
Abstract
Dopamine D4 receptors mediate inhibition of vasopressin-dependent sodium reabsorption by dopamine in collecting tubules. At present, the distribution of D4 receptors in other renal districts remains an open issue. The renal distribution of D4 receptor was assessed in normally innervated and denervated male Sprague-Dawley rats by quantitative immunohistochemistry using an anti-dopamine D4 receptor rabbit polyclonal antibody. D4 receptor protein immunoreactivity was observed perivascularly in the adventitia and the adventitia-media border. The density of perivascular dopamine D4 receptor was higher in afferent and efferent arterioles than in other segments of the renal vascular tree. Renal denervation abolished perivascular dopamine D4 receptor protein immunoreactivity. In renal tubules, the epithelium of collecting tubules showed the highest dopamine D4 receptor protein immunoreactivity, followed by the epithelium of proximal and distal tubules. No dopamine D4 receptor protein immunoreactivity was observed in the epithelium of the loop of Henle. Denervation did not change dopamine D4 receptor protein immunoreactivity in renal tubules. These results indicate that rat kidney expresses dopamine D4 receptors located both prejunctionally and nonprejunctionally in collecting, proximal, and distal tubules. This suggests that the dopamine D4 receptor may be involved in the control of neurotransmitter release and in renal hemodynamic and tubule function.
I
The D3 receptor is the renal dopamine D2-like receptor subtype most investigated. It was characterized by reverse transcription-polymerase chain reaction (RT-PCR) (Gao et al. 1994), radioligand binding assay, and autoradiography (Barili et al. 1997a) as well as by immunohistochemistry and electron microscopic immunocytochemistry (O'Connel et al. 1998). This receptor has a rather widespread distribution, being located in the renal vasculature, glomeruli, and in the proximal, distal, and collecting tubules (O'Connel et al. 1998). Information on the renal dopamine D4 receptor is less extensive. Recent investigations have reported the expression of dopamine D4 receptors in the kidney by molecular biology (Matsumoto et al. 1995; Sun et al. 1998), radioligand binding assay (Ricci et al. 1997), immunohistochemistry (Sun et al. 1998; Schafer et al. 2000), and functional approaches (Sun and Schafer 1996; Schafer et al. 2000). Studies in rat indicated that D4 receptors are located almost exclusively in collecting tubules, where they are involved in inhibition of vasopressin-dependent sodium reabsorption (Sun et al. 1998; Schafer et al. 2000). The observation that D4 receptors are also present in glomerular arterioles (Amenta 1997; Sanada et al. 1997; O'Connel et al. 1998) suggests that these receptors may have a wider distribution in kidney. To date, the potential localization of D4 receptor in renal prejunctional endings has not been reported. However, this possibility is supported by a recent study showing the presynaptic localization of D4 receptors in rat brain (Svingos et al. 2000).
To define the post- and prejunctional localization of renal D4 receptors, we investigated their microanatomic localization in kidneys of sham-operated and denervated rats.
Materials and Methods
Animals and Tissue Treatment
Male Sprague-Dawley rats (n=20; bw ∼290 g) were used. Ten animals were anesthetized with an IP injection of pentobarbital sodium (60 mg/kg) and laparotomized. The right renal artery was exposed and denervated by stripping the artery surgically and swabbing it with a 10% phenol solution dissolved in ethanol according to the procedure detailed elsewhere (Lakhlani et al. 1994). The remaining rats were sham-operated. One week after surgery, five sham-operated rats and five denervated animals were anesthetized as described above and laparotomized. Right (denervated) and left (non-denervated) kidneys were removed, washed in an ice-cold 0.9% NaCl solution, and divided into two halves (upper and lower half). The upper half was weighed and used for norepinephrine (NE) assay and the lower half for Western blotting analysis.
The remaining rats were perfused through the abdominal aorta with a 0.9% NaCl solution containing heparin (20 IU/100 ml). This solution was replaced by a fixative solution of a 4% freshly prepared paraformaldehyde and 0.4% picric acid in phosphate buffer (pH 6.9). At the end of perfusion, kidneys were removed, cut into slices of 0.2–0.5 cm, and put in the same perfusion for an additional 72 hr. At the end of fixation, slices were washed, dehydrated in ethanol, and embedded in a semi-synthetic paraffin. Paraffin blocks were cut serially using a motorized microtome and mounted on gelatin-coated microtome slides. Each slide contained two 10-μm-thick sections of kidney.
NE Assay
The upper halves of the right and left kidneys were homogenized in ice-cold 0.1 M HClO4 (5 ml/g tissue). The homogenate was centrifuged at 2000 × g for 20 min at 4C and the resulting supernatant was used for NE assay. NE was assayed by high-pressure liquid chromatography (HPLC) with electrochemical detection (Lakhlani et al. 1994).
Western Blotting Analysis
The upper halves of the right and left kidneys were homogenized in 0.32 M sucrose. The homogenate was centrifuged at 2000 × g for 20 min at 4C to remove nuclei and cell debris, and the resulting supernatant was centrifuged at 14,000 × g for 30 min. The pellet representing kidney membranes was re-suspended in a buffer containing phenylmethyl sulfonylfluoride, aprotinin, and leupeptin. Equivalent amounts of proteins were separated by 10% SDS-PAGE and transferred to nitrocellulose. Anti-dopamine D4 receptor protein antibody was dissolved in 0.1 M PBS containing BSA (1%) and Tween-20 (0.05%) at a dilution of 1:400. This antibody concentration was established in a series of preliminary experiments. The specificity of the immune reaction was assessed using an antibody preadsorbed with the peptide used for generating it. A 0.05% 3–3′-diaminobenzidine tetrahydrochloride (DAB) solution was used as a chromogen for the peroxidase reaction.
Dopamine D4 Receptor Protein Immunohistochemistry
Groups of six consecutive slides were used. The first section was stained with hematoxylin-eosin to verify microanatomic details. The second and fourth sections were exposed to the primary anti-dopamine receptor antibody in phosphate buffer containing 0.2% (w/v) BSA, 0.03% Triton X-100, and 0.1% (w/v) sodium azide, diluted 1:2500. The third and fifth sections were incubated as above with the primary antibody pre-adsorbed with its peptide antigen at a dilution of 10 μg/ml. The sixth section was exposed to a nonimmune serum instead of the primary antiserum. Incubation with antibody was accomplished in a humid chamber at 4C for 12–18 hr. Optimal antisera dilutions and incubation times were assessed in a series of preliminary experiments.
After incubation, slides were rinsed twice in phosphate buffer and exposed for 30 min at 25C to a biotinylated anti-rabbit secondary antibody at a dilution of 1:200. The product of the immune reaction was then revealed using a biotin-streptavidin immunostaining kit with 0.05% DAB in 0.1% H2O2 as a chromogen. Sections were then washed, dehydrated in ethanol, and mounted in a synthetic mounting medium.
Microdensitometry
The intensity of immunoreactivity developed in the epithelia of the proximal tubule, loop of Henle, and distal and collecting tubules was assessed microdensitometrically. For microdensitometry, six sections of right and left kidneys in five denervated and sham-operated rats were examined at a final X400 magnification. Fifty proximal, distal, and collecting tubules or loop of Henle epithelial cells per section were identified and delineated by a measuring diaphragm whose size was adapted to cover one cell at a time. In parallel sections, proximal tubules were distinguished from distal tubules by alkaline phosphatase histochemistry. Microdensitometry was performed using an IAS 2000 image analyzer (Delta Sistemi; Rome, Italy) connected via a TV camera to a Leica 500 AC microscope. Microdensitometry system was calibrated taking as zero the background obtained in sections exposed to non-immune serum. The intensity of immune staining was then measured using a microdensitometry program of the image analyzer. Microdensitometry technique was validated in a series of preliminary experiments in which the primary antibody was used in a range of dilutions from 1:10,000 to 1:500. The antibody dilution causing the lowest background (1:2500) was considered optimal and was used in subsequent microdensitometry experiments.
Antibodies and Chemicals
Rabbit anti-dopamine D4 receptor protein antibody (cat. no. 324405, lot no. B17850) was purchased from Calbiochem-Novabiochem (San Diego, CA). Peptide immunogen corresponding to that used for generating antibody was synthesized by the Department of Medicinal Chemistry of Camerino University. It corresponded to amino acids 175–186 for the human D4 receptor. The biotin-streptavidin immunostaining kit was from Calbiochem-Novabiochem. Other chemicals were purchased from Sigma Chemical (St Louis, MO).
Statistics
Data for microdensitometry of dopamine receptor protein immunostaining in the different types of tubule cells were analyzed statistically by ANOVA followed by Duncan's multiple-range test as a post-hoc test.
Results
The renal distribution of D4 receptors was investigated by immunohistochemistry using an anti-dopamine D4 receptor antibody raised against the amino acid fragment 176–185 of the human D4 receptor protein. The use of an antibody directed against the same peptide allowed the demonstration of D4 receptor expression in rat cortical collecting duct by immunohistochemistry (Sun et al. 1998). In Western blotting analysis, this antibody revealed a 40–42-kD band in rat brain frontal cortex and renal membrane preparations (Figure 1, Lanes 1 and 3). This apparent molecular weight corresponds to that expected for the dopamine D4 receptor protein (Van Tol et al. 1991) and previously reported for renal cortex (Sun et al. 1998). Immunodetection of the 40–42-kD band was completely prevented by antibody pre-incubation with the immunogen peptide (Figure 1, Lanes 2 and 4).
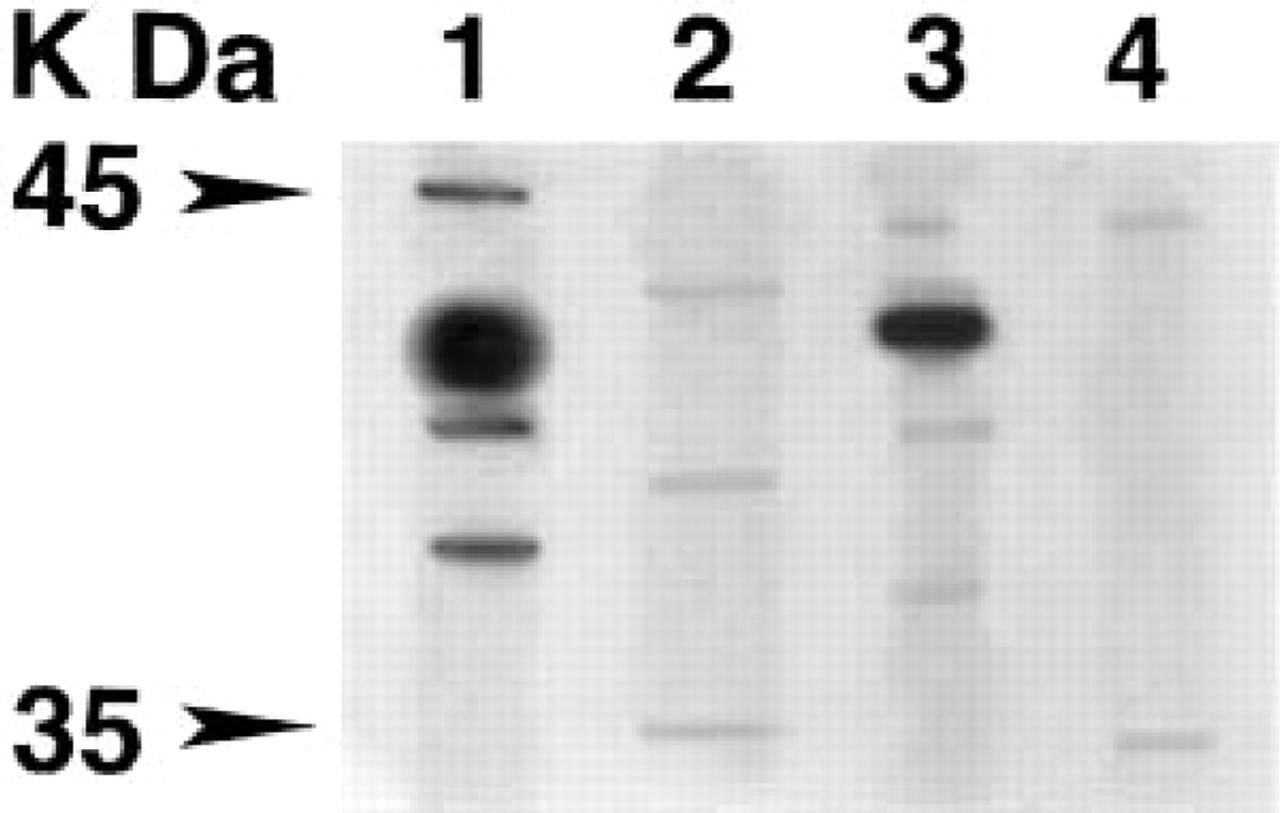
Western blotting analysis using the anti-dopamine D4 receptor protein antibody. Experiments were performed in membrane preparations from rat brain frontal cortex (Lanes 1 and 2) and kidney (Lanes 3 and 4). Lanes 2 and 4 show the results obtained with antibodies pre-adsorbed with the corresponding receptor blocking peptide. The blot is representative of three to five separate experiments.
Immunohistochemistry was performed in kidneys of sham-operated and denervated rats. The efficacy of renal denervation was evaluated by the measure of renal NE concentrations. In right kidneys of sham-operated rats, the NE concentration averaged 40 + 2.1 ng/g tissue. Denervation caused a decrease of NE concentration to 3.5 + 0.1 ng/g tissue (p<0.01 vs shamoperated). NE concentrations in left kidneys were not different from those assayed in right kidneys of shamoperated rats (data not shown). Sections of shamoperated kidneys exposed to antibodies against dopamine D4 receptor protein developed a sparse dark-brown staining in the adventitia-media of renal arteries (Figure 2A) and in an appreciable percentage (approximately 40%) of nerve fibers present in nerve trunks visible in the hilum (data not shown). Perivascular dopamine D4 receptor protein immunoreactivity was located in large segmental arteries and in arcuate, interlobar, and interlobular arteries (data not shown). This immunoreactivity disappeared in denervated kidneys (Figure 2C). Sections exposed to pre-immune serum or to antibodies pre-adsorbed with the corresponding receptor protein peptide developed background only (Figure 2E). Dopamine D4 receptor protein immunoreactivity was also located perivascularly in locations corresponding to afferent (Figure 2B) and, to a lesser extent, efferent (Figure 2B) glomerular arterioles. This immunoreactivity was no longer observed in denervated kidneys (Figure 2D) or after pre-exposure to antibodies pre-adsorbed with the corresponding receptor protein peptide (Figure 2F).
No dopamine D4 receptor protein immunoreactivity was observed in the glomerular tuft (Figures 2A and 2D) or in the loop of Henle epithelium (Table 1). Slight D4 receptor protein immunoreactivity was found in proximal and distal tubules (Table 1) and moderate immunoreactivity in collecting tubule epithelium (Figures 3A and 3C; Table 1). Reaction was specific for D4 receptor protein because it did not occur when an antibody pre-adsorbed with blocking peptide was used (Figure 3B) and was unaffected by renal denervation (Figure 3C; Table 1).
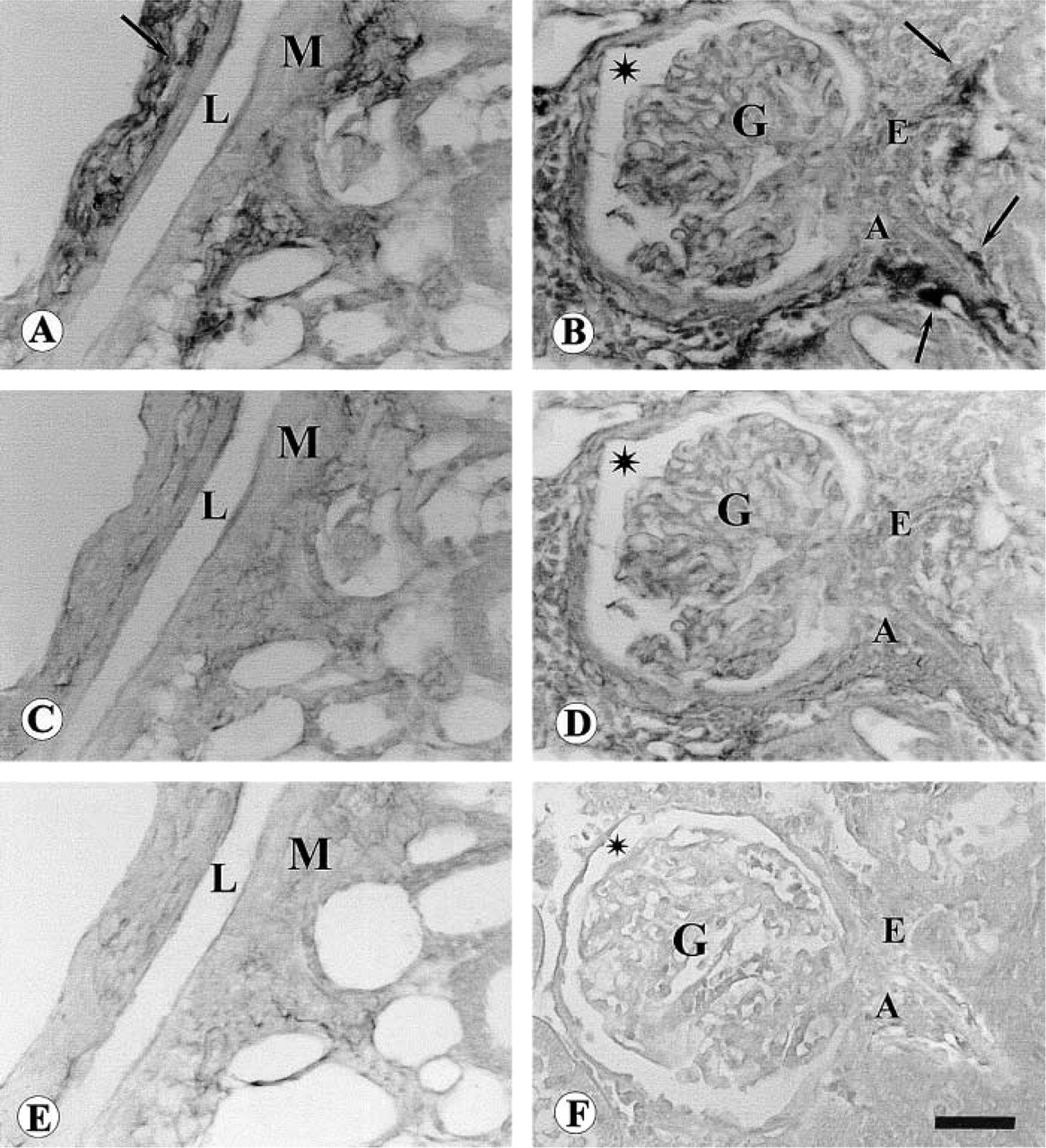
Micrographs of rat kidney processed for dopamine D4 receptor protein immunohistochemistry. (
Dopamine D4 receptor protein immunoreactivity in rat renal tubules a
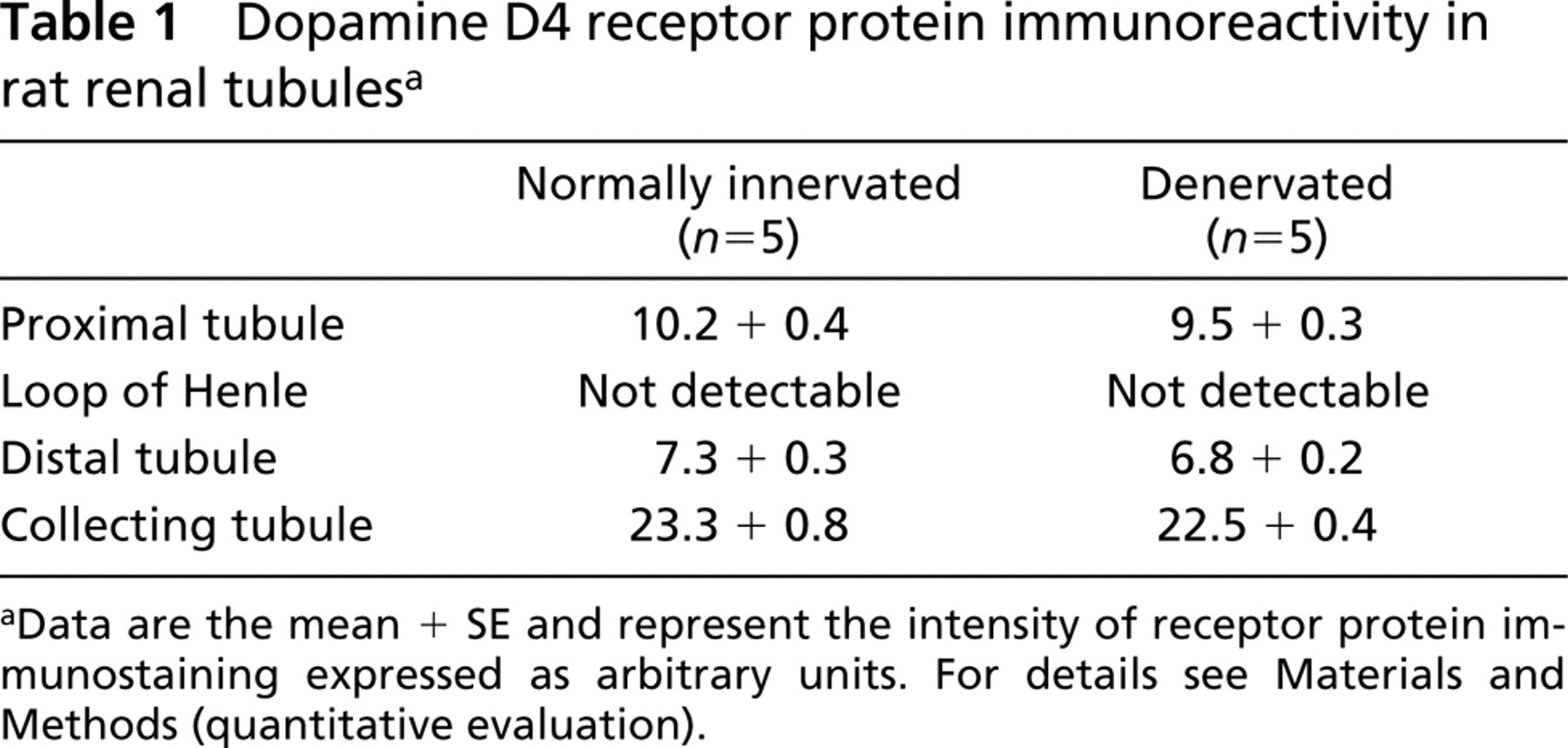
aData are the mean + SE and represent the intensity of receptor protein immunostaining expressed as arbitrary units. For details see Materials and Methods (quantitative evaluation).
Discussion
During the past decade, several efforts have been made to determine the molecular, pharmacological, and functional properties of dopamine D4 receptors. Most of the studies concerned the role of these receptors in the regulation of brain function and their abnormalities in neurological disorders (Oak et al. 2000). Peripheral D4 receptors have been relatively less well investigated. The relatively low density of these receptors and the lack of selective radioligands represented major problems for their identification and characterization. In this study, using a quantitative immunohistochemical technique, we defined their vascular and tubule distribution and, for the first time, we supplied evidence for the prejunctional localization of D4 receptors in rat kidney. According to data reported by other authors (Sun et al. 1998; Schafer et al. 2000), we showed that dopamine D4 receptor protein immunoreactivity was localized in renal collecting tubules. In addition, we observed a faint to moderate dopamine D4 receptor protein immunoreactivity in proximal and distal tubules.
In this study, extending the indirect results of other investigations (Amenta 1997; Sanada et al. 1997), we demonstrated the localization of dopamine D4 receptors in glomerular afferent and, to a lesser extent, efferent arterioles. The apparent discrepancy of these results with those of a previous study by our group in which no dopamine D4 receptor immunoreactivity was found in the rat renal vasculature (Amenta et al. 2000) may be due to strain differences (Wistar vs Sprague-Dawley rats) and/or to a sparser expression of prejunctional D4 receptors in the renal vasculature of Wistar rats. Because no other dopamine D2-like receptor subtypes have thus far been demonstrated in glomerular arterioles, our findings suggest that the D4 receptor is the receptor subtype playing a major role in the regulation of glomerular blood flow. The immunoreactivity of D4 receptors follows the same distribution of sympathetic nerve fibers (Lokhandwala 1988) and disappears after denervation of the kidney. These results suggest that D4 receptors are located prejunctionally, probably at the level of sympathetic neuroeffector junctions. Prejunctional dopamine D2-like inhibition of NE release was extensively characterized by functional studies and localized microanatomically by light microscopic autoradiography (Amenta 1990,1997; Lokhandwala and Amenta 1991; Cheung and Barrington 1996; Jose et al. 1998). These receptors mediate vasodilatation indirectly by decreasing the noradrenergic vasoconstrictor tone (Lokhandwala 1988; Vega et al. 1990; Rump and Schollmeyer 1993) and are markedly reduced in a model of denervation similar to that used in the present study (Barili et al. 1997b). In the central nervous system, both dopamine D2 and D3 receptors can represent autoreceptors located presynaptically (Tepper et al. 1997). Recently, Svingos and co-workers reported that, in the rat nucleus accumbens shell, most of the D4 receptors are located in the presynaptic endings (Sanada et al. 1997). Our findings suggests that, as observed in the CNS, renal D4 receptors have a prejunctional localization and may behave as autoreceptors. It is noteworthy that the prejunctional localization of renal D4 receptors appears to be limited to blood vessels because renal denervation did not modify the immunoreactivity observed in tubuli. This suggests that prejunctional D4 receptors may play a role in the indirect control of vascular tone rather than in the regulation of NE release and dopamine effects in tubuli.
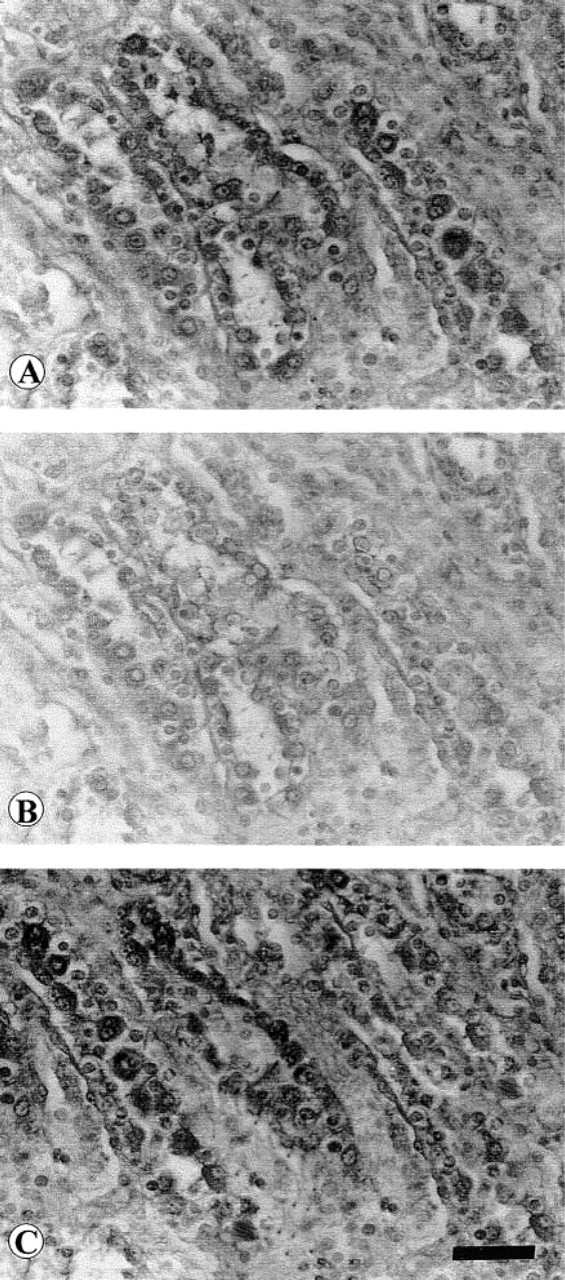
Micrographs of renal collecting tubules processed for dopamine D4 receptor protein immunohistochemistry. (
In conclusion, we have demonstrated that dopamine D4 receptors are distributed in different segments of the nephron and, in the vascular distribution, they also have a prejunctional localization. These results may represent a starting point for the further characterization of the functional properties of D4 receptors in kidney and for comprehension of the mechanisms of renal function regulation by dopamine.
Footnotes
Acknowledgements
Supported in part by an INSERM/CNR collaborative grant and by a financial grant from Servier Laboratories.