. (1997) described immunoreactivity in the form of transverse banding and intense staining at intercalated discs of rat cardiac myofibers. Later, Ueda et al. (1998) reported a similar immunolocalization of DMPK in human cardiac muscle. In skeletal muscle, the specific localization of DMPK protein to the SR terminal cisternae has been demonstrated by our recent studies (Salvatori et al. 1997) and was shown by electron microscopic observations of Shimokawa et al. (1997).
The results presented here on immunogold labeling (Figure 2) provide for the first time in rat papillary muscles a precise identification at the ultrastructural level of SR elements with which DMPK is associated, i.e., junctional and extended junctional SR. As in skeletal muscles, two components of sarcoplasmic reticulum can be distinguished in mammalian ventricular muscle, i.e., junctional and nonjunctional SR. The former, however, is more complicated in cardiac than in skeletal muscle. In addition to terminal cisternae proper, which make direct contact with T-tubules, two other forms have been included in the junctional types of SR: corbular and peripheral SR. Corbular SR is preferentially seen near the Z-band, whereas peripheral SR forms a multilayered network in subsarcolemmal spaces (Ogata and Yamasaki 1990). The two forms, i.e., corbular and peripheral SR, are called extended junctional SR (EJSR) because they are structurally identical to JSR. They contain electron-dense material in the lumen and junctional processes on their surfaces (Dolber and Sommer 1984; Sommer and Jennings 1986). On the basis of our results, we can therefore conclude that in cardiac muscle DMPK is distributed on the cytoplasmic surface of junctional SR. Very recently, similar conclusions were also drawn by Ueda et al. (1998), based on their ultrastructural observations in ventricular muscle taken from autopsied human hearts. They found strong immunoreactive products of DMPK along the Z-band, as already reported by Mussini et al. (1996) in rat ventricular muscle. The findings presented here definitively confirm and extend previous observations of Salvatori et al. (1994, 1997), who showed that in skeletal muscles DMPK is mainly associated with purified membrane fractions corresponding to SR terminal cisternae. The possibility that junctional proteins, such as the ryanodine receptor or triadin, could be targets for DMPK cannot be ruled out. On the other hand, Damiani et al. (1995) have shown that histidine-rich Ca-binding protein (HCP) and triadin are phosphorylated by Ca/Calmodulin-dependent protein kinase II in vivo and by other protein kinases in vitro. On this basis, it can be suggested that HCP is also a substrate candidate for DMPK activity in junctional SR. Likewise noteworthy, the gene for HCP is located close to that for DMPK, suggesting a possible coordinated expression of the two proteins. We have already reported (Salvatori et al. 1997) that the immunofluorescence at intercalated discs (IDs) had a punctate appearance. This suggested that DMPK could be not homogeneously distributed along ID. Such a hypothesis was at first verified by a more accurate analysis with confocal laser microscopy. Low levels of laser source intensity applied to thin samples revealed a very discontinuous positive immunoreactivity for DMPK, overlapping that for Cx43 (Figure 1).
In human cardiac myofibers, DMPK has been localized at various distances from the intercalated discs (Ueda et al. 1998). Immunogold results presented here for the first time demonstrate that along the intercalated disc DMPK is exclusively localized at the GJs (Figure 3). By the use of our polyclonal antibodies, DMPK is revealed on the cytoplasmic face of the junction (Figures 3A and 3B) with a distribution similar to that of Cx43 (Figure 3C). Moreover, DMPK is absent in the other specialized regions of the ID, i.e., at desmosomes and at fasciae adherentes (Figure 3D).
In the past few years, several reports have shed light on the protein composition of ID and the different specific proteins found in its different regions. Desmosomes are characterized by the presence of desmin filaments, desmogleins and desmocollins (Buxton et al. 1993). The fascia adherens is mainly composed of classical cadherins (Kemler 1993; Angst et al. 1995). In addition, as reported by Kijma et al. (1993), the IP3 receptor, a type of intracellular calcium release channel, is localized at the same adhesive junction. Very recently, Cartegni et al. (1997) reported that emerin, the gene product of the pathological gene inherited with Emery-Dreifuss disease, is localized both to desmosome and fascia adherens but not to GJs. Finally, GJs are built up of connexons, the intercellular channels formed from connexins (for review see Goodenough et al. 1996). We have shown here that in rat ventricular muscle DMPK, if not a component of GJ, is at least tightly and firmly bound to it. Immunogold staining for DMPK of GJ fractions demonstrated that after isolation and purification treatments the protein was still present on the junctions and with a distribution similar to that of Cx43. In addition, the two proteins were shown to co-localize (Figure 4). These observations were then confirmed by Western blot analyses of the same fractions (Figure 5).
Since the first report on Cx32 phosphorylation in hepatocytes (Saez et al. 1986), it has become clear that all connexins are phosphoproteins except for Cx26. Connexin phosphorylation has been linked to several physiological events such as assembly, docking, and gating of the channel. It occurs prevalently at Ser residues in the C-terminal cytoplasmic tail of the proteins. In the presence of a high concentration of cAMP, permeability or pH-dependent gating of connexons is apparently not affected (Saez et al. 1990), suggesting that PKA-dependent phosphorylation regulates other functions. Musil and Goodenough (1991) reported that in vivo Cx43 is phosphorylated after connexon assembly and surface localization, thus suggesting that post-translational addition of phosphate may regulate later events. Moreover, by recording single-channel activity of hepatoma cells transfected with Cx43, Moreno et al. (1994) reported that variations of the unitary conductance of human Cx43 gap junction channels are consistent with the phosphorylation state of the protein, thus relating the Cx43 phosphorylation to the channel activity. On the other hand, Cx43 phosphorylation has been associated not only with the gating properties of the channel but also with its correct assembly during morphogenesis (Stagg and Fletcher 1990; Godwin et al. 1993). The possibility that Cx43 may be phosphorylated by more than one protein kinase has to be taken into consideration. Given our results (Figures 3–5), we propose here that DMPK takes part in the regulatory mechanism of connexon activity. By immunofluorescence studies, Meldrum et al. (1996) reported that in ventricular sections of isolated rat hearts the Ca-induced myocardial inotropy is characterized by the translocation of PKCα from the cytoplasm to the sarcolemma and intercalated discs. It would be worthwhile to investigate in greater detail if DMPK and some of the PKC isoforms may have differential or synergistic effects on signal transduction mechanisms.
The crucial role played by the phosphorylation of Cx43 is dramatically demonstrated by a complex familial syndrome characterized by heart malformations and defects of laterality (visceroatrial heterotaxia) due to mutations at one or more phosphorylatable serine or threonine residues of the C-terminal tail of the protein (Britz-Cunningham et al. 1995). Malformations such as stenosis or atresia of the pulmonary artery were found in knockout mice lacking the Cx43 gene (Reaume et al. 1995). Several recent articles lead to the idea that Cx43 undergoes multiple phosphorylations, possibly in a hierarchical manner, which are responsible for assembly, docking and gating of the channel (see Bruzzone et al. 1996 for a detailed discussion). One can therefore speculate that stable, i.e., genetically determined, defects directly related to the Cx43 gene are mainly involved in developmental disorders, whereas more subtle defects, such as altered phosphorylation of Cx43, may be responsible for disturbances in the gating properties of the channel. As we have shown here, DMPK co-localizes with Cx43 in the GJ of adult rat cardiac muscle. The hypothesis can therefore be advanced that DMPK is involved in regulation of the connexon activity, possibly affecting the gating of the channel and thus playing an important role in the transmission of signals between myocytes.
Mice lacking the DMPK gene (Jansen et al. 1996; Reddy et al. 1996) or overexpressing a human DMPK transgene (Jansen et al. 1996) have been generated. Surprisingly, knockout mice were found to exhibit minor changes in fiber size in old age only. Mice overexpressing DMPK occasionally show a mild hypertrophic cardiomyopathy. In both cases, however, the phenotype is normal. As discussed by the same authors, this may be due to relevant differences in physiological features of human and mouse. However, differences between the two species in body size as well as posture characteristics have to be considered, together with differences in metabolic pathways. On the other hand, the mild hypertrophy of cardiac muscle in transgenic mice is manifested only when more than 20 copies of the gene are introduced into the chromosome (Jansen et al. 1996). Indeed, it could be inferred that the overexpressing mouse is a rather unsuitable model for clarifying the multisystemic effects of the triplet repeat expansion in humans. In humans, cardiac involvement is a common and frequent cause of death in myotonic dystrophy and is not confined to severely disabled patients. Interestingly, the gene for progressive familial heart block Type 1 has been mapped close to the DM locus (Brink et al. 1995), and cardiac signs of patients suffering from this disease partially overlap those of DM patients. Unlike ischemic diseases of the heart, in which profound alterations of the connexons are morphologically evident (Severs 1995), DM cardiac disturbances are mainly related to conduction defects and arrhythmias, without gross modifications of the tissue organization (Thomson 1968).
As mentioned before, Cx43 phosphorylation has been implicated in channel formation (Musil and Goodenough 1991) and, more recently, in the rapid turnover of Cx43 as well as in the remodeling of gap junctions (Beardslee et al. 1998). It has also been suggested that such remodeling may lead to conduction discontinuity in the heart and, as a consequence of tissue anysotropy, to arrhythmia (Spach and Heidlage 1995; Spray 1998). Given our results, we conclude that DMPK, alone or together with other protein kinases, may have a relevant role in the modulation either of the functional activity of connexons or of their metabolic pathways.
Footnotes
Acknowledgements
Supported by institutional funds from Consiglio Nazionale delle Ricerche and by grant funds (to SS) from Telethon Italy (project No. 913).
The technical assistance of Mr V. Gobbo is gratefully acknowledged.