
Abstract
It has been known that group II phospholipase A2 (PLA2) mRNA and protein are present in the homogenates of the spleen, lung, liver, and kidney in normal rats, but the cellular origin of this enzyme has not been yet identified. At present, five subtypes of group II PLA2 have been identified in mammals. Antibodies or mRNA probes previously used for detecting group II PLA2 need to be evaluated to identify the subtypes of group II PLA2. In this study we tried to identify group IIA PLA2-producing cells in normal rat tissues by in situ hybridization (ISH) using an almost full-length RNA probe for rat group IIA enzyme. Group IIA PLA2 mRNA was detected in megakaryocytes in the spleen and Paneth cells in the intestine by ISH. These cells were also immunopositive for an antibody raised against group IIA PLA2 isolated from rat platelets. Group IIA PLA2 mRNA-positive cells were not detected in lung, liver, kidney, and pancreas. Under normal conditions, group IIA PLA2-producing cells are splenic megakaryocytes and intestinal Paneth cells in rats.
Phospholipase A2 (PLA2) comprises a diverse family of enzymes that catalyzes the hydrolysis of glycerophospholipids at the sn-2 position to produce free fatty acids and lysophospholipids (Tischfield 1997; Valentin et al. 1999). At present, mammalian secretory PLA2s are classified into groups I, II, V, and X. Group II PLA2 has been further classified into five subtypes (Type IIA, IIC, IID, IIE, and IIF) on the basis of their primary structures. Group II PLA2 is one of the key enzymes in the process of inflammation that regulates the synthesis of arachidonic acid and lysophospholipids (Nevalainen 1993; Nevalainen et al. 1993). High plasma levels of group II PLA2 have been observed in patients with septic shock, multiple injuries, rheumatoid arthritis, and acute pancreatitis, and a specific inhibitor of group II PLA2 improved the mortality in experimental pancreatitis (Yoshikawa et al. 1999).
Group II PLA2 mRNA and protein were detected in homogenates of the spleen, lung, liver, and kidney of normal rats (Murakami et al. 1988; Ishizaki et al. 1989; Inada et al. 1991a; Hara et al. 1995; De Windt et al. 1997), but the cellular origins of this enzyme have not been yet identified. Although PLA2, referred to as group II PLA2 in previous reports, was assumed to be a group IIA subtype, in the light of recent findings on the diversity of this group of enzymes, characteristics of antibodies or mRNA probes need to be specified. In the present study, therefore, we prepared an almost full-length RNA probe for group IIA PLA2 mRNA and tried to identify group IIA PLA2-producing cells in normal rat tissues using both ISH and IHC techniques.
Materials and Methods
Preparation of Rats
Male Wistar rats (180–220 g) were anesthetized with an IP injection of pentobarbital sodium (50 mg/kg) and transcardially perfused with Tyrode's solution and then with 4% paraformaldehyde in 0.01 M PBS, pH 7.4. The jejunum, spleen, lung, liver, kidney, and pancreas were removed and postfixed in the same fixative for 3 hr. They were then immersed in a series of cold sucrose solutions (10%, 20%, 30% sucrose in PBS). Ten-μm-thick sections were cut on a cryostat microtome (Bright; Huntingdon, UK) and mounted on silanized slides (DAKO; Kyoto, Japan). The sections were dried on a hotplate at 37C and then kept frozen until use.
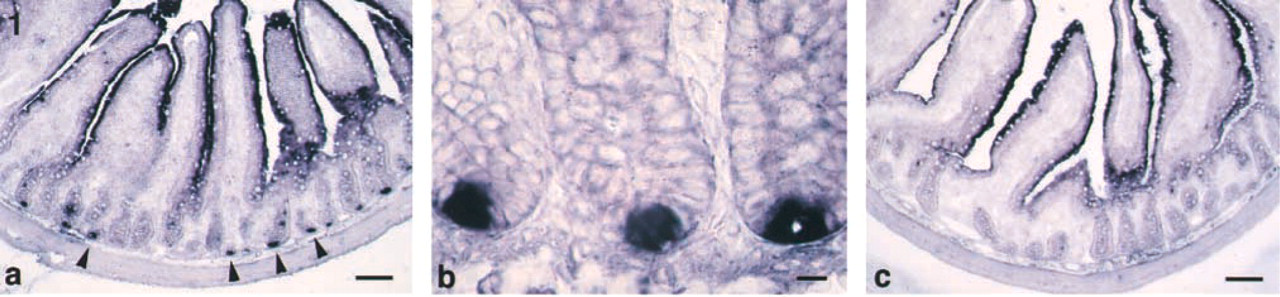
Group IIA PLA2 mRNA distribution in the rat jejunum. ISH with antisense probe for group IIA PLA2 (
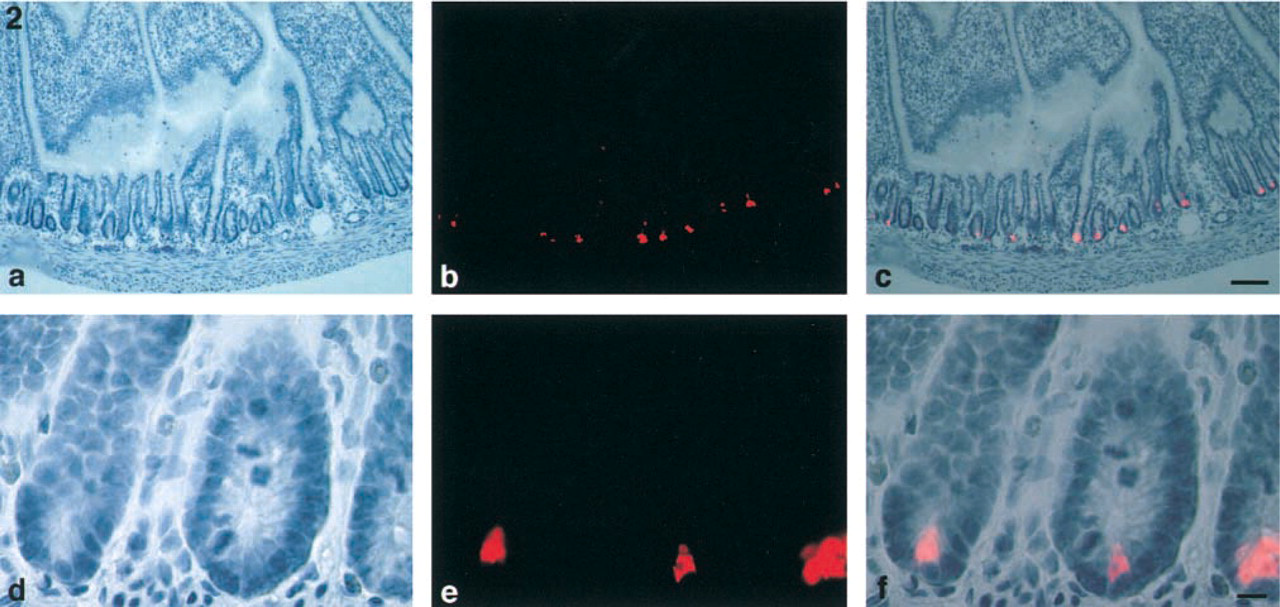
Group IIA PLA2-positive cells in the rat jejunum. Hematoxylin staining (
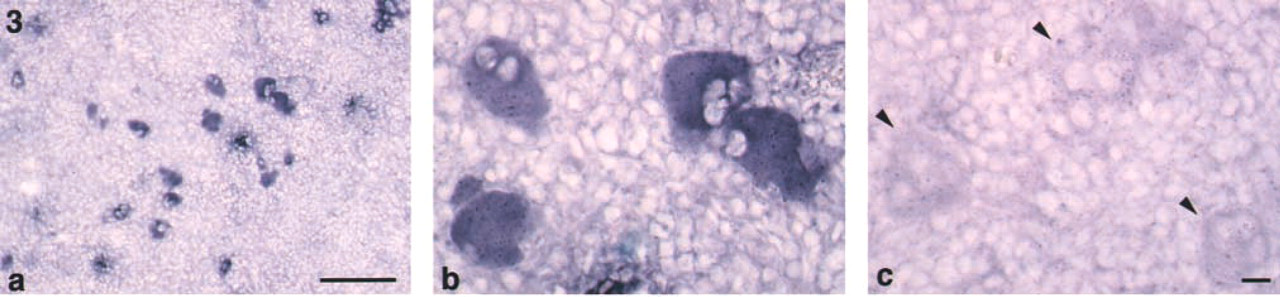
Group IIA PLA2 mRNA expression in the rat spleen. ISH with antisense probe for group IIA PLA2 (
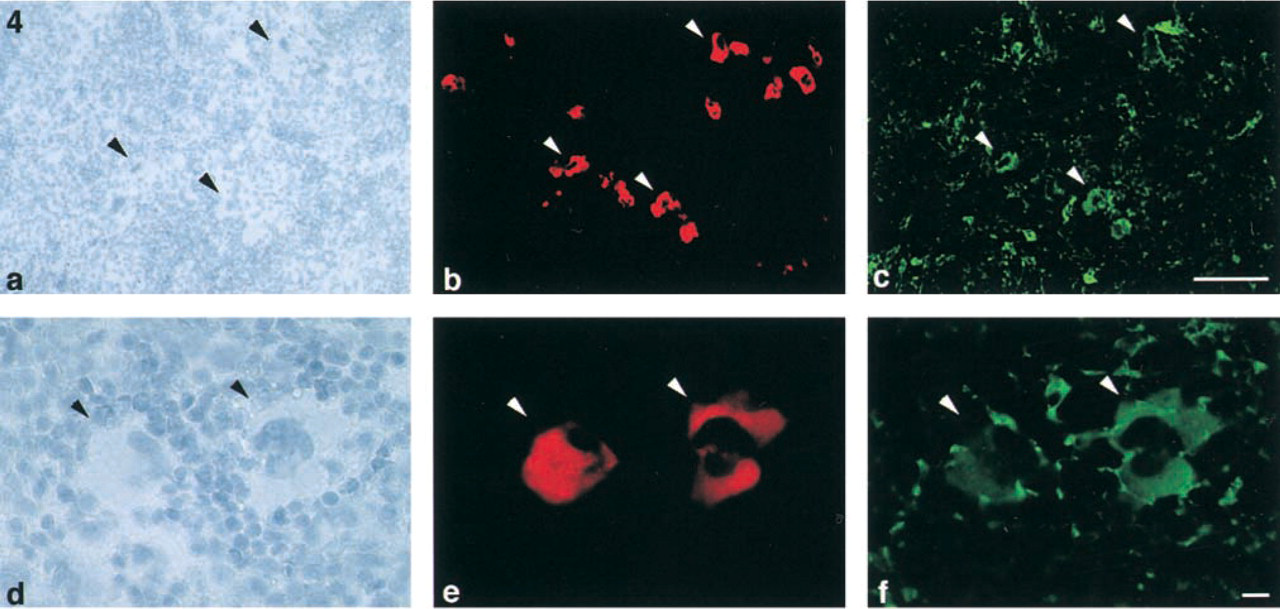
Group IIA PLA2-positive megakaryocytes in rat spleen. Hematoxylin staining (
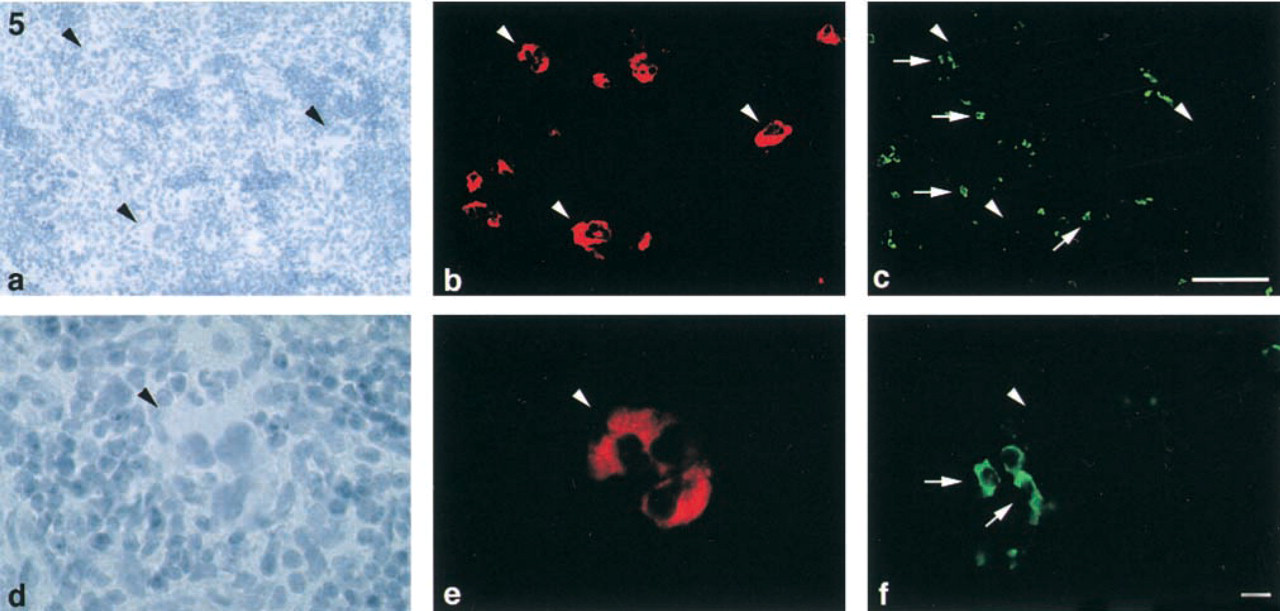
Group IIA PLA2 and macrophages in rat spleen. Hematoxylin staining (
Digoxigenin-labeled RNA Probe
We used a 0.7 kilobase pair fragment of digoxigenin-labeled RNA probe in the present study. This RNA probe corresponds to bp 76–748 of group IIA PLA2 mRNA, which covers about 90% of the whole mRNA. The NheI/EcoRI 0.7-kb pair fragment of rat group II PLA2 cDNA was excised from its full-length cDNA (Ishizaki et al. 1989) and subcloned into the XbaI/EcoRI sites of the pGEM-3Z. Antisense digoxigenin-labeled RNA probes were prepared using digoxigenin-11-UTP and T7 RNA polymerase to transcribe the group II PLA2 insert cDNA. Sense strand RNA probes were generated using SP6 RNA polymerase.
In Situ Hybridization
The sections were incubated in 0.3% Triton X-100 in PBS for 5 min and immersed in 10 μg/ml proteinase K (Boehringer Mannheim Biochemical; Indianapolis, IN) in PBS for partial proteolysis for 10 min at 37C. They were postfixed in 4% paraformaldehyde in PBS for 10 min and immersed in a solution containing 0.25% acetic acid anhydride (Nippon Gene; Toyama, Japan), 0.1 M triethanolamine for 20 min at room temperature (RT). They were then prehybridized in 50% formamide in 2 × SSC for 30 min at 42C. Hybridization was done with digoxigenin-labeled RNA antisense probe diluted to 1 μg/ml in hybridization buffer in a moist chamber for 16 hr at 42C. For negative control, digoxigenin-labeled RNA sense probe was hybridized to a section adjacent to the test section. After hybridization, the slides were washed in 50% formamide in 2 × SSC for 60 min at 42C and treated with 20 μg/ml RNase A in 0.5 M NaCl, 10 mM Tris-HCl, 1 mM EDTA (pH 8.0) for 30 min at 37C. They were rinsed in the same buffer at 37C for 30 min, then in 0.1 × SSC for 60 min at 42C, and finally in PBS for 1 min at RT. For immunological detection of digoxigenin, the slides were preblocked in 1% skim milk fraction in PBS for 30 min at RT and then incubated with alkaline phosphatase-conjugated polyclonal anti-digoxigenin Fab fragments (Boehringer Mannheim) (1:500 dilution) for 90 min at RT. After incubation, the slides were washed in PBS and developed in a solution of nitroblue tetrazolium and X-phosphate for 16 hr at RT. The color reaction was stopped using 1 mM EDTA, 10 mM Tris-HCl. The sections were mounted in Vectashield (Vector Laboratories; Burlingame, CA).
Immunohistochemistry
Rabbit anti-rat group II PLA2 IgG raised against purified PLA2 released from thrombin-stimulated rat platelets was used for histochemistry. The slides were rinsed with PBS and nonspecific binding was blocked in 1% skim milk in PBS. Sections were incubated with rabbit anti-rat group II PLA2 antibody. (Nakano et al. 1990) (1:2000 dilution) in a moist chamber overnight at RT, followed by Alexa546-conjugated goat anti-rabbit IgG (Molecular Probes; Eugene, OR) (1:100 dilution). Double-labeling studies were performed to characterize the group II PLA2-positive cells. For the second staining sequence, mouse anti-rat ED1 monoclonal antibody (Chemicon International; Temecula, CA) (1:400 dilution) or goat anti-rat fibrinogen antibody (ICN Pharmaceuticals; Aurora, OH) (1:1000 dilution) was applied as second primary antibody. Then sections were incubated with biotinylated secondary antibody (Vector) and streptavidin-FITC (Amersham Pharmacia Biotech; Tokyo, Japan). The sections were mounted with Vectashield. The sections were examined with an Olympus epifluorescence microscope (Provis AX80; Olympus, Tokyo, Japan). To check the specificity of the immunolabeling, the same immunostaining procedure was followed except that preimmune rabbit or goat serum was applied as the primary antibody. No appreciably stained structures were observed.
Results
Jejunum
In the jejunum, group IIA PLA2 mRNA-positive cells were present in the bottom of crypts (Figures 1a and 1b). No significant signal was seen in the crypts in the negative control. Staining of the epithelial cells in the negative control indicated that it was due to the intrinsic alkaline phosphatase (Figure 1c). Group IIA PLA2 immunoreactivity was also observed in the bottom of crypts (Figures 2a-2c). These cells were pyramidal, with a round or ovoid nucleus (Figures 2d-2f). These characteristic features indicated that they were Paneth cells.
Spleen
ISH revealed that signals of group IIA PLA2 mRNA were observed in large cells (~30–40 μm) with multilobular nuclei in the red pulp (Figures 3a and 3b). Group IIA PLA2-positive cells were roughly spherical but often had blunt, irregular pseudopods on their surfaces. Their nuclei were extraordinarily elaborate, with multiple lobes of various sizes. The double-labeling study demonstrated that group IIA PLA2-positive cells were also positive for fibrinogen (Figures 4a-4f). These morphological and immunohistochemical findings indicated that group IIA PLA2-positive cells in the spleen were megakaryocytes. Neither group IIA PLA2 mRNA nor immunoreactivity was observed in other types of cells in the spleen.
The ED-1 immunohistological study revealed that ED1-positive cells were found in the red pulp. These cells were ~10 μm in diameter, mononuclear, and rich in cytoplasm. Therefore, these ED1-positive cells in the red pulp appeared to be macrophages. A double-immunolabeling of group IIA PLA2 and ED1 revealed that group IIA PLA2 and ED1 were never co-localized in any cells. Thus, group IIA PLA2-positive cells were completely separated from ED1 positive cells (Figures 5a-5f). These results indicated that megakaryocytes, but not macrophages, contained group IIA PLA2.
Other Organs
Group IIA PLA2 mRNA and immunoreactivity were not detected in any cells of the lung, liver, kidney, and pancreas.
Discussion
In this study, using a specific RNA probe and an antibody for rat group IIA PLA2, we demonstrated that group IIA PLA2-producing cells were megakaryocytes in the spleen and Paneth cells in the small intestine in normal rats. The antibody used in the present study was raised against group IIA PLA2 isolated from the rat platelet (Nakano et al. 1990). Although crossreactivities of this antibody with other groups of PLA2s were not examined, only those cells that expressed group IIA mRNA showed a positive reaction with this antibody. It has been reported that group V PLA2 produced by a murine P388D1 macrophage has crossreactivity with the antibody raised against human synovial PLA2, a group IIA enzyme (Balboa et al. 1996). The amino acid sequence identities of mouse IIA PLA2 with other types of PLA2 are 31–51% (Valentin et al. 1999). The overall amino acid identity of rat group IIA and group V PLA2 was reported to be 42.5% (Chen et al. 1994). Because the RNA probe used in the present study covers about 90% of the total mRNA of group IIA PLA2, it is unlikely that this probe detects mRNAs of other groups of PLA2.
IHC of smeared bone marrow cells indicated that megakaryocytes produced group II PLA2 (Murakami et al. 1990). Inada et al. (1991b) tried to identify the cellular origin of group II PLA2 by separating spleen cells on a density gradient. They found that about one third of the cells in the monocyte/macrophage fraction were immunopositive for group II PLA2. Although about 70% of cells in the monocyte/macrophage fraction were of macrophage nature, i.e., immunopositive for ED1, it was not shown that megakaryocytes were not included in the remaining 30% of cells in this fraction. To solve this problem, we performed a double staining of the spleen with both antibodies and found that no group II PLA2 protein was present in the macrophages of the spleen.
The concentration of group II PLA2 was very low in human spleen (Murakami et al. 1988; Nevalainen and Haapanen 1993). Immunohistochemical study failed to detect any group II PLA2 immunopositive cells in the human spleen (Kallajoki and Nevalainen 1997). The spleen of rodents, unlike that of human, is rich in megakaryocytes. This can explain the quantitative difference of group II PLA2 in the spleen between rat and human.
Northern blotting analysis (Ishizaki et al. 1989) indicated the presence of group II PLA2 mRNA in normal rat small intestine, but the cellular origin was not identified. In the human intestine, group II PLA2 was detected in Paneth cells by both IHC and ISH (Nevalainen et al. 1995). The present study confirmed the production of group IIA PLA2 in Paneth cells in rats. The function of PLA2 in Paneth cells is not fully understood. It is believed that they are involved in the host defense because they produce lysozymes and PLA2, both of which have antibacterial properties (Sandow and Whitehead 1979; Wright et al. 1990). Recently, it was shown that group IIA PLA2-deficient mice were susceptible to multiple intestinal adenomas (MacPhee et al. 1995). Therefore, group IIA PLA2 may modulate cell proliferation directly by acting on tumor cells or indirectly by removing potentially harmful dietary fatty acids or bacteria.
Because group II PLA2-specific activities, protein, or mRNA were present in the homogenate of the normal rat liver (Inada et al. 1991b; Dong et al. 1997), kidney (Hara et al. 1995), lung (Murakami et al. 1988; Ishizaki et al. 1989), and pancreas (Lauritzen et al. 1994), we tried to identify group IIA PLA2-producing cells in these tissues by IHC and ISH. However, we could not detect group IIA PLA2 protein and mRNA in normal rat liver, kidney, lung, and pancreas. One explanation for this discrepancy is that the amounts of group II PLA2 protein and mRNA in these organs were much less than those of the spleen (Murakami et al. 1990; De Windt et al. 1997). Therefore, the expression of group IIA PLA2 in these tissues may be too low to be detected by our methods. Another possibility is that group IIA PLA2 protein or mRNA in the megakaryocytes present in the circulation (Kaufman et al. 1965) may have been detected in the previous studies. In this study, we removed the intravascular cellular elements by perfusing blood vessels with Tyrode's solution. In view of the diversity of PLA2s, further studies are necessary to identify the cellular origin of each subtype of this enzyme.
In conclusion, under normal conditions group IIA PLA2-producing cells are splenic megakaryocytes and intestinal Paneth cells in rats. The induction of group IIA PLA2 in other types of cells under inflammatory conditions awaits further studies.
Footnotes
Acknowledgements
Supported by a research grant for intractable pancreatic disease from the Ministry of Health and Welfare in Japan and by a grant from Pancreatic Research Foundation of Japan.