
Abstract
The major structural protein in skeletal muscle, myosin heavy chain (MyHC), is primarily transcriptionally controlled. We compared the expression of MyHC isoforms on the mRNA and protein level in biopsies from the m. gluteus medius from adult untrained horses. In transverse sections, the majority of fibers showed qualitatively identical mRNA and protein expression patterns. However, coexpression of 2a and 2d/x MyHCs was substantially more common at the protein than at the mRNA level, suggesting a fine-tuning of these two genes in normal muscle not subjected to any training protocol. Because transverse sections give a limited sampling of mRNA expression in the case of uneven distribution of transcripts in a muscle fiber, we also analyzed longitudinal sections. We present, for the first time, evidence that expression of MyHC mRNA and protein was equal along the length of the fiber. Hence, mRNA expression is not regulated by differential expression of isoforms by separate myonuclei. It is concluded that the number of protein hybrid fibers in equine gluteus medius muscle is controlled by alteration of the transcription pattern uniformly along the fiber, rather than by simultaneous transcription of genes. The differences with the results in muscle of small animals and humans are discussed.
S
MyHC proteins are primarily transcriptionally controlled; the time course and threshold stimuli needed to trigger changes at the mRNA level are thus important aspects of gene regulation (O'Neill et al. 1999). In fibers that undergo fiber-type transformation, changes in MyHC mRNA should precede changes in the corresponding protein products, leading to an at-least-temporary mismatch in the distribution of transcripts and corresponding protein products (Andersen and Schiaffino 1997; Jaschinski et al. 1998). By comparing changes in expression of both MyHC mRNA and protein, a clue can be obtained for the direction of change in MyHC gene expression. We previously studied mRNA and protein expression in serial transverse sections of the gluteus medius muscle of the horse. We showed in one horse that the majority of fibers analyzed had the same RNA and protein expression, whereas a minority of fibers showed a mismatch. Most of these mismatches concerned the common type of hybrid fibers, expressing both MyHC type 2a and 2d protein but only a single MyHC RNA (Eizema et al. 2003). However, the mismatch between mRNA and protein MyHC expression observed in transverse sections could be the result of non-homogeneous distribution of the mRNA along the length of the fiber (Newlands et al. 1998). In the present paper we extend and confirm our previous observations by including data from more horses and demonstrate that MyHC expression in hybrid fibers is regulated at the transcriptional level. To test the homogeneity of mRNA and protein expression along the length of a fiber, we analyzed, for the first time, longitudinal sections by in situ hybridization and immunohistochemistry.
Materials and Methods
All chemicals were obtained from Merck (Amsterdam, The Netherlands) unless otherwise indicated.
Animals
Samples were taken from four adult, untrained healthy horses, two Dutch Warmblood (DW) mares (4 and 8 years old, horse B and horse C), one DW gelding (15 years old, horse D), and one Frisian stallion (26 years old, horse A). All procedures were reviewed and approved by the Animal Care Committee of our Institution.
Muscle Biopsies
Percutaneous muscle biopsies from the gluteus medius were taken according to the protocols of Lindholm and Piehl (1974) and Dingboom et al. (1999). Biopsies were taken on an imaginary line drawn from the coxal tuber to the sacral tuber, at one-third distance from the sacral tuber, perpendicular to the skin. They were taken as deep as possible (until resistance from the iliac wing was reached). Tissue samples were either open (horse A and horse D, 500 mg) or needle (horse B and horse C, 100 mg) biopsies, taken from the same location in the muscle to ensure comparable fiber-type composition (Serrano et al. 1996; Linnane et al. 1999). Samples were divided into small portions (maximum 50 mg) and rolled in talcum powder, mounted on cork blocks with the use of OCT embedding medium, and oriented so that the fibers could be sectioned either transversely or longitudinally and frozen in liquid nitrogen. All samples were stored at –80C. Longitudinal sections were analyzed from horse A and horse D.
In Situ Hybridization
Ten-μm transverse or longitudinal serial sections were made with a cryostat at –20C, placed on Superfrost Plus slides (Menzel; Merck, Amsterdam, The Netherlands), dried for 1 hr, fixed for 20 min with 4% paraformaldehyde in 1× phosphate buffered saline (PBS, pH 7.4), washed in 1× PBS, dehydrated, and stored at –80C. In situ hybridization was performed as described previously (Eizema et al. 2003). In brief, slides were returned to room temperature, rehydrated, rinsed, and digested with 20 μg/ml of proteinase K at 37C for 3 min. Acetylation was performed with 0.25% acetic anhydride in 0.1 M triethanolamine for 10 min. After several rinses, refixation, and several washes, sections were completely dehydrated before hybridization. cDNAs included MyHC 1 (276 bp) in pGEM-T (Promega; Leiden, NL), 2a (278 bp) in pGEM-T, and 2d/x (282 bp) in pCRII-TOPO (Invitrogen; Breda, NL) (Eizema et al. 2003). Riboprobes were synthesized according to the manufacturer's guidelines (Roche Molecular Biochemicals; Almere, The Netherlands) and purified by a Qiagen RNeasy kit (Westburg, Leusden; The Netherlands). The riboprobes (500 ng/ml final concentration) were suspended in 40% (deionized) formamide, 1× SSC (0.15 M NaCl, 0.015 M NaCitrate), 10% dextran sulfate, 1× Denhardt solution (0.02% Ficoll, 0.02% polyvinilpyrrolidine, 0.02% BSA), 0.67 M NaCl, 0.1 μg/μl yeast tRNA, and 0.1 μg/μl herring sperm DNA and heated at 80C for 5 min. Prehybridization was performed for 30 min. Approximately 30 μl of probe was used per slide, overlaid with a coverslip. Hybridization was performed overnight at 45C in a humidified In Slide Out incubator (Boekel Scientific, Merck; Amsterdam, The Netherlands). Coverslips were removed, followed by two high-stringency washing steps at 60C for 20 min in 0.5 × SSC and 20% formamide and two rinses in 2× SSC at room temperature. Unhybridized probe was digested with 1 μg/ml RNase A in 0.5 M NaCl, 10 mM Tris-HCl, pH 8.0, at 37C for 30 min, followed by five washes in 2× SSC at room temperature, and another high-stringency wash for 10 min. The sections were rinsed twice with 2× SSC and maleic buffer (0.1 M maleic acid 0.15 M NaCl, pH 7.5). Tissue sections were blocked with 5% inactivated BSA in maleic buffer at room temperature. Blocking buffer was replaced with sheep anti-digoxigenin Fab-alkaline phosphatase conjugate (1:2000 in 1% BSA in maleic buffer) overnight at room temperature. Several washes with the same buffer were followed by washes with 0.1 M NaCl, 0.1 M Tris-HCl, pH 9.5, 50 mM MgCl2, and 0.1% Tween 20 at room temperature. Alkaline phosphatase activity was visualized by incubation with 0.18 mg/ml BCIP (5-bromo-4-chloro-3-indoyl-phos-phate), 0.34 mg/ml nitroblue tetrazolium, in the buffer described above. The staining was allowed to develop for ≃16 hr at room temperature, rinsed with distilled water, and embedded in Aquamount (BDH; Boom, Meppel, The Netherlands).
Immunohistochemical Staining
Monoclonal antibodies (MAbs) used were previously shown to cross-react with horse myosins (Dingboom et al. 2002). MAb 219 reacts with type 1, MAb 333 reacts with type 2a, and MAb 412 reacts with type 1 and 2d/x. MAb A4.74 was obtained from Alexis (Kordia; Leiden, The Netherlands) and recognizes MyHC type 2a. Transverse serial and longitudinal sections (10 μm) were obtained as described above. The protocol was followed as described previously (Dingboom et al. 2002; Eizema et al. 2003). Briefly, the slides were rinsed in PBS, blocked in Teng-T (10 mM Tris, 5 mM EDTA, 0.15 M NaCl, 0.25% gelatin, and 0.05% Tween 20, pH 8.0) for at least 15 min, followed by rinsing in PBS, and incubation overnight at room temperature with the MAbs at a dilution of 1:10 (333), 1:25 (219, 412), or 1:50 (A4.74) in PBS. The slides were rinsed with PBS and incubated for 90 min with a biotinylated horse anti-mouse polyclonal antibody (1:100 in PBS, ABC-peroxidase staining kit; Elite, Vector Laboratories, Burlingame, CA), rinsed in PBS, and incubated for 90 min with the components avidin (1:100 in PBS) and biotin (1:100 in PBS) of the ABC staining kit. After rinsing, visualization was by incubation with 0.05% 3, 3'-diaminobenzidine tetrachloride in 30 mM imidazole and 0.09% H2O2. The slides were subsequently embedded in DePeX (BDH). For MAb 412, the slides were rinsed in 0.1 M acetate buffer (pH 6.0), and di-ammonium nickel sulfate (2.5% in acetate buffer) instead of imidazole was used.
Analyses
Between 146 and 296 fibers per biopsy were used for fiber typing and calculation of fiber-type composition. The fibers were classified into type 1, type 2a, type 2ad, and type 2d/x on the basis of their reactions with the different in situ probes and the MAbs.
Results
Identification of Myofiber Types by In Situ Hybridization and Immunohistochemistry
We analyzed the myofiber types of the gluteus medius muscle of four different horses by using in situ hybridization and immunohistochemistry on transverse serial sections. Figure 1 (left panels) shows serial sections from the equine gluteus medius muscle processed for in situ hybridization with mRNA probes specific for MyHC 1, 2a, and 2d isoforms. The fiber type is indicated according to the mRNA expression analyses. These classifications were compared with the corresponding fiber types according to the protein expression analyses (Figure 1, right panels). Clearly, the staining pattern of the in situ hybridization and immunohistochemistry experiments show a high correlation. Very few of the total analyzed fibers coexpressed two MyHC transcripts (Table 1). The majority expressed one mRNA isoform. The only coexpression detected was in a small minority of fibers expressing 2a and 2d/x MyHC transcripts (type 2ad) (0–1.7%, Table 1). On the protein level, however, many more hybrid fibers (15.4–34.4%) were detected, and they were all the type 2ad fiber (Table 1).
Of all fibers analyzed, the majority had identical mRNA and protein expression, but roughly 20% showed a mismatch (Table 1). Most of these mismatches concerned fibers expressing type 2ad protein. In fact, except for one fiber (in horse B), type 2ad protein fibers never showed a match with 2ad mRNA. Two examples are seen in Figure 1, expressing type 2a mRNA but type 2ad protein (asterisks). Of the type 2ad fibers at the protein level, the majority expressed only type 2a mRNA, a minority only type 2d/x mRNA, and 0.3% (one fiber in one animal) expressed type 1 mRNA (Table 1). In total this results in a higher number of pure 2a fibers on the mRNA level as compared with the protein level, the number of pure 2d fibers being approximately equal. An exception was the result obtained with the 15-year-old gelding (results in Table 1D), where half of the type 2ad protein fibers expressed type 2a mRNA and the other half expressed type 2d mRNA.
In this paper we compare results obtained by comparing two different techniques introduced in our previous paper (Eizema et al. 2003). Analyses of the results must take into account that differences can arise from differences in sensitivity of the two techniques. We have extensively optimized the in situ hybridization procedure (e.g., by following staining patterns over a prolonged period of time); the detection limit is quite low. The immunohistochemistry protocol is also optimized to give the best signal-to-noise ratio. With these optimized protocols we have performed the described experiments and obtained reliable results.
Analysis of Longitudinal Sections
A hypothesis for the occasional appearance of a protein without its mRNA is that the entire fiber in question contains mRNA for more than one MyHC isoform, but that each nucleus expresses only a single mRNA or expresses it in pulses as was demonstrated in murine muscle by Newlands et al. (1998). In this case, transverse sections would give a limited sampling of mRNA expression in a muscle fiber. To test this hypothesis we analyzed longitudinal sections of horse A and horse D by in situ hybridization and immunohistochemistry. Figure 2 depicts a representative specimen of horse D. The left panels show the mRNA expression, the right panels the protein expression. The mRNA expression is slightly higher under the sarcolemma indicating the localization in the sarcoplasm, in agreement with the study of Russell et al. (1992). No differences in mRNA or protein expression level were seen along considerable stretches of fiber (≃500 μm) containing many myonuclei (distance between myonuclei ≃20 μm) (Murphy 1998) (Figure 2). We have also analyzed longitudinal sections of two other unrelated KWPN horses by in situ hybridization with the same results (results not shown). Unfortunately, comparison of different protein and mRNA expression within the same fiber was not possible because the fibers could not be aligned due to limitations in fiber diameter. We tried to use 5-μm instead of 10-μm sections to improve the alignment; however, these thin sections proved not suitable for in situ hybridization experiments. The distribution of the transcripts along the length of the fiber (Figure 2) was homogeneous. Switches of fiber type along the length of the fiber or localized areas with deviating MyHC expression as observed in human vastus lateralis fibers of very old subjects was never seen (Andersen 2003).
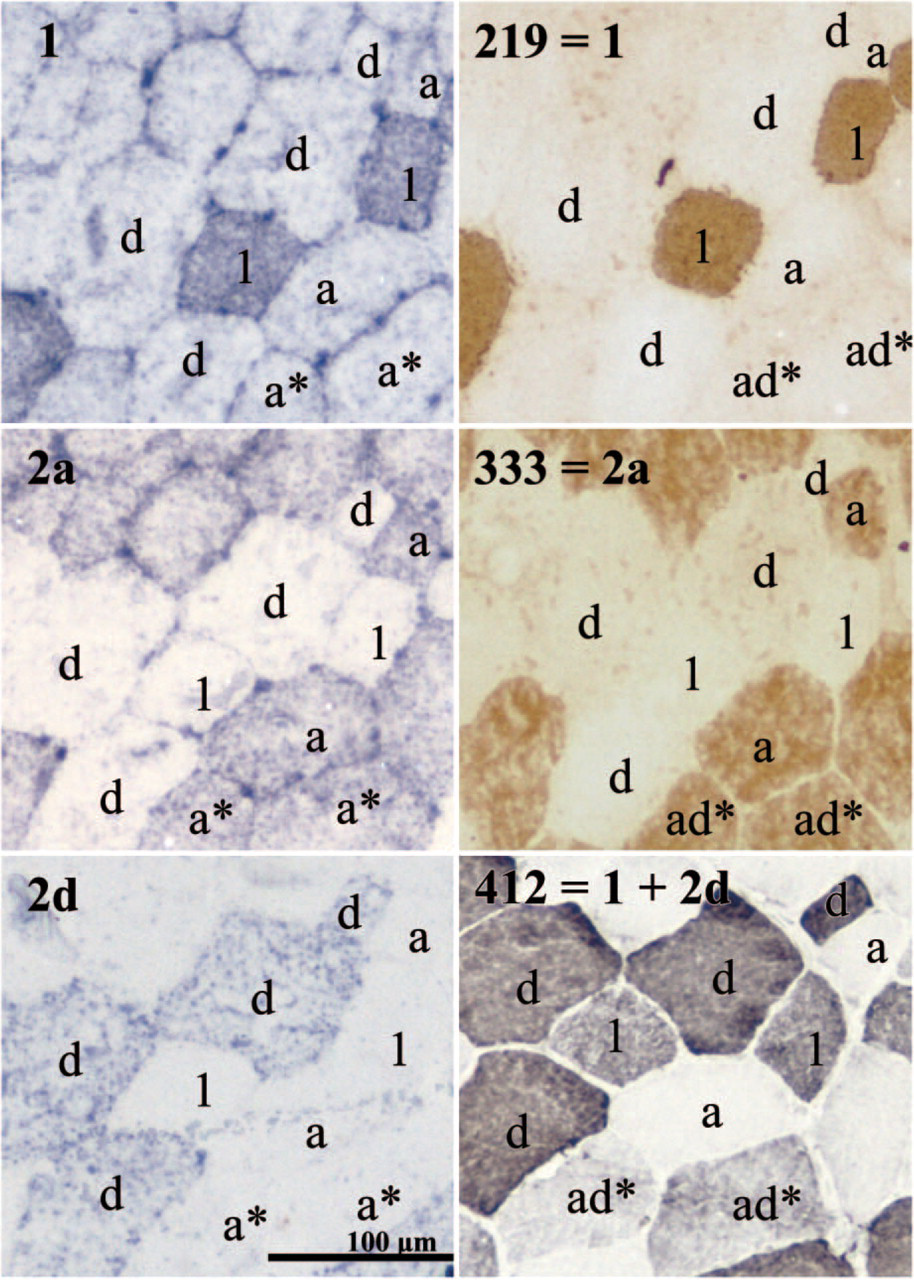
Identification of myofiber types by in situ hybridization using non-radioactive myosin heavy chain (MyHC) mRNA probes (left panels) and immunohistochemistry (right panels) in serial sections (from the same biopsy as Table 1C). The specificity of the probes (left panels) and antibodies (right panels) are indicated. Fiber types are indicated according to the mRNA expression (left panels) or according to the protein expression (right panels).
Calculation of myosin heavy chain (MyHC) fiber types (%) present in the equine gluteus medius muscle as assessed by in situ hybridization (RNA) and immunohistochemistry (protein) on transverse serial sectionsa
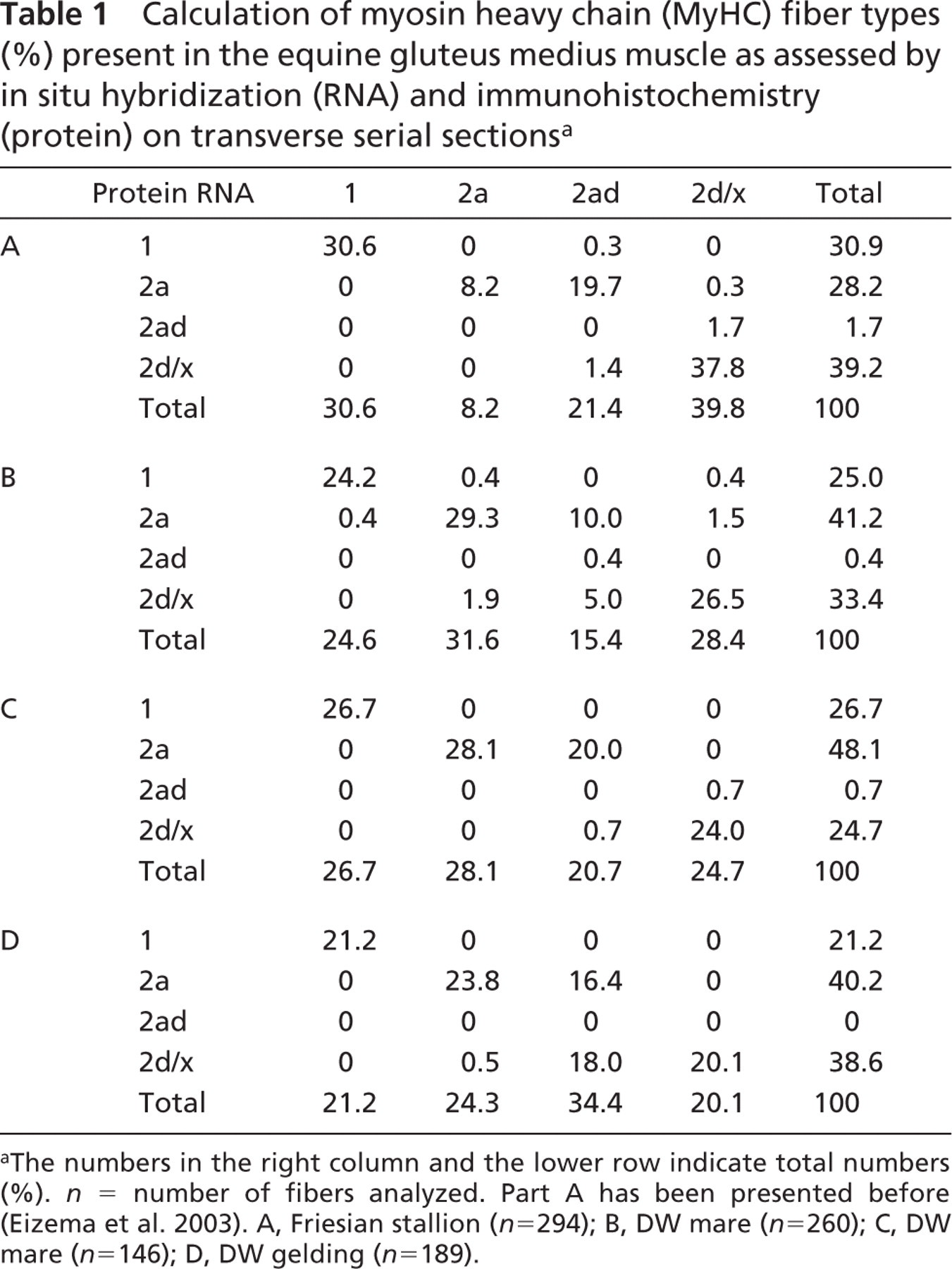
The numbers in the right column and the lower row indicate total numbers (%). n = number of fibers analyzed. Part A has been presented before (Eizema et al. 2003). A, Friesian stallion (n = 294); B, DW mare (n = 260); C, DW mare (n = 146); D, DW gelding (n = 189).
Additionally, we analyzed 12 consecutive transverse 10-μm sections (total of 120 μm) stained with the different mRNA probes. Again, fibers were positive in all 12 sections for the MyHC mRNA tested (tested for all three probes, results not shown), indicating consistent mRNA expression.
Discussion
In the present paper we demonstrate conclusively that MyHC expression in hybrid fibers is regulated at the transcriptional level. The MyHC mRNA and protein is homogeneously expressed along the length of a fiber as analyzed for the first time using longitudinal sections assayed by in situ hybridization and immunohistochemistry. The distribution of transcripts along the length of the fiber was homogeneous in contrast with the results of Newlands et al. (1998). Using regenerating or isolated single fibers, they showed that not all muscle nuclei were transcriptionally active at a given time. Comparison of the results of this paper with the results of Newlands et al. (1998) is difficult because different organisms, muscle, experimental conditions, and proteins were studied. An intriguing hypothesis, however, could be that the MyHC genes are transcribed in all nuclei as opposed to transcription in pulses for the thin-filament genes studied in Newlands et al. (1998). Clearly, expression studies of the same proteins and using the same experimental conditions could provide more insight into this issue.
Most fibers analyzed expressed a single, corresponding, MyHC isoform on the mRNA and protein level, but mismatches were the rule in hybrid fibers. In these, two MyHC protein isoforms were expressed, but the mRNA for only one of them (interestingly mostly 2a) was present. The low level of coexpression at the mRNA level and the homogeneous expression along a considerable length of fiber suggests that a hybrid fiber synthesizes only one mRNA isoform at a time. Fibers containing two MyHC proteins are most probably converting to the type corresponding to the expressed mRNA; the other isoform is no longer synthesized and will progressively disappear as a result of protein turnover. Hence, regulation of hybrid fibers is transcriptionally controlled in the equine gluteus medius muscle. Although hybrid MyHC 2ad protein fibers are regarded as a ‘true’ type because of their regular appearance, they can also be classified as transitory fibers as suggested by Pette and Staron (1997) and demonstrated in this paper (Andersen et al. 1999b; Linnane et al. 1999; Rivero et al. 1999; Stephenson 2001; Dingboom et al. 2002). The appearance of hybrid and mismatched fibers indicates that fiber-type transition occurs in presumably normal skeletal muscles not subjected to any training protocol.
Here we present the absence of hybrid 2ad fibers on the mRNA level in equine gluteus medius muscle. The same results were presented for longissimus and rhomboideus muscle of the pig, also a large animal (Lefaucheur et al. 2002). In human vastus lateralis the vast majority of MyHC mRNA profiles, including hybrid fibers, corresponded with the protein isoforms detected (Ennion et al. 1995; Andersen and Schiaffino 1997; Andersen et al. 1999a; Serrano et al. 2001). In a comparative study of 14 different human muscles, ≃60% of the hybrid 2ad fibers on the protein level were also 2ad hybrids on the mRNA level, ≃30% expressed only 2a mRNA, and 10% expressed only 2d mRNA (Smerdu et al. 1994; Smerdu and Erzen 2001). Unfortunately, the authors did not state from which muscles these particular hybrid fibers were isolated.
In rabbit gastrocnemius, psoas, and adductor magnus muscle, single-fiber analysis revealed that, depending on the muscle, the number of hybrids was equal or larger at the mRNA level than at the protein level (Peuker and Pette 1997). Another study (Conjard et al. 1998) analyzed single fibers from rabbit rectus femoris, extensor digitorum longus, tibialis anterior, and soleus muscle. mRNA MyHC isoform patterns were consistent with MHC protein profiles, including hybrid fibers, in the majority of fibers from normal, untransforming muscle (Conjard et al. 1998). In rat, single fibers from the soleus muscle were analyzed; in this muscle more hybrid fibers were again detected on the mRNA level than on the protein level (Stevens et al. 1999).
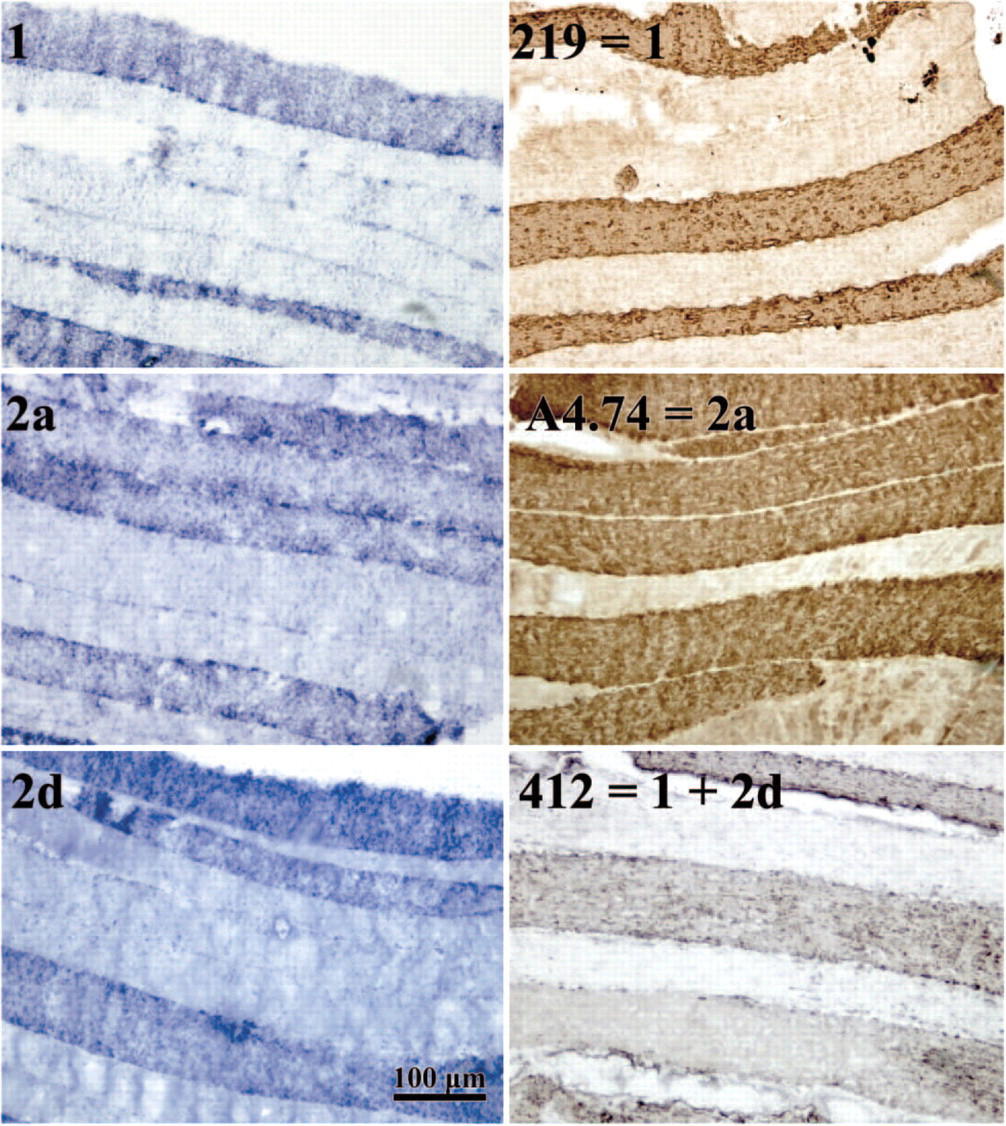
Part of a longitudinal section (from the same biopsy as Table 1D) stained with the indicated mRNA probes (left panels) or with the indicated antibodies (right panels). Alignment of the sections is not possible. Clearly, the staining is equal along the visible fiber length.
From the above-mentioned studies it is clear that the detection of hybrids in normal, non-trained muscle is a general phenomenon. The number of hybrid fibers is, however, highly species and muscle specific. Studies using small animals show a higher number of hybrids on the mRNA level as compared with the protein level. Human muscles show equal or lower numbers, whereas the horse (this study) and pig muscles show (almost) no hybrid fibers on the mRNA level. These differences might be due to the size of the animal, which imposes different demands on the muscle. However, the architecture and innervation of the muscle can also be important factors. Small animals have a different architecture and pattern of innervation as compared with primates (Paul 2001; Paul and Rosenthal 2002). In the study of Paul (2001), it was shown that non-primates have fascicles shorter than 35 mm with generally single fibers from end to end, whereas primate fascicles could be as long as 140 mm. Data on fascicle length and innervation of horse muscle are limited. A study of elbow extensor muscles showed fiber length of 20–30 mm (Ryan et al. 1992). Other studies detected very variable fiber lengths ranging from 17 mm to 90 mm (Brown et al. 2003; Zarucco et al. 2004). From these limited data it can be concluded that series-fibered muscle seems to be more common in the horse than long single fibers stretching from tendon to tendon. Regulation of fiber type of a single fiber stretching the full fiber length is probably different as compared with series-fibered muscle, a possible explanation for the difference in results.
Because humans and horses do not express 2b MyHC protein in their limb muscles, it is conceivable that the existence of hybrid 2ad fibers is even more important for the fine-tuning of contraction properties as compared with animals that have three fast isoforms available (Horton et al. 2001; Eizema et al. 2003; Chikuni et al. 2004).
Several protocols applied to induce transformation of muscle fibers show that upon transformation the amount of hybrid fibers increases, even up to the expression of three MyHC protein isoforms (Talmadge 2000). Upon this induced transformation, regardless of the type of protocol applied, more mismatching between mRNA and protein expression is observed (Andersen and Schiaffino 1997; Conjard et al. 1998; Andersen et al. 1999a; Stevens et al. 1999). Clearly, analyzing muscle fibers (pure and hybrid) at the mRNA and protein level is important for understanding the steady-state situation and the transformation process. The regulation of muscle MyHC fiber type is a complex process, fine-tuned by transcriptional regulation of the expression of different MyHC mRNAs, enabling the muscles to tune even with minimal changes in mechanical demands.
Footnotes
Acknowledgements
We thank Dr. A.F.M. Moorman and Dr. J.A.M. Korfage (University of Amsterdam) for the generous gift of the monoclonal antibodies. We are grateful for the skillful technical assistance of C.W. van der Wiel.