
Abstract
The clinical importance of carbamoyl phosphate synthase I (CPSI) relates to its capacity to metabolize ammonia, because CPSI deficiencies cause lethal serum ammonia levels. Although some metabolic parameters concerning liver and intestinal CPSI have been reported, the extent to which enterocytes contribute to ammonia conversion remains unclear without a detailed description of its developmental and spatial expression patterns. Therefore, we determined the patterns of enterocytic CPSI mRNA and protein expression in human and rat intestine during embryonic and postnatal development, using in situ hybridization and immunohistochemistry. CPSI protein appeared during human embryogenesis in liver at 31–35 e.d. (embryonic days) before intestine (59 e.d.), whereas in rat CPSI detection in intestine (at 16 e.d.) preceded liver (20 e.d.). During all stages of development there was a good correlation between the expression of CPSI protein and mRNA in the intestinal epithelium. Strikingly, duodenal enterocytes in both species exhibited mosaic CPSI protein expression despite uniform CPSI mRNA expression in the epithelium and the presence of functional mitochondria in all epithelial cells. Unlike rat, CPSI in human embryos was expressed in liver before intestine. Although CPSI was primarily regulated at the transcriptional level, CPSI protein appeared mosaic in the duodenum of both species, possibly due to post-transcriptional regulation.
Keywords
C
In addition to CPSI, which encodes both liver and intestinal mitochondrial CPSI, there are two heterologous enzymes with CPS activity (EC 6.3.4.16), CPSII and III. Of these, only CPSI and II are expressed in mammals, whereas CPSIII has only been described in elasmobranchs (McCudden and Powers–Lee 1996). Moreover, in contrast to CPSI, which is mitochondrially localized, CPSII is a cytoplasmic enzyme that is mainly important in de novo polyamine synthesis, is practically absent in both enterocytes and hepatocytes, and is not recognized by the antiserum used in this study to detect CPSI.
We describe here the distribution of human and rat CPSI during development in the pre- and postnatal intestine. We performed immunohistochemistry (IHC) to localize CPSI protein and we used rat and human CPSI cDNA fragments as probes in in situ hybridization (ISH) to localize the respective CPSI mRNAs. We performed immunoelectron microscopy to study CPSI protein localization in the intestine at the subcellular level. Thus, we were able to give a detailed description of the regulation of the developmental expression patterns of human and rat CPSI in the intestine.
Materials and Methods
Tissue
Human tissue included embryonic samples of 31–35, 35–38, 44–51, 59, 62, 63, 64, 68, and 72 embryonic days (e.d.), according to Carnegie staging (Dingemanse and Lamers 1994b). Furthermore, duodenal biopsies (distal to the papilla of Vater) were taken with informed consent of the patients and/or their parents as well as with permission of the ethical committee of our institution. Patients described underwent endoscopy for various gastrointestinal complaints. For each patient, age, racial origin, clinical symptoms, and medical history were recorded. Duodenal biopsies were analyzed from 64 patients representing all stages of infancy and childhood (3 months to 18 years). Furthermore, eight normal proximal jejunal sections were obtained as resections from adult humans who underwent surgery for distant tumors, mostly of the pancreas. Rat tissue was obtained with permission of the Animal Research Committee of our institution and included stomach, duodenum, jejunum, and colon from gestational days 9, 13, 14, 16, 18, 20, postnatal days 0, 4, 8, 14, and 28, and from older rats at 5 months.
Cloning of the Human CPSI Probe
We cloned 1413
RNA Isolation and CPSI mRNA Quantification
CPSI mRNA was quantified as described earlier (Van Beers et al. 1997). In brief, RNA was isolated from human biopsies, and CPSI mRNA was quantified by spot-blotting, using a 1395 nt CPSI probe transcribed from the linear CPSI cDNA fragment described above. Quantitative autoradiographs were prepared with a PhosphorImager using ImageQuant software (Molecular Dynamics; Sunnyvale, CA). Membranes were stripped of adherent CPSI probe and checked for absence of radioactivity by exposing PhosphorImager screens for 48 hr. Sequentially, all the blotted RNA samples were incubated with a 1-
Immunohistochemistry
Unless otherwise indicated, tissue was fixed in 4% paraformaldehyde (PFA), essentially as previously described (Van Beers et al. 1995a). Seven-μm-thick sections from 30 min 4% PFA-fixed, paraffin-embedded tissue were deparaffinized and incubated for 30 min in 3% H2O2 in PBS to inactivate endogenous peroxidase activity. All sections were incubated for 30 min in 10 mM Tris-HCl, 5 mM EDTA, 150 mM NaCl, 0.25% gelatin, 0.05% Tween-20 to reduce background. The sections were subsequently incubated overnight with adequate dilutions in PBS of the primary antibodies: 1.5 hr with rabbit anti-mouse serum (1:7500; Dako, Glostrup, Denmark), 1.5 hr with goat-anti-rabbit serum (1:1000; Dako), and 1.5 hr with rabbit peroxidase-anti-peroxidase (1:1000; Dako) at room temperature. Anti-lactase (HBB 1/90/ 34/74) and anti-SI (HBB 2/219/20 and HBB 2/219/88) monoclonal antibodies were diluted 1:1000 from ascites (both antibodies were a kind gift of Dr. H.-P. Hauri). A polyclonal anti-rat sucrase–isomaltase serum (a gift of Dr. K.Y. Yeh) was used 1:500 to detect rat sucrase–isomaltase. Rabbit anti-human intestinal mucin (MUC2) polyclonal serum was diluted 1:1000 as described previously (Tytgat et al. 1994), and rabbit anti-human chromogranin A was used, diluted 1:1000 (Dako). The CPSI-monospecific polyclonal antiserum raised against rat CPSI and crossreacting with human CPSI was obtained from Dr. R. Charles (Charles et al. 1980).
Finally, as controls, in the case of duodenal sections that consistently exhibited mosaic staining on PFA-fixed tissue, duplicate biopsies were used to test the effect of other fixation protocols, i.e., methanol/acetone/H2O (40:40:20) fixation with or without subsequent boiling for 5 min in sodium citrate buffer 0.1 M (pH 6.0) or IHC on unfixed cryostat sections.
Ultracryotomy and Immunolabeling
Freshly excised tissue blocks were fixed with 2% PFA, 0.2% glutaraldehyde in 0.1 M phosphate buffer, pH 7.4 (PB) for 2 hr and stored in 2% PFA in PB. The blocks were washed three times with PBS, 0.15 M glycine, and finally embedded in 10% gelatin in PB. The gelatin was allowed to solidify and small cubic blocks were cut at 4C and infused with 2.3 M sucrose in PB for at least 2 hr at 4C. The blocks were mounted on a copper specimen holder and frozen in liquid nitrogen. Ultrathin cryosections were prepared at −120C on a Leica Ultracut S (Vienna, Austria) using a Drukker diamond knife (Drukker International; Cuick, The Netherlands) according to Liou et al. (1996). The sections were immunolabeled according to Slot et al. (1991) with anti-CPSI antiserum (diluted 1:100 in PBS/1% bovine serum albumin) and protein A–gold 10 nm.
In Situ Hybridization
Essentially as described earlier (Rings et al. 1994), 35S-labeled DNA probes used in ISH on rat tissue were synthesized using random priming from 2386 kb of rat CPSI cDNA, 1000-BP rat albumin cDNA, or 1800-
Succinate Dehydrogenase (SDH) Activity
Cryosections were thawed, treated with acetone for 20 min, and incubated at 37C for 60 min in a solution containing 40 mM PB, pH 7.4, 40 mM sodium succinate, 0.8 mg/ml nitroblue tetrazolium, 0.125 mM CaCl2, 0.2 mM AlCl3, 25 mM NaHCO2, and phenazine methosulfate (N-methyldibenzopyrazine) 0.5 mg/ml. After staining, sections were fixed in 4% PFA in PB and coverslips were mounted for microscopy. Dark precipitate indicated SDH enzyme activity.
Results
Rat CPSI Expression During Development
To analyze the developmental onset of CPSI gene expression, we performed ISH on embryonic rat sections. CPSI mRNA was undetectable in the embryos of 9–16 e.d. (not shown). CPSI mRNA is very abundant in intestinal loops at 18 e.d., but almost, if not entirely, absent from liver (Figure 1A). Lactase mRNA was also expressed in these intestinal cross-sections at 18 e.d. (Figure 1B). At 20 e.d., in addition to intestine, CPSI mRNA was also expressed in liver (Figure 1C). In control experiments, lactase mRNA was detected exclusively in intestine at 20 e.d., whereas albumin was detected exclusively in liver (Figures 1D and 1E). During embryonic development in rat, CPSI mRNA and protein were always simultaneously detected in the same tissues (IHC not shown).
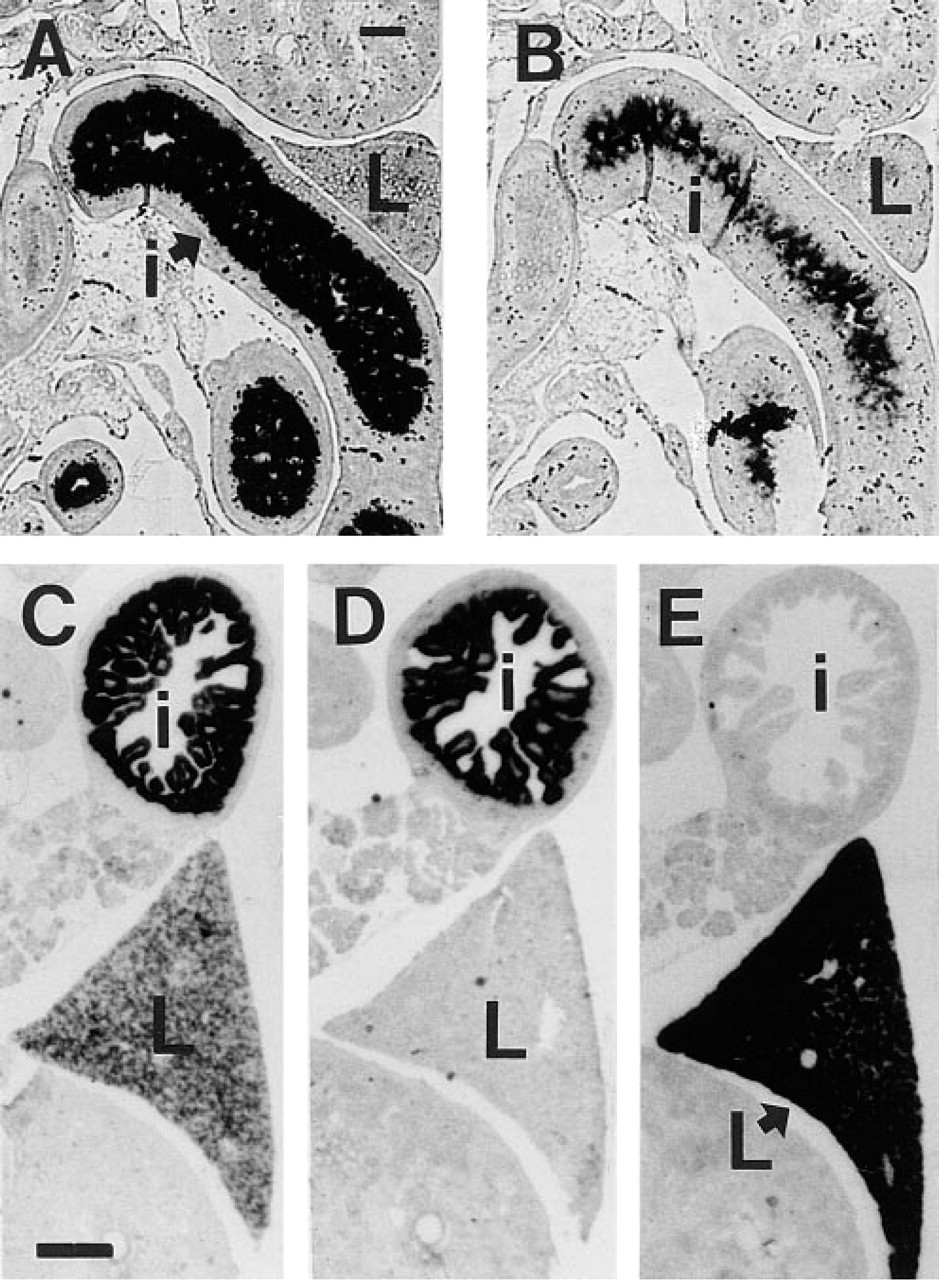
In situ hybridization using random primed single stranded 35S-labeled cDNA probes on sections of rat embryos at 18 (
To analyze the spatial arrangement of CPSI mRNA and protein expression in more detail, we studied the crypt–villous axis of rat jejunum during postnatal development. Immediately after birth, the epithelium consists of short villi and intervillous regions, but crypts have not yet formed. Between 0 and 28 days, villous length increased and crypt formation occurred. CPSI protein was present throughout the entire epithelium at all stages of development, although at later stages (28 days and older) the intensity of the staining declined from the crypt–villous junction towards the villous tip (Figures 2A-2C). Concurrent with the morphological changes, the CPSI mRNA expression pattern changed. At birth, CPSI mRNA was detected in the entire epithelium, whereas expression was confined to the crypts at 28 days (Figures 2D-2F). The findings presented for 28 days were representative for adult animals up to 5 months (not shown) and therefore reflect the adult CPSI mRNA and protein patterns.
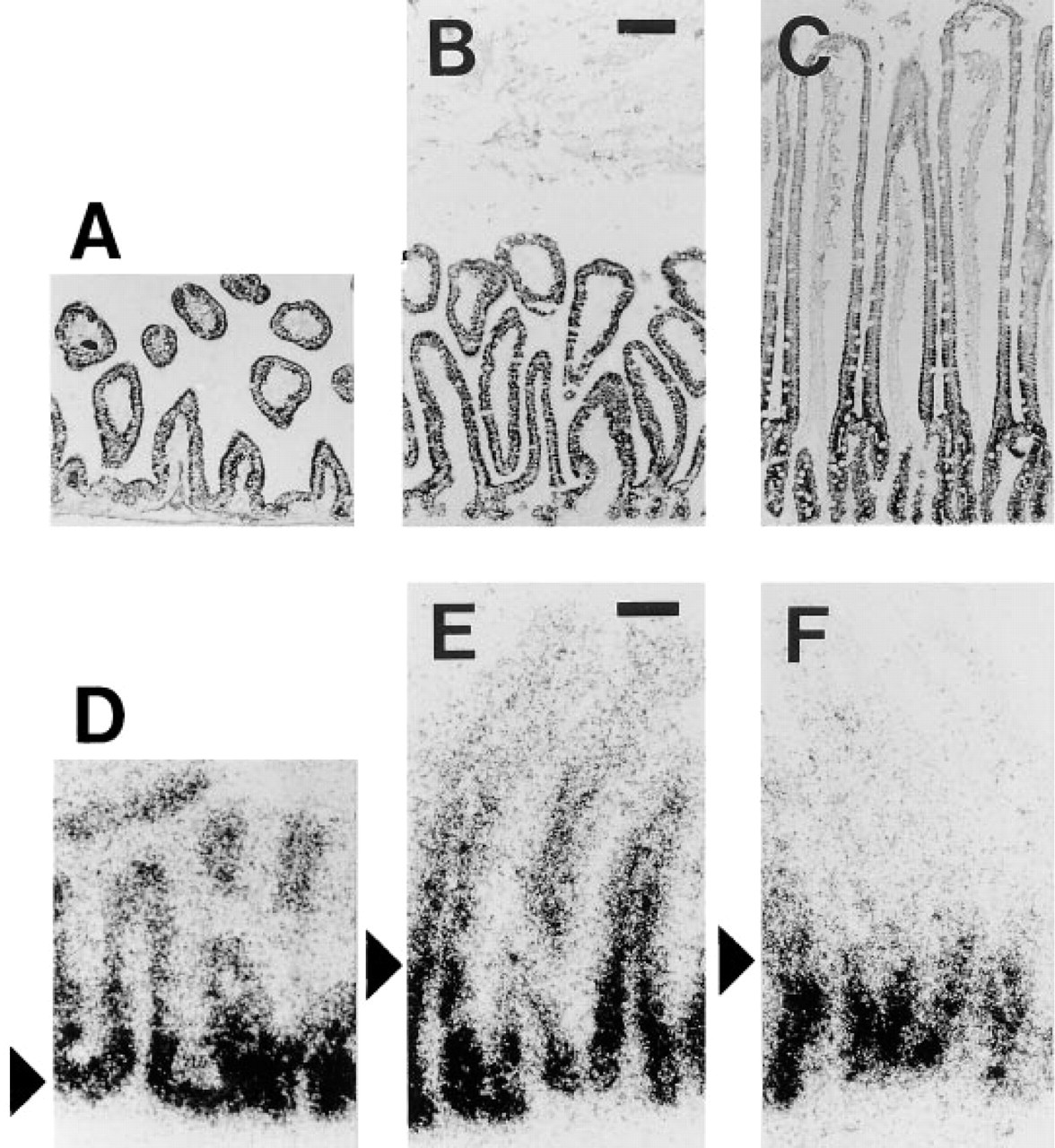
Immunohistochemistry for CPSI on sections of rat jejunum at 0 (
Human CPSI Expression During Development
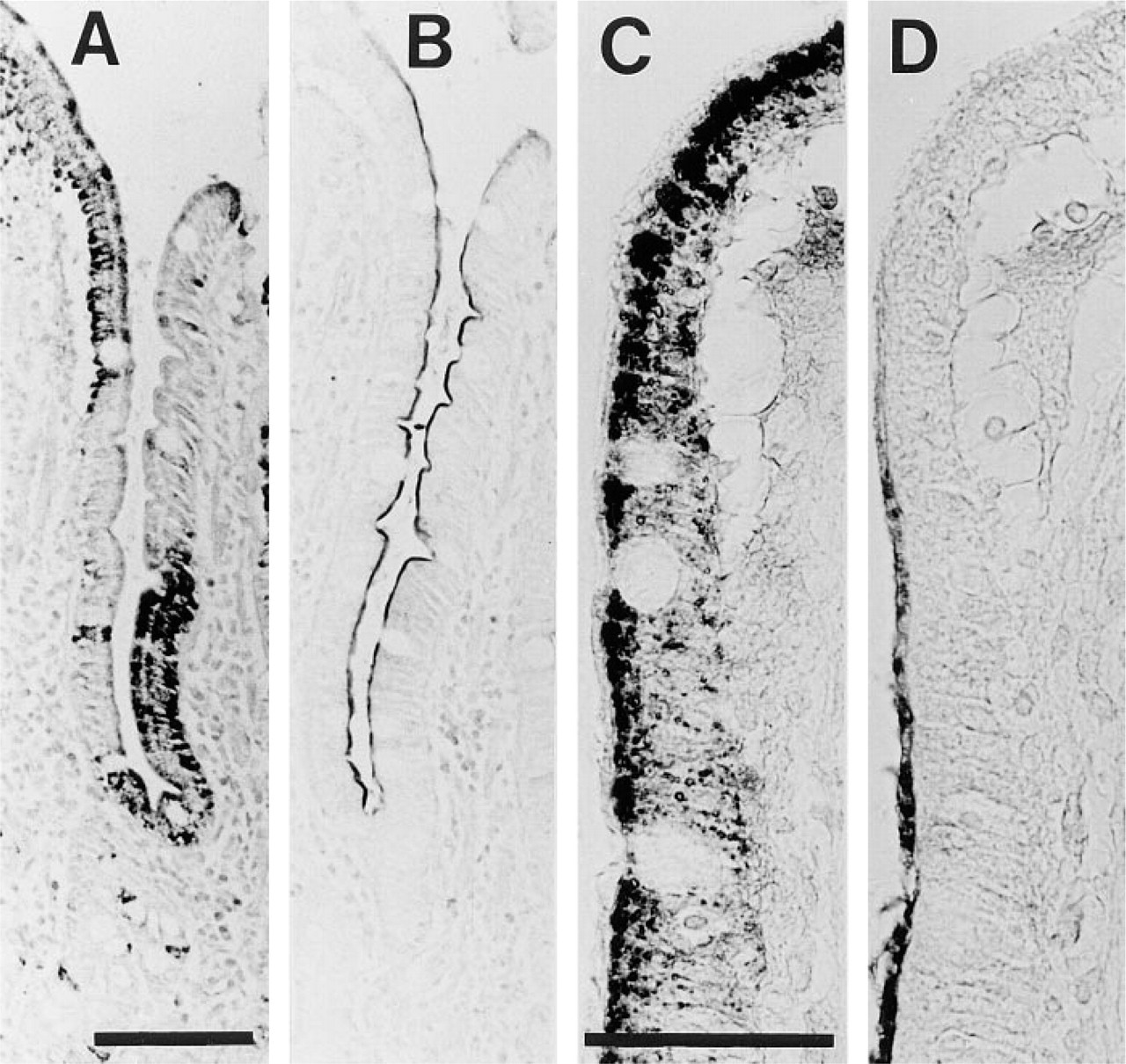
We analyzed CPSI protein expression in human developmental stages (Figure 3). The earliest detection of CPSI protein was in liver at 31–35 days of gestation, whereas at that stage CPSI was absent from all other tissues, including intestine (Figure 3A). At 59 days of gestation, in addition to liver, CPSI was abundantly detected in both villous and intervillous epithelium of the small intestine (Figure 3B). The protein remained highly expressed in villous enterocytes of the developing small intestine at 72 days of gestation as well as in the enterocytes of villi and crypts of the jejunum of adults (Figures 3D and 3E). CPSI protein was not detected in human stomach or in sigmoid biopsy sections (not shown).
Immunohistochemistry for CPSI on human tissue. (
Mosaic CPSI Protein Distribution in Human Duodenal Epithelium
Duodenal biopsies from a large number of human subjects between 3 months and 18 years old were analyzed for CPSI mRNA and protein expression. To assess the distribution of CPSI mRNA along the crypt–villous axis of the normal human duodenum, we performed ISH. Figure 4 shows a representative example from an ISH experiment that included duodenal specimens from four different individuals. It shows CPSI mRNA in most if not all enterocytes along the crypt–villous axis. Note, however, the absence of staining in the Brunner's gland epithelium, which is extensively present in the duodenal mucosa.
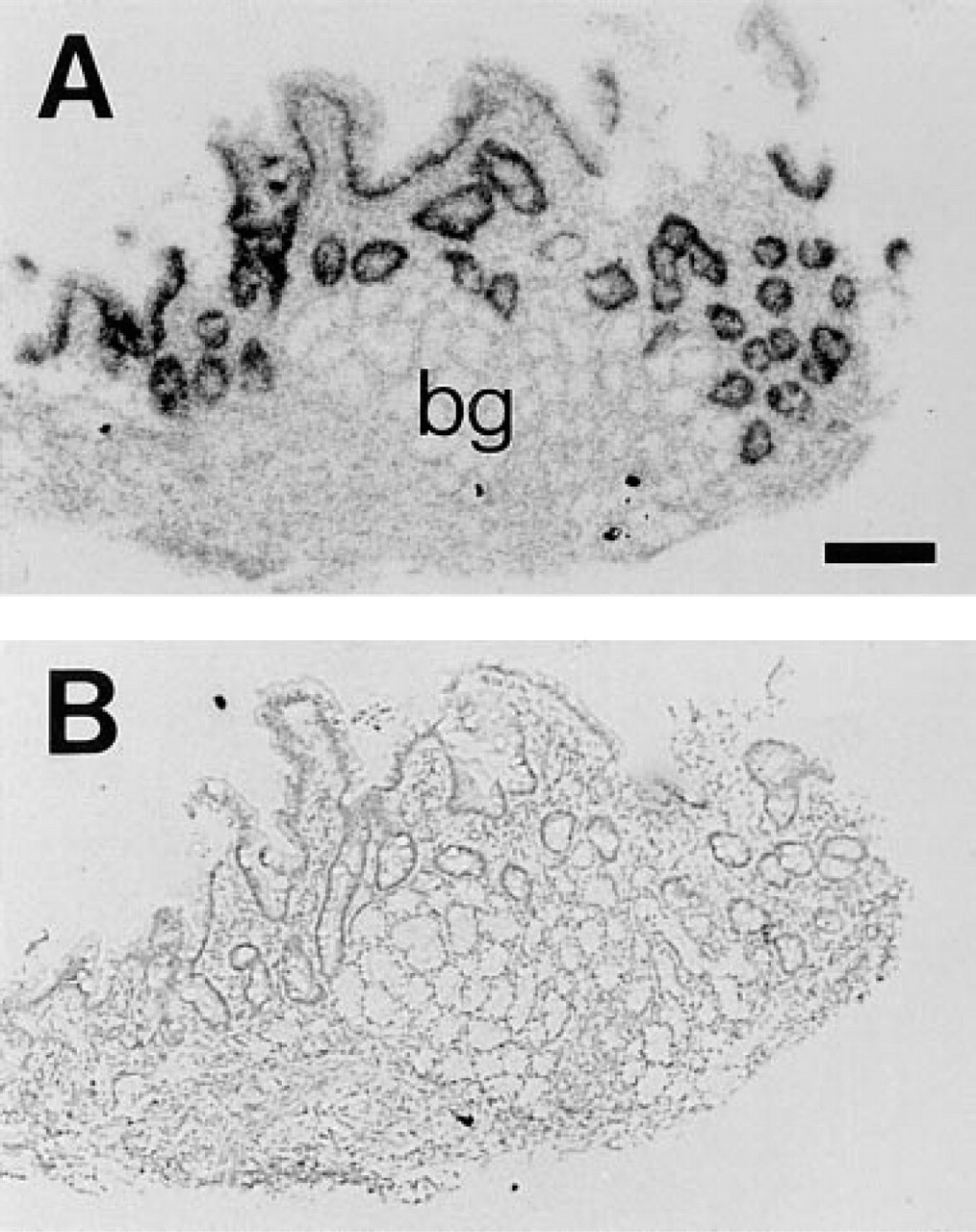
In situ hybridization using in vitro-transcribed single-stranded 35S-labeled human CPSI riboprobes on sections of healthy human duodenum. (
In sharp contrast to the mRNA expression, we found a mosaic pattern for CPSI protein, whereas lactase, another enterocytic enzyme, appeared uniformly expressed in adjacent sections (Figures 5A and 5B). This mosaic expression pattern was not observed in fetal tissue and did not change during post-natal development, because similar patterns of expression were observed in duodenal biopsies from all 64 individuals aged 3 months to 18 years. Moreover, we found similar results in individuals from various racial backgrounds, as well as in individuals with or without affected duodenal histology (not shown). Furthermore, this mosaicism did not depend on tissue fixation, because PFA or methanol–acetone fixation yielded very similar results (not shown). This pattern also was not affected by boiling of the tissue sections in sodium citrate buffer (not shown). In addition, IHC on unfixed duodenal cryosections revealed mosaic CPSI protein expression (not shown). In contrast to duodenum, CPSI protein was uniformly present in all jejunal enterocytes along the crypt–villous axis, whereas lactase protein levels declined towards the tip of the villi in adjacent sections (Figures 5C and 5D).
Immunohistochemistry on sections of healthy human duodenal biopsy (
We compared the areas of CPSI-negative cells in the epithelium with the spatial arrangement of nonenterocytic cell types. We used anti-human MUC2 antiserum to label all goblet cells and anti-human chromogranin A antibody to label all enteroendocrine cells. Specific Paneth cell staining was not performed because Paneth cells reside only at the base of the crypts and could never contribute to CPSI mosaicism among villous cells. The numbers and distributions of the goblet and enteroendocrine cells were very different from the CPSI-positive areas in the epithelium. Goblet cells and enteroendocrine cells were very scarce, whereas the CPSI-positive areas comprised about half of the epithelium (not shown).
We further analyzed the spatial arrangement of CPSI-expressing cells using various planes of sectioning. In all human duodenal biopsies sectioned along the crypt–villous axis as in Figure 5, CPSI-positive cells were mosaically arranged. In contrast, biopsies sectioned through the plane perpendicular to the length of the crypts often showed clusters of CPSI-positive crypts (Figure 6).
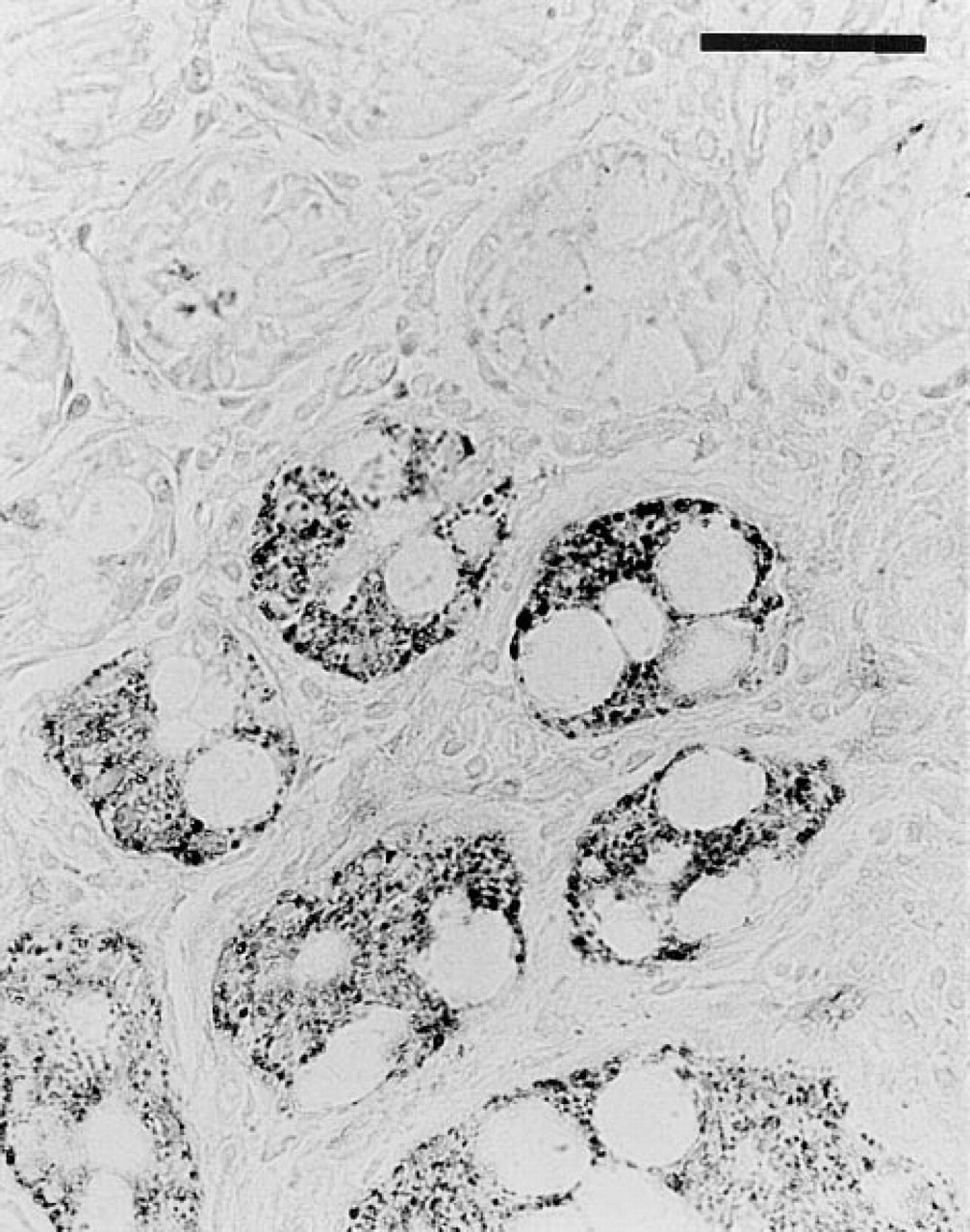
Immunohistochemistry for CPSI on a human duodenal biopsy section along the plane of the upper crypts. Bar =100 μm.
All Human Duodenal Enterocytes Contain Functional Mitochondria
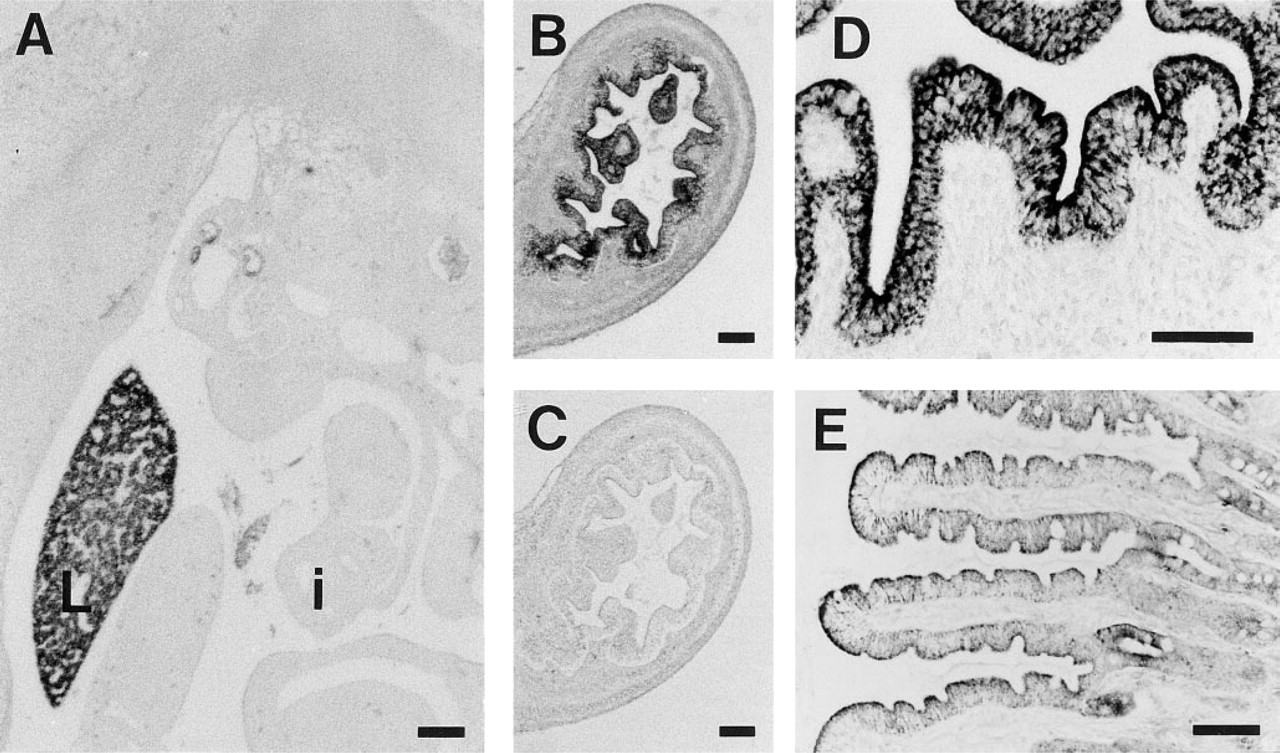
SDH enzyme activity was studied by enzyme histochemistry on frozen sections of human duodenum, as a hallmark for mitochondrial function. In contrast to the mosaic CPSI protein expression, all epithelial cells displayed similar SDH activity (Figure 7). At higher resolution with transmission electron microscopy (TEM), all enterocytes were found to possess comparable numbers of morphologically identical mitochondria. By immunogold labeling, CPSI was found in the mitochondria of some but not all enterocytes. Many examples were found in which all mitochondria of an enterocyte were labeled for CPSI, whereas the mitochondria of a directly neighboring enterocyte were completely devoid of CPSI labeling (Figure 8A). Using TEM, it was also demonstrated that CPSI expression was absent from goblet cells (Figure 8B).
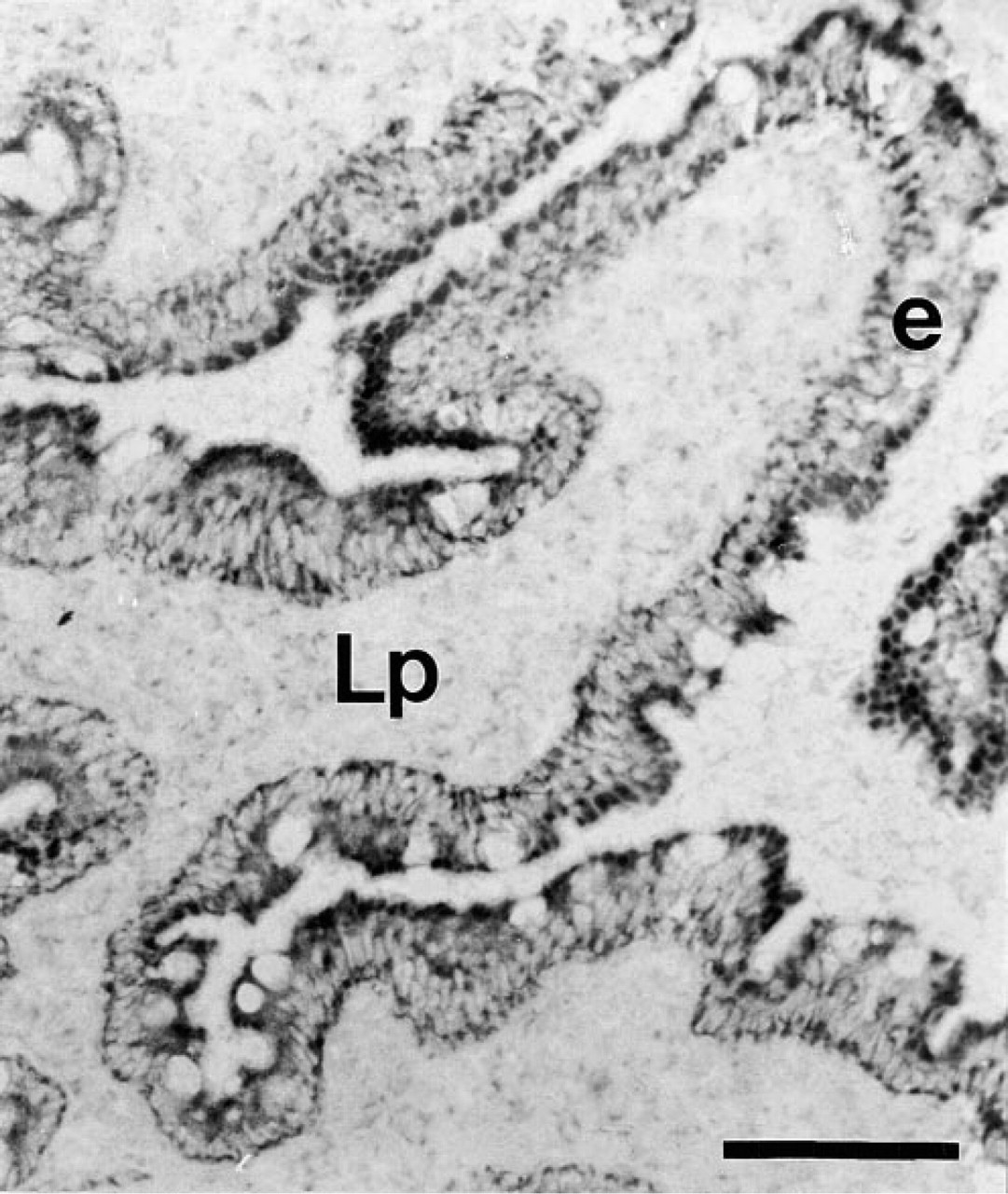
Succinate dehydrogenase activity was assayed by enzyme histochemistry, indicative for mitochondrial function. A representative duodenal section of one of seven patients is shown. Dark staining was extensive and continuous throughout the entire epithelium (e) and to a much lesser extent in the lamina propria (Lp). Bar = 100 μm.
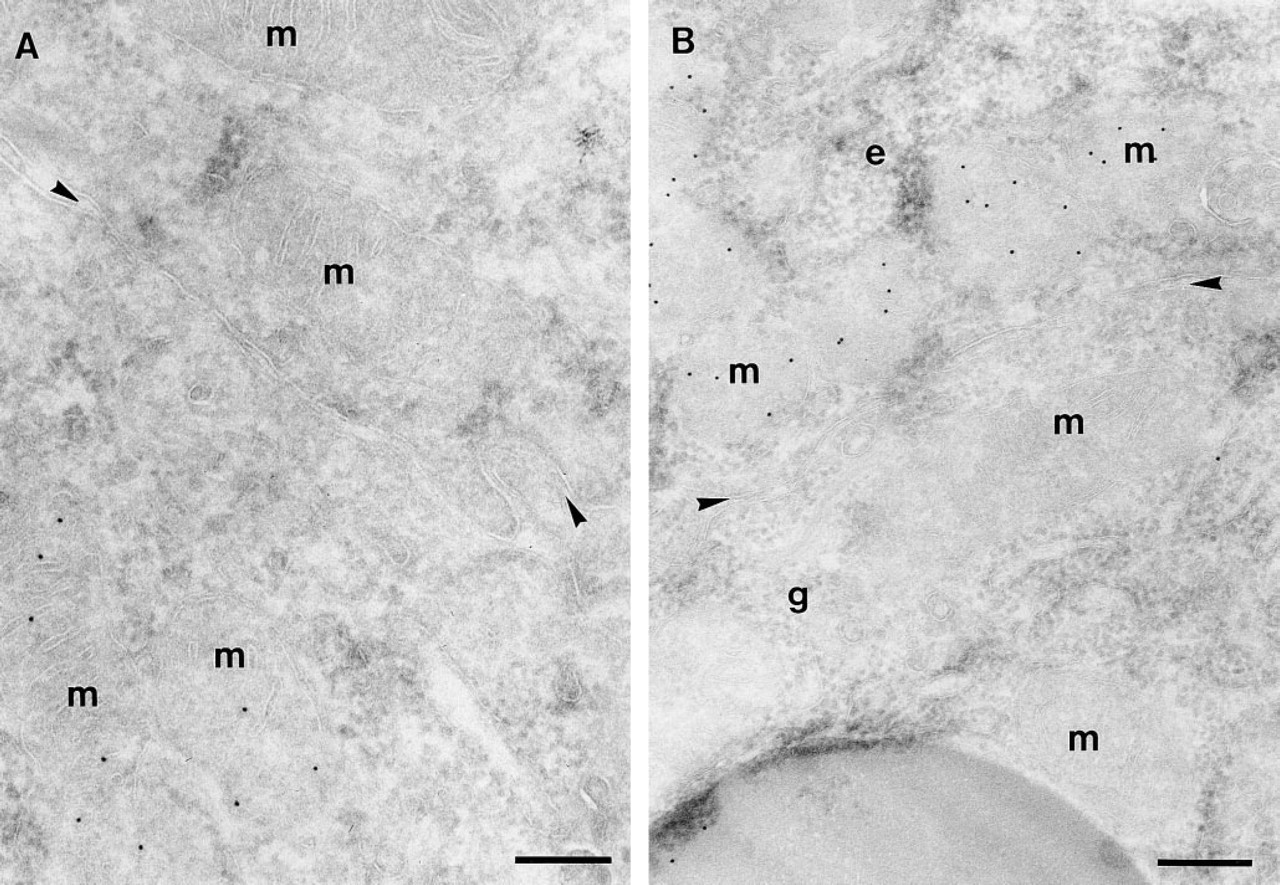
Ultrathin cryosections of human duodenum labeled with anti-CPSI and protein A–gold (10 nm). (
Mosaic CPSI Distribution in Rat Duodenal Epithelium
In rat we found a mosaic CPSI expression pattern similar to that of human duodenum (Figure 9A). This mosaic expression pattern was not observed in fetal tissue and could be demonstrated at all postnatal stages examined. As expected, sucrase–isomaltase was uniformly present in the brush borders of all duodenal villous enterocytes (Figure 9B). With respect to the longitudinal distribution in the small intestine, CPSI protein appeared mosaic only within 1 cm directly distal to the pylorus. Beyond this small region, CPSI was uniformly expressed at high levels in all enterocytes. ISH studies showed uniform CPSI mRNA distribution among all duodenal crypt enterocytes, similar to the CPSI mRNA in jejunum (not shown).
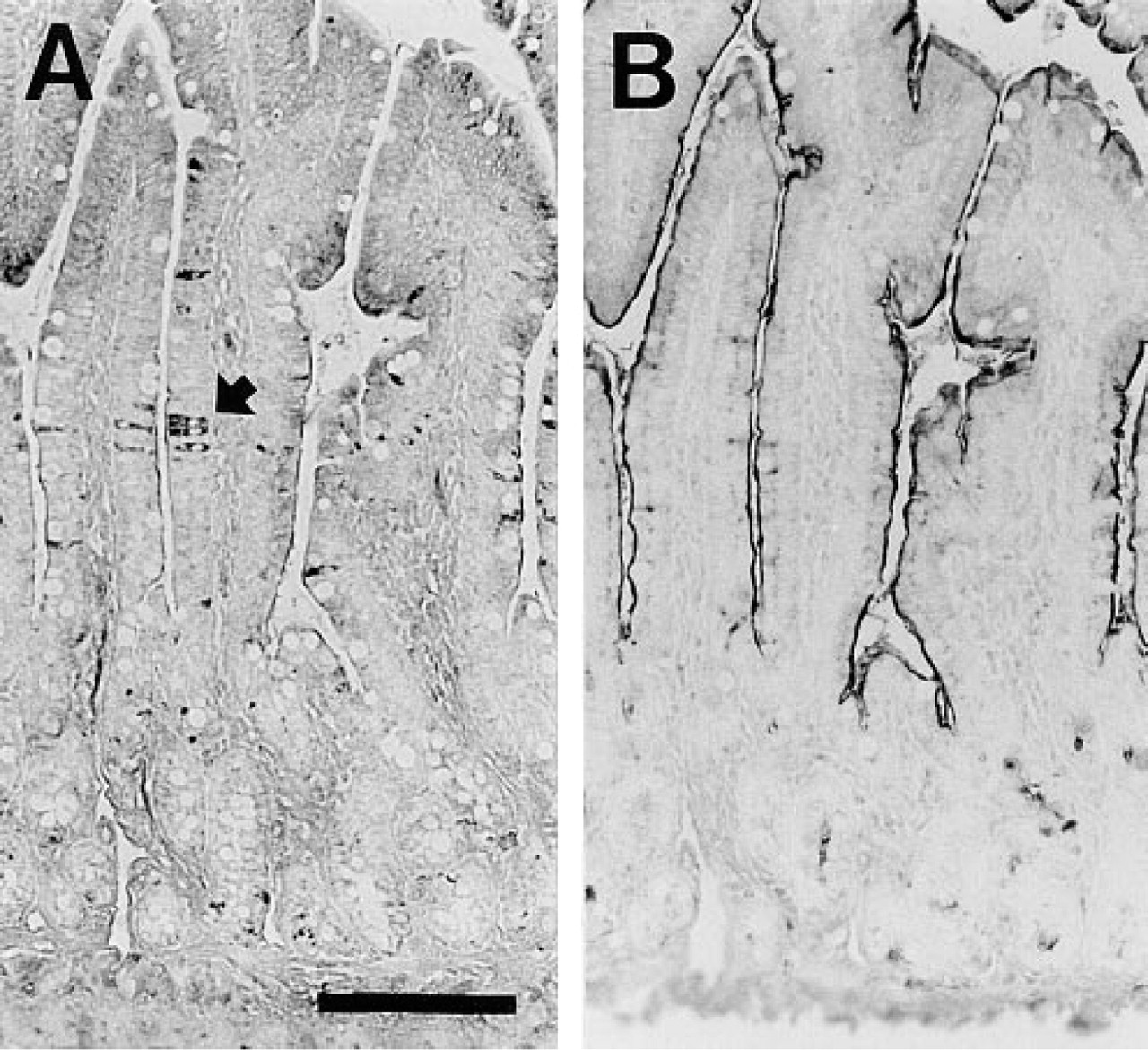
Immunohistochemistry on adult rat duodenum of the first several villi distal of the pylorus. (
Levels of CPSI mRNA in Human Duodenum Increased with Age
Among the duodenal biopsies studied, the expression patterns of CPSI protein or mRNA did not change at the histological level, as described above. However, in addition to histological examination, we performed a quantitative analysis of CPSI mRNA levels in duodenal RNA samples from 22 healthy white subjects. A Spearman rank correlation analysis revealed a significant increase in CPSI mRNA levels with age up to 12 years (n = 22, r = 0.43, p = 0.008). A similar analysis on biopsies from a more proximal location, from the bulbus duodeni, yielded comparable results (n = 24, r = 0.63, p = 0.0009).
Discussion
CPSI Expression During Development
The earliest expression of CPSI mRNA and protein in rat intestine was at 18 e.d. (i.e., 86% of gestation) and in liver at 20 e.d. (95% of gestation). These results are in agreement with findings on Northern blots by Ryall et al. (1986). Before and immediately after birth, there is complete coincidence between the intestinal cells expressing CPSI mRNA and protein. During later stages of postnatal development, CPSI mRNA was localized only in crypts, whereas the protein remained present in all villous enterocytes. This distribution is in accordance with the half-lives of the CPSI mRNA (4–6 hr) compared to the CPSI protein (4–5 days) (Dingemanse and Lamers 1994a).
In human, the first appearance of CPSI protein was relatively early in gestation compared to rat. We detected human CPSI protein in liver at 31–35 e.d. (12% of gestation) before detection in intestine at 59 e.d. (22% of gestation). This species difference may be due to the relatively long gestation in humans. This may already necessitate catabolic processes, including amino acid conversion through the urea cycle, before birth, whereas in rat anabolism may dominate until birth and the urea cycle is not needed until approximately the time of birth.
CPSI Protein, but not Its mRNA, Is Mosaically Distributed in the Duodenum
A mosaic pattern of expression can be defined as a seemingly random distribution among cells for a certain phenotypic trait. The localization of CPSI protein in human and rat duodenum consistently appeared mosaic at the light and electron microscopic levels. The causal mechanism remains obscure, but three explanations could be excluded. (a) This phenomenon is very likely not correlated with the presence of different cell types within the epithelium. Neither the numbers nor the localizations of goblet, Paneth, and entero-endocrine cells appear to correlate with CPSI-positive or negative areas. Moreover, enterocyte markers, such as lactase and sucrase–isomaltase, are expressed in a continuous fashion in the epithelium, whereas the CPSI protein is mosaically expressed by these cells. (b) The SDH activity and TEM studies indicated that neither distribution, number, function, nor morphology of mitochondria varied among human duodenal enterocytes. The absence of CPSI protein is therefore not due to absence of functional mitochondria. (c) Expression of CPSI protein in duodenum is independent of the position of the enterocytes along the crypt–villous axis. This strongly suggests that cellular differentiation along this axis does not determine CPSI protein expression, as opposed to that of many other intestinal proteins, such as lactase and sucrase–isomaltase (Van Beers et al. 1995b).
For CPSI mRNA, we found no evidence of mosaic expression in either human or rat duodenum, suggesting a post-transcriptional mechanism causing the CPSI protein mosaicism. Because the mRNA appears to be expressed in all enterocytes, there are three possible explanations for the absence of CPSI protein in the mitochondria of some cells: (a) the CPSI mRNA is not translated; (b) CPSI protein is not imported into mitochondria but degraded instead; or (c) CPSI protein is degraded rapidly and selectively after its import into mitochondria. At present, we have no means to distinguish among these possibilities.
Comparison with Other Duodenal Mosaicisms
It is important to note that the region of the intestine at which the CPSI mosaicism occurs is relatively small. In most of the small intestine of rat and human the expression of CPSI protein and mRNA is continuous. Nevertheless, the phenomenon is very interesting from a cell biological point of view and may add to the insight into intestinal cell migration, stem cell hierarchy, and differentiation programs of enterocytes.
The duodenal CPSI protein mosaicism was a novel finding. Earlier, we showed mosaicism for lactase and sucrase–isomaltase in developing rat intestine (Rings et al. 1994). This study strongly suggested that mosaic gene expression reflects a local phenomenon along the longitudinal axis, as a transition between regions with and without expression of a certain gene. The position of the mosaicism appears to be guided by the underlying stem cells, because these cells are the only epithelial cells to contain and perpetuate the necessary positional information with respect to the position of the enterocytes along the longitudinal axis.
Maiuri et al. (1991,1992,1994) showed mosaic expression in the intestine for lactase in human, rat, and rabbit. The mosaic expression of human lactase was distinctly different from the CPSI mosaicism, because not only the protein but also the lactase mRNA appeared mosaic in the epithelium. These authors concluded that, for human lactase biosynthesis, a high degree of variation exists in transcriptional and post-transcriptional regulation among enterocytes within a single individual. An unrelated mosaicism in human intestine was found, displaying heterogeneity among enterocytes in the expression of a blood group antigen (Maiuri et al. 1993).
We hypothesize that CPSI protein mosaicism is mainly explained by differences among intestinal stem cells that are reflected in their daughter cells. It has been shown unequivocally that mouse intestinal crypts become increasingly more homogeneous until only monoclonal crypts were detected at 7 days post partum (Hermiston and Gordon 1995). Interestingly, these studies indicated that even with exclusively but different monoclonal crypts mosaicisms still occur for any phenotypic trait on villi, owing to the fact that enterocytes derive from multiple crypts (up to 12 in mice) and together populate the same villus. This leads to stretches, or ribbons, of phenotypically different enterocytes on villi. Because only two-dimensional analyses were performed on duodenal CPSI patterns, our conclusions regarding the presence of interrupted or uninterrupted stretches of CPSI-positive cells are somewhat limited. Nevertheless, we can limit our possible explanations for mosaic CPSI expression to two options: (a) the stem cells in the crypts of the duodenum remain polyclonal with respect to CPSI expression; or (b) both CPSI-positive and CPSI-negative monoclonal crypts are present throughout the length of the duodenum, supporting the continuous generation of a mixture of CPSI positive and negative enterocytes on villi.
The physiological relevance and the molecular mechanisms that cause these intestinal mosaicisms remain obscure. It is our opinion that these mosaicisms merely reflect transition zones along the cephalocaudal axis between the presence and the absence of CPSI. We do not attempt to attribute intrinsic physiological meaning to these mosaicisms.
Potential Physiological Significance of Intestinal CPSI
Our results show that CPSI is highly expressed in most of the small intestine, most likely during the entire lifespan in human and rat. The physiological role of enterocytic CPSI was demonstrated in pig and rat and appears to be different from that of liver CPSI (Hoogenraad et al. 1985; Blachier et al. 1991,1993; Wu 1995). Adult rat enterocytes produce and release citrulline, whereas the action of liver CPSI leads to conversion of ammonia into urea and no intermediates are released from hepatocytes in significant amounts (Hoogenraad et al. 1985). In humans, enterocytic CPSI could also have a role in the total body nitrogen balance, especially through the consumption of ammonia and its contribution to the intestinal formation of citrulline. The role of intestinal CPSI in the systemic nitrogen balance may be underestimated compared to hepatocyte CPSI. It is conceivable that intestinal and hepatic CPSI work in a concerted fashion to regulate serum ammonia levels, although intestinal CPSI has not previously been proposed for this role. Consequently, in patients with fatal hyperammonemia, a condition that shows peak incidence around birth and adolescence or at 51 years in one case (Johnson et al. 1972; Lo et al. 1993; Wong et al. 1994; Zimmer et al. 1995; Lichtenstein et al. 1997), it would be highly interesting to measure enterocytic CPSI in addition to hepatic CPSI activity rather than evaluating liver biopsies alone. More close studies of the differences between the regulation of liver and intestinal CPSI, or the extent of damage in surgery or disease in either organ, would allow better understanding of the causes of hyperammonemia. More suggestive evidence that intestinal CPSI may significantly contribute to the regulation of serum ammonia homeostasis comes from findings in patients treated with cytostatic drugs, resulting in hyperammonemia in several cases (Fine et al. 1989; Liaw et al. 1993). Unfortunately, in none of these cases was intestinal mucosal damage or CPSI activity assessed, whereas the intestine is much more prone to damage by cytostatic treatment compared to liver. We believe that more attention is warranted to elucidate possible vital roles of the intestine in ammonia metabolism.
Footnotes
Acknowledgements
Supported by Nutricia, the Netherlands (EHVB, HAB) and by the Netherlands Organization for Scientific Research (NWO) (EHHMR).
We thank R. Charles, Dept of Anatomy and Embryology, University of Amsterdam, for his gift of the anti-CPSI polyclonal antiserum, H.P. Hauri, Biocenter, Basel, Switzerland, for his anti-human lactase and sucrase–isomaltase monoclonal antibodies HBB 1/90/34/74, HBB 2/219/20, and HBB 2/219/88, and K.Y. Yeh, School of Medicine, Shreveport, LA, for the polyclonal anti-rat sucrase–isomaltase serum.