
Abstract
The multidrug resistance-associated protein (MRP) that is involved in drug resistance and the export of glutathione-conjugated substrates may not have the same epithelial cell membrane distribution as the P-glycoprotein encoded by the MDR gene. Because intestinal and kidney epithelial cells are polarized cells endowed distinct secreting and absorptive ion and protein transport capacities, we investigated the tissue and cell distribution of MRP in adult mouse small intestine, colon, and kidney by immunohistochemistry. Western blot analyses revealed the 190-kD MRP protein in these tissues. MRP was found in the basolateral membranes of intestinal crypt cells, mainly Paneth cells, but not in differentiated enterocytes. All the cells lining the crypt-villous axis of the colon wall contained MRP. MRP was found in the glomeruli, ascending limb cells, and basolateral membranes of the distal and collecting tubule cells of the kidney but not in proximal tubule cells. Cultured mouse intestinal m-ICc12 cells and renal distal mpkDCT cells that have retained the features typical of intestinal crypt and renal distal epithelial cells, respectively, also possess MRP in their basolateral membranes. The patterns of subcellular and cellular distribution indicate that MRP may have a specific role in the basolateral transport of endogenous compounds in Paneth, renal distal, and collecting tubule cells.
T
Pgp and MRP belong to the ATP-binding cassette family of transporter proteins and both cause concomitant decreases in cellular accumulation of cytotoxic drugs and their increased excretion (Gottesman and Pastan 1993; Cole et al. 1994; Zaman et al. 1994). MRP can protect cells against cytotoxic xenobiotic compounds, but the exact way in which it removes drugs may be different from that of Pgp(s). MRP transports substrates conjugated to glutathione and leukotriene C4, and is involved in inflammation (Jedlitschky et al. 1994; Leier et al. 1994; Muller et al. 1994; Wijnholds et al. 1997). Immunocytochemical studies of cultured cancer cells and normal human tissues and tumors also suggest that MRP may have a distinct membranous or cytoplasmic distribution (Flens et al. 1996). It has been detected in the human intestine, colon, and kidney (Flens et al. 1996), but little is known about the exact cellular location of the protein in these tissues. The intestine and kidney contain several types of cells with specialized transport functions, and MRP may be restricted to particular cell types.
Rather unexpectedly, MRP has been shown to be located in the basolateral membrane of pig kidney proximal-like LLC-PK1 epithelial cells transfected with human MRP, where it increases the transport of S-conjugated glutathione to the basal side of the cells (Evers et al. 1996). MRP has also been found in the basolateral membrane domain of human ciliated bronchial epithelial cells (Brechot et al. 1998). These results suggest that MRP may not necessarily be present in the same membrane domain as Pgp, i.e., the apical brush border of polarized epithelial cells (Thiebaut et al. 1987). We have therefore carried out immunocytochemical experiments on frozen tissue sections from normal adult mice and two mouse trans-immortalized cell lines that have kept the main features of crypt intestinal and renal distal tubule cells to define more precisely the tissue and cell distribution of MRP in the gastrointestinal tract and kidney.
Materials and Methods
Mouse and Human Tissues
Experiments were performed on gastrointestinal and kidney tissues from adult male mice fed a standard diet with free access to tapwater. Specimens of human terminal ileum were obtained from a patient during surgery for colon carcinoma. Pieces of the nontumorous part of a human renal cortex were obtained from a kidney cancer removed by surgery. Samples were rapidly frozen in liquid nitrogen and kept frozen until required.
Cell Lines
We used two immortalized cell lines, one from the small intestine and the other from the kidney of transgenic mice carrying L-type pyruvate kinase/SV40 hybrid genes (Cartier et al. 1992; Miquerol et al. 1996). The intestinal m-ICc12 crypt cells (Bens et al. 1996) were derived from microdissected crypts from the small intestine of 20-day-old fetuses from a pregnant transgenic mouse (L-PK/Tag1) harboring the SV40 large T and small t antigen (Tag) under the control of the 5' regulatory sequences (3200
Electrophysiological Studies
The mpkDCT cells were grown on Transwell filters (Corning Costar; D. Dutscher sa. Brumath, France) in DM until confluence (Day 6) and then in DM without EGF, hormones, and FCS (HFM; the HEPES was replaced by 15 mM NaHCO3) for a further 18 hr. Transepithelial electrical resistance (RT) and voltage (VT) were measured using dual silver/silver chloride (Ag/AgCl) electrodes connected to the Millicell Electrical Resistance System (ERS; Millipore, Bedford, MA). Equivalent Isc was calculated from RT and VT by Ohm's law.
cAMP Assay
cAMP was assayed as previously described (Duong Van Huyen et al. 1998). Confluent mpkDCT cells grown on collagen-coated 12-well plates were incubated with or without dD-arginine vasopressin (dDAVP, 100 M) or parathormone [1-34 synthetic fragment from bovine parathormone (PTH) 10-7 M] for 7 min at 37C. The reaction was stopped by rapidly removing the medium and adding 1 ml ice-cold 95% ethanol/5% formic acid solution. The supernatants were removed and the residues allowed to dry. cAMP was determined using the Pasteur radioimmunoassay kit (no. 79830; Institut Pasteur, Paris, France).
Protein Extraction and Immunoblot Analysis
MRP was detected by Western blotting using MRPr1, an anti-rat IgG2A MAb (Flens et al. 1994,1996). The abdominal cavity of the sacrificed mice was opened. Kidneys and heart were removed and pulverized in liquid nitrogen. The intestine and colon were canulated and rinsed with ice-cold PBS. Small pieces (1 cm long) of duodenum, jejunum, ileum, and colon were removed and placed in ice-cold PBS, centrifuged (150 × g for 5 min at 4C) and kept in liquid nitrogen until use. Confluent cultured cells were scraped off into PBS with a rubber policeman and centrifuged (200 × g for 5 min). The pelleted cells were kept frozen at −80C until use. Tissue samples and pelleted cells were homogenized in a hypertonic solution (0.25 M sucrose, 3 mM imidazole, 1 mM EDTA) containing 0.1 mM PMSF and 100 μ g/ml protease inhibitor cocktail (Boehringer; Mannheim, Germany) in a glass Dounce homogenizer (10 strokes at 4C). All samples were kept at 4C for 30-45 min and centrifuged at 150 × g for 15 min at 4C to remove nuclei and any remaining intact cells. The supernatant was then centrifuged (105,000 × g for 1 hour at 4C). Pelleted membranes from tissues and cultured cells were suspended in 250 μ l lysis buffer and the cytosol from cultured cells was used for Western blotting. Protein content was determined by the Bradford method using bovine serum albumin (BSA) as standard (Bradford 1976). Membrane-enriched or cytosolic fractions were subfractionated by SDS-PAGE using a 6% resolving gel and a 4% stacking gel. Proteins were transferred to polyvinylidene difluoride (PVDF) membrane in 25 mM Tris-HCl, 192 mM glycine, 25% methanol. Rainbow molecular weight markers were used as size standards (Amersham; Arlington Heights, IL). The membrane was blocked by incubation with 5% skim milk in TBS-T (10 mM Tris-HCl, pH 8.5, 150 mM NaCl, 0.1% Tween 20) overnight at 4C. The membrane was probed with MRPr1 (1:500; Kamiya Biochemical, Tukwila, WA) alone, or with MRPr1 plus an anti-β-actin MAb (A-5441; Sigma, St Quentin Fallavier, France) by incubation overnight at 4C and then with the secondary antibodies (horseradish peroxidase-conjucated rabbit anti-rat IgG or alkaline phosphatase-conjugated goat anti-rat or anti-mouse IgG; Dako, Trappes, France) by incubation for 1 hr at room temperature (RT). Positive controls were performed using a membrane-enriched fraction of T47D cells and negative controls were performed by omitting the primary antibody or by replacing MRPr1 by an isotype-matched rat nonimmune IgG2A that did not specifically bind to hematopoietic cells (1:2500; Valbiotech, Paris, France). The membrane was washed with TBS-T and the antigen-antibody complexes were detected with the ECL Plus Western blotting detection system (Amersham Pharmacia Biotech Europe; Orsay, France) or with NBT-BCIP alkaline phosphatase substrate (Sigma).
Immunofluorescence Studies
Mice were anesthetized by
Statistical Analysis
Results are expressed as means ± SE. Means were compared using Student's t-test.
Results
MRP in Normal Mouse Tissues
Western blot analyses to detect MRP were performed on membrane-enriched preparations from normal mouse tissues using MRPr1 (Flens et al. 1994). This anti-rat Mab detected a major 190-kD band, the expected size for MRP (Flens et al. 1996), in mouse kidney, heart, duodenum, jejunum, ileum, and colon (Figure 1). As control, the MRPr1 antibody also recognized a single 190-kD MRP protein band in cultured human breast carcinoma T47D cells expressing MRP mRNA [as do the human breast cancer MCF7 cells (Flens et al. 1994)]. MRP was no longer detected in these cells when MRPr1 was replaced by an isotype-matched rat IgG (Figure 1). As reported for human tissues (Flens et al. 1996), the anti-rat IgG2A MAbs used also crossreacted with a lower molecular weight band (~150 kD) in the jejunum, ileum, and colon using the control isotype-matched rat IgG2A as primary antibody (Figure 1). The colon contained more of the MRP protein than the proximal and distal segments of the small intestine (Figure 1). The kidney also contained a substantial amount of MRP protein, but less than the heart, a tissue that stains strongly for MRPr1 in humans (Flens et al. 1996). The differences in the amount of MRP in the small intestine and colon suggested that the abundance and/or cellular localization of MRP in these tissues differed.
Distribution of MRP in Mouse Small Intestine and Colon
Indirect immunofluorescence studies was used to determine the cellular distribution of MRP in frozen tissue sections of the intestine (jejunum and ileum) and colon. The proliferative crypt compartment of intestinal villi from the jejunum was stained, whereas the cells lining the apex of the villi were not (Figures 2A and 2B). Similar results were obtained with frozen sections of the ileum (not shown). CLSM clearly showed that the six to eight Paneth cells lining the bottom of the intestinal crypts had intensely labeled basolateral membranes (Figure 2C). The population of crypt cells just above the Paneth cells had faint basolateral staining (Figures 2B and 2E). The types of intestinal crypt cells containing MRP were identified by immunofluorescence studies on frozen sections of mouse jejunum using MRPr1 and MIM-1/39 MAb which reacts with the 330- and 350-kD proteins present in the secretory granules of intestinal crypt cells, but not in goblet and Paneth cells (Calvert et al. 1993). In contrast to the gradient of labeling observed in crypt cells with MRPr1 (Figures 2A and 2E), there was intense cytoplasmic staining in the crypt epithelial cells with the MIM-1/39 MAb, which stopped abruptly at the crypt-villous junction (Figure 2F). Therefore, these immunofluorescence studies distinguished two crypt cell populations, one corresponding to Paneth cells and containing MRP only, the other corresponding to partially differentiated enterocytes containing both the MIM-1/39 antigen and weak MRP expression. No labeling was observed in the jejunum of controls in which the primary MRPr1 MAb was replaced by the isotype-matched rat IgG2A (Figure 2D) or by omitting the primary antibody (Figure 2G). Immunofluorescence studies on frozen sections of human ileum also showed that the plasma membrane and, to a lesser extent, the cytoplasm of crypt cells from the terminal ileum were stained for MRPr1 (not shown).
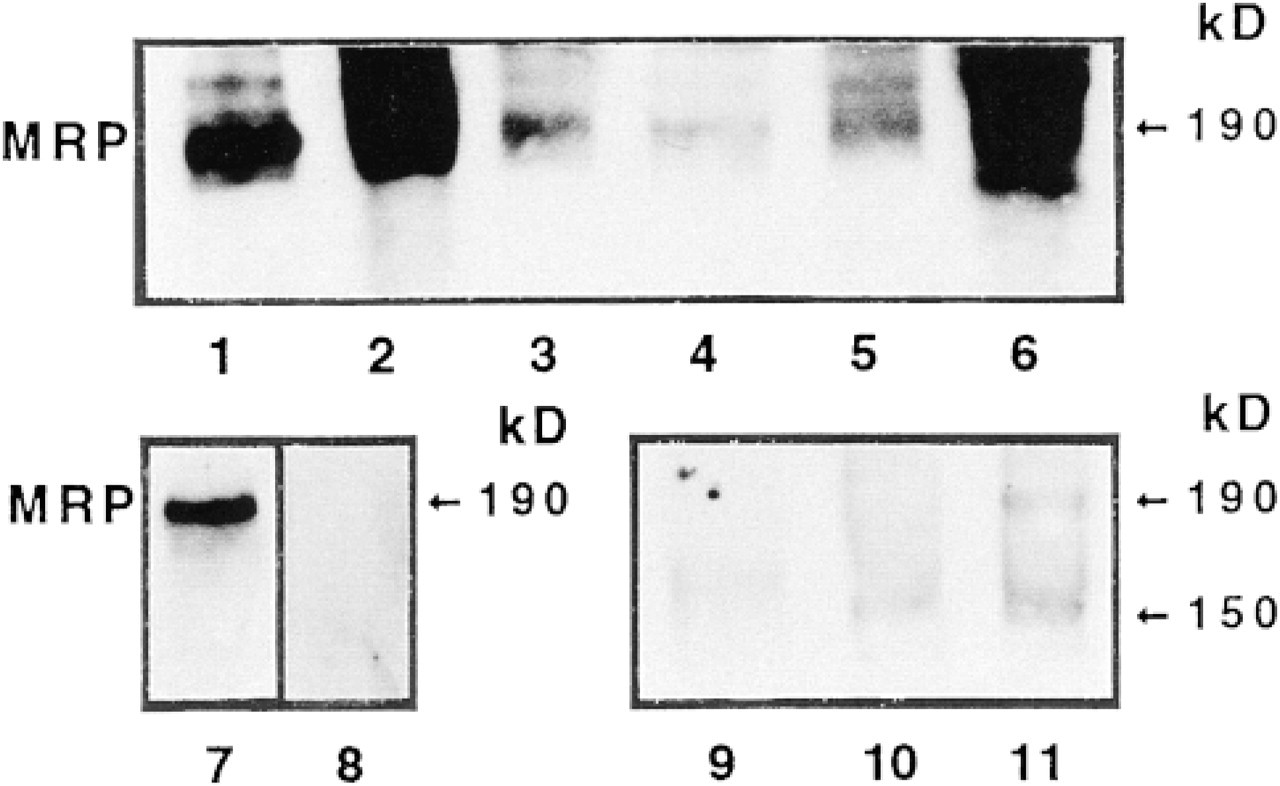
Detection of MRP in normal tissues from adult mice. Western blots with MRPrl were performed on membrane-enriched fractions from kidney (10 μg, Lane 1), heart (5 μg, Lane 2), duodenum (10 μg, Lane 3), jejunum (10 μg, Lane 4), ileum (10 μg, Lane 5), and colon (5 μg, Lane 6). MRPr1 recognizes one major 190-kD band. As controls, the single 190-kD band recognized by MRPr1 in T47D cells (20 μg, Lane 7) was not detected using an isotype-matched rat IgG2A as primary antibody (Lane 8). The MRP 190-kD band was also almost undetectable when MRPr1 was replaced by the isotype-matched rat nonimmune IgG2A in mousejejunum (Lane 9), ileum (Lane 10), and colon (Lane 11). The faint 150-kD band recognized by the rat IG2A isotype is due to crossreactivity.
Colon cells stained positive for MRPr1. In agreement with the abundant MRP detected by Western blotting of the colon membrane-enriched fraction, all colon cells stained intensely all along the entire crypt-villous axis, in contrast to the small intestine (Figures 2H and 2I). CLSM also showed the same predominantly basolateral location of MRP in all colon cells (Figure 2J), as in Paneth cells. Almost no positive staining was observed in the colon using the isotype-matched rat IgG2A as control (Figure 2K) or when the primary antibody was omitted (not shown).
Distribution of MRP in Mouse Kidney
Indirect immunofluorescence studies on frozen mouse kidneys with MRPr1 showed that only some tubule sections were positively stained in the cortex (Figure 3A). Most tubule sections in the medulla were labeled (Figure 3B). In contrast, almost no staining was detected in the papilla (Figure 3C). The controls incubated with the isotype-matched rat IgG2A (Figures 3D–3F) or without the primary antibody (not shown) showed no immunolabeling.
The glomeruli in the kidney cortex were positively stained with MRPr1, and some of the adjacent tubule sections were also stained (Figure 4A). The cellular and subcellular distribution of MRP in the various tubule epithelial cells from the kidney cortex was determined by double indirect immunofluorescence using MRPr1 and antibodies directed against segment-specific antigens. Double labeling with MRPr1 and an antibody against villin, an actin binding protein of the apical brush border of enterocytes and renal proximal tubule cells (Louvard 1989), showed that proximal tubule cells were not stained with MRPr1 (Figures 4B and 4C). Thus, MRP was not detected in proximal tubule cells. Similar studies using MRPr1 and an antibody against TH, present in the thick ascending limb cells (Hoyer et al. 1979), showed that the tubule sections with cytoplasmic MRPr1 labeling were thick ascending limb (TAL) cells (Figures 4D and 4E). Further studies with MRPr1 and an antibody against AQP2, present in the apical membrane of the principal cells (PC) of the cortical collecting duct (CCD) (Nielsen et al. 1993), revealed uneven MRPr1 staining of the membrane in AQP2-positive tubule sections adjacent to the TAL cells (Figures 4F and 4G). CLSM analysis of a CCD double labeled with MRPr1 and the anti-AQP2 antibody clearly showed that MRP was located mostly on the basolateral side of both intercalated cells (IC) and principal cells (PC), which were stained on the apical side with the anti-AQP2 antibody (Figures 5A and 5B). Immunofluorescence studies on frozen sections from the nontumorous part of a human kidney cortex also showed that glomeruli and tubule sections corresponding to distal collecting tubules were stained for MRPr1, as in the mouse (not shown).
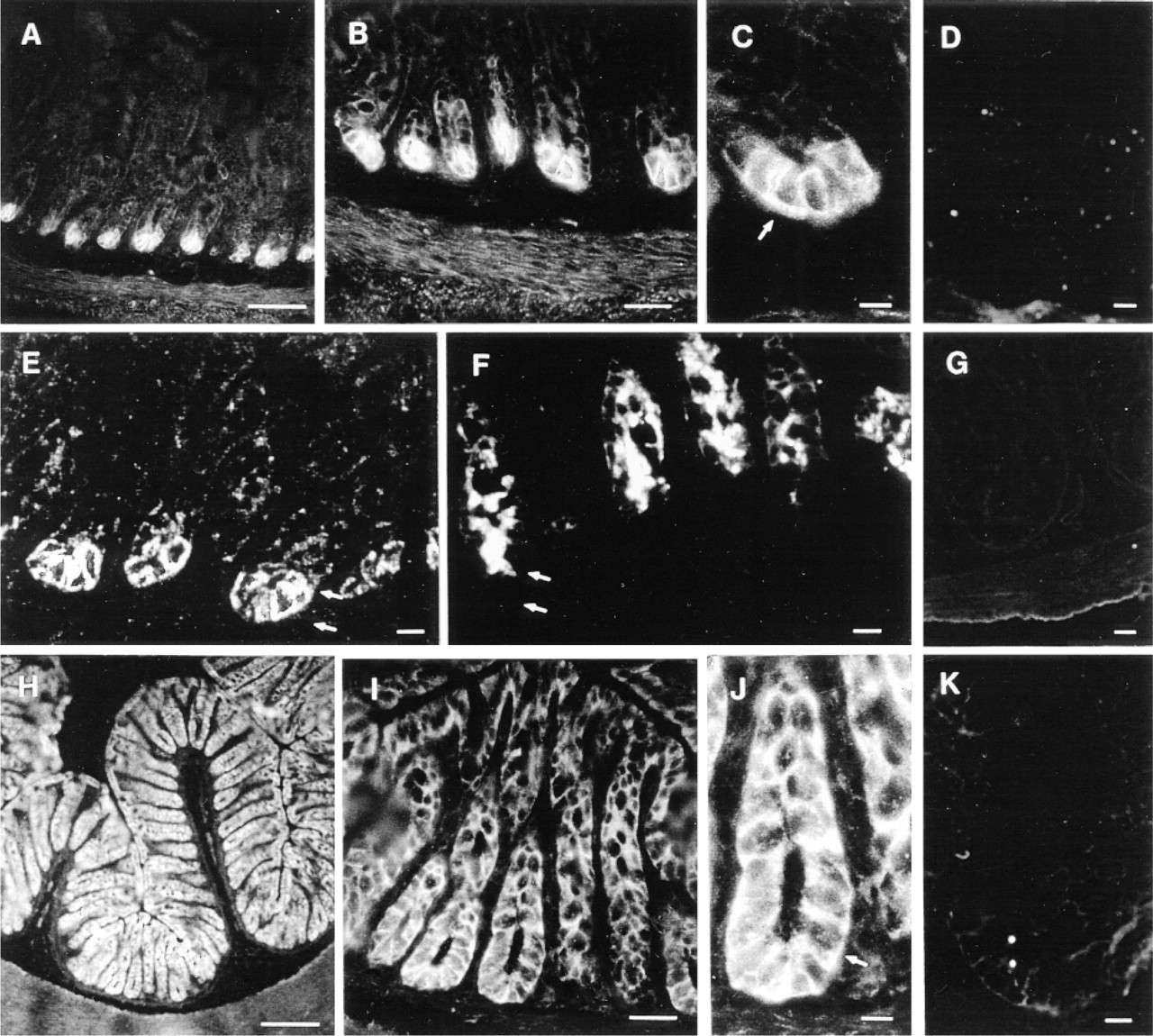
Distribution of MRP in the mouse intestine and colon. Cryosections of mouse jejunum (
Basolateral Location of MRP in Cultured Intestinal Crypt Cells of Mice
We examined established epithelial cell lines not selected by cytotoxic drugs or transfected with MRP cDNA for MRP. We had established a mouse intestinal crypt cell line, m-ICcl2, in which the cells have all the major features of intestinal crypt cells (Bens et al. 1996). These cells were ideal for analyzing the subcellular distribution of MRP in intestinal crypts ex vivo (see Figure 2). The m-ICcl2 cells were grown in Petri dishes and formed monolayers of cuboidal cells and small domes (Figure 6A). Western blotting of membrane-enriched fractions with MRPr1 gave a single 190-kD protein band, the expected size for MRP (Figure 6B, Lane 1). MRP was detected in the membrane-enriched but not in the cytosolic fraction using MRPr1 plus anti-β-actin MAb as internal standard (Figure 6B, Lanes 1 and 2). Indirect immunofluorescence studies analyzed by CLSM (x-y axis) showed that m-ICcl2 cells were strongly stained for MRPr1 around the cell periphery (Figure 6C). Optical sections (x-z plane) showed that MRP was present in the basolateral membranes of the cells (Figure 6D). Positive MRPr1 staining was occasionally seen on juxtaposed (not fully differentiated) cells (see Figure 6G). Most of the apical membrane domain of the cells delimited by tight junctions, as detected by the anti-ZO-1 antibody (Figure 6E) which binds to the tight junction-associated protein ZO-1 (Stevenson et al. 1986), was not labeled with MRPr1. We then tested m-ICcl2 cells for the MIM-1/39 secreted antigen (Calvert et al. 1993). CLSM analysis from optical sections (x-z plane) labeled with the anti-MIM-1/39 MAb alone (Figure 6F) or double labeled with anti-MIM-1/39 MAb and MRPr1 (Figure 6G) showed granular cytoplasmic staining of the antigenic protein within m-ICcl2 cells.
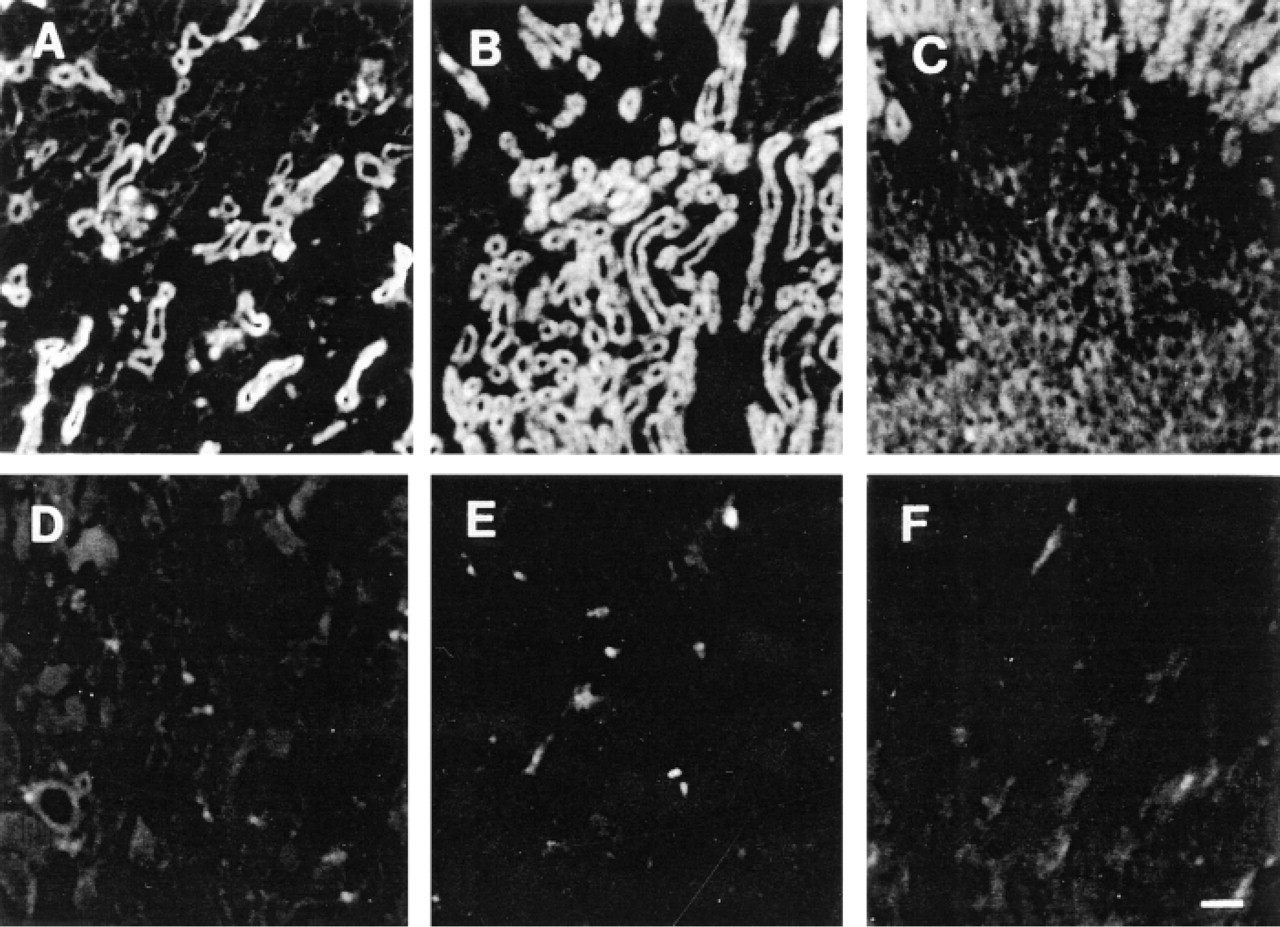
Detection of MRP in the mouse kidney. Indirect immunofluorescence performed with MRPr1 in the renal cortex (
Basolateral Location of MRP in Cultured Renal Distal Tubule Cells of Mice
We then tested cultured distal tubule (DCT) cells for endogenous MRP. We have recently produced new lines of transgenic mice, the SV-PK/Tag mice, containing Tag placed in front of the proximal region (1
Discussion
MRP gene expression and MRP protein have been detected in many tissues and cancer cell lines, showing that MRP has a broad tissue distribution (Lautier et al. 1996; Loe et al. 1996; Flens et al. 1996). Previous immunocytochemical studies have also shown differences in the cytoplasmic and membrane staining patterns in different epithelial cell types, particularly in intestinal tissues, and that MRP is predominantly present in the cytoplasm of cells in normal human tissues (Flens et al. 1996). We now show that the MRP protein may be restricted to particular cell types in the gastrointestinal tract and renal tubule. MRP is present in the cells delimiting the bases of the crypts of normal mouse and human small intestine, mainly in the basolateral membranes of the crypt cells from the small intestine. Paneth cells are the most strongly labeled crypt cells. Another intestinal crypt cell population possessing the MIM-1/39 secreted antigen (Calvert et al. 1993) has weak basolateral MRPrl staining, whereas differentiated enterocytes from the tip of the villi are MRP-negative. Therefore, MRP is present predominantly in the proliferative cell compartment, the site of intestinal epithelial renewal (Cheng and Leblond 1974). In contrast, all the colon cells lining the crypt-villous axis contain MRP. As for intestinal crypt cells, MRP is abundant in the basolateral membranes of these epithelial cells. The same striking heterogeneous cellular distribution of MRP is observed in mouse and human kidneys. Glomeruli and cells from the tubule segments beyond the proximal convoluted tubule contain MRP. However, its subcellular distribution differs from one cell type to another. TAL cells have diffuse intracytoplasmic MRPr1 staining, whereas distal and collecting duct cells have predominantly basolateral MRPr1 staining, with differences in the localization of MRP in the population of intercalated cells. These results are in contrast to the intrarenal distribution of Pgp and MRP2, the hepatocyte canalicular isoform of the multidrug resistance protein encoded by MRP2, also called cMOAT (Büchler et al. 1996; Evers et al. 1998). Both these ABC transporters have been identified in glomeruli (Bello-Reuss and Ernest 1994) and in the brush border of proximal convoluted tubule cells (Thiebaut et al. 1987; Schaub et al. 1997), but not in the distal parts of the renal tubule. Therefore, there is a striking distribution of two distinct ABC transporters (Pgp vs MRP) in the kidney, suggesting that they have distinct physiological functions.
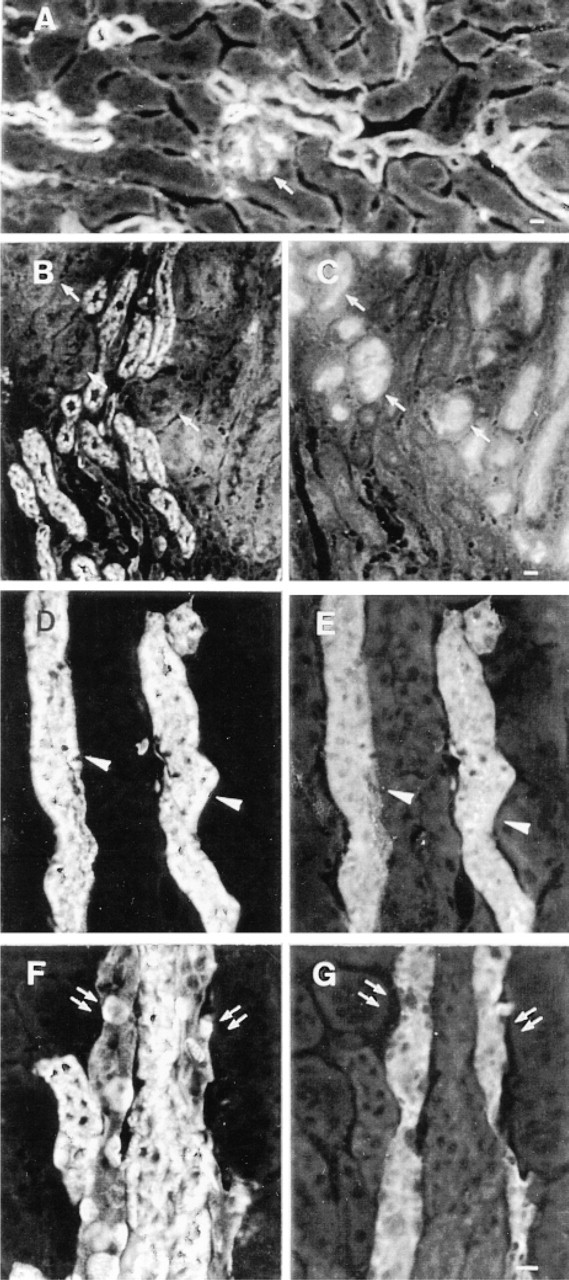
Intrarenal distribution of MRP. Frozen mouse kidney sections were double labeled with MRPr1 and anti-villin, anti-TH, or anti-AQP2 antibodies. One glomerulus (arrow) and some tubule sections close to the glomerulus are stained with MRPr1 (
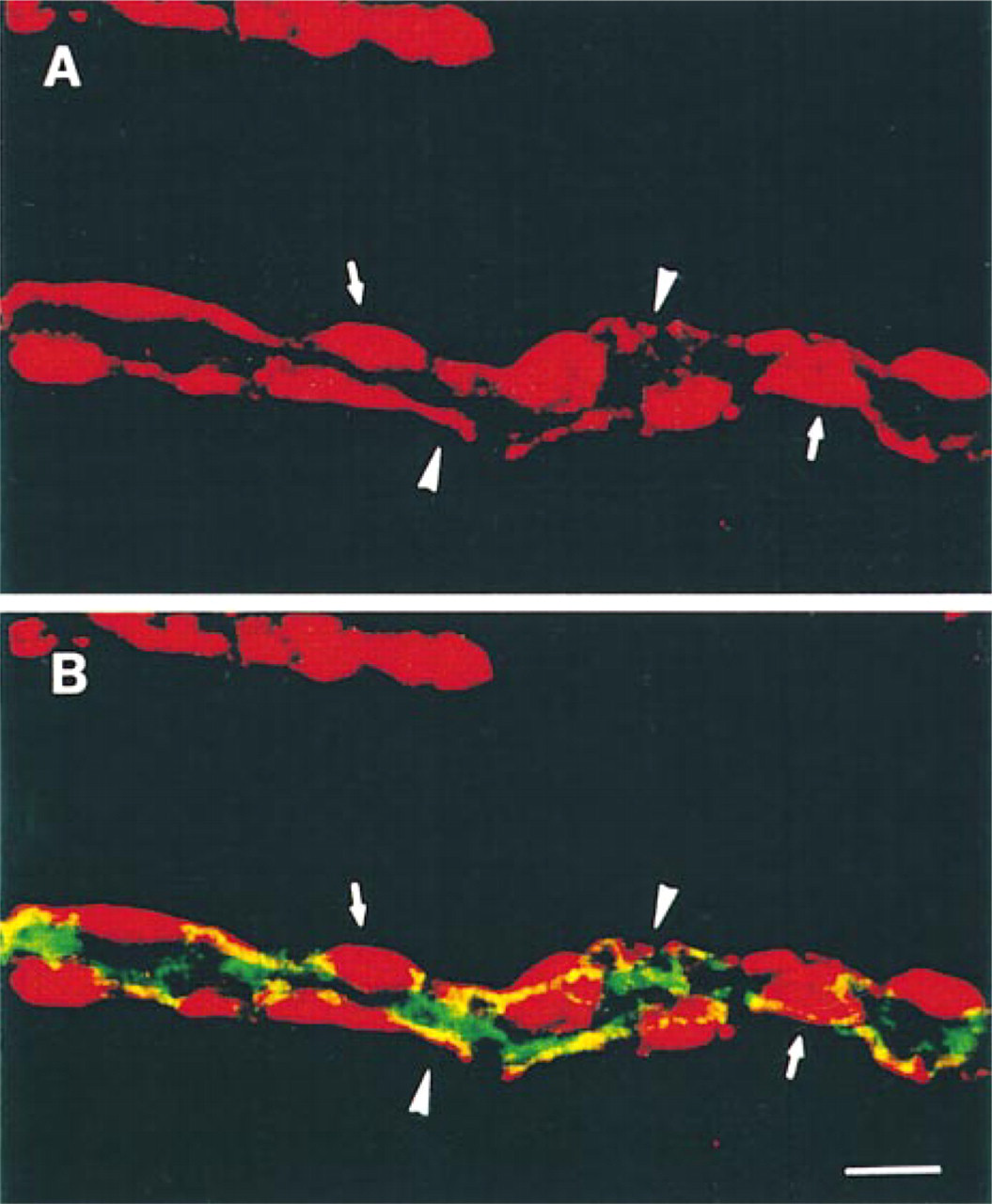
Distribution of MRP in cortical collecting duct cells from the mouse kidney. Double immunofluorescence analyzed by CLSM using MRPr1 (in red) and the anti-AQP2 antibody (in green). Some cells from the collecting duct are intensely labeled with MRPr1 (
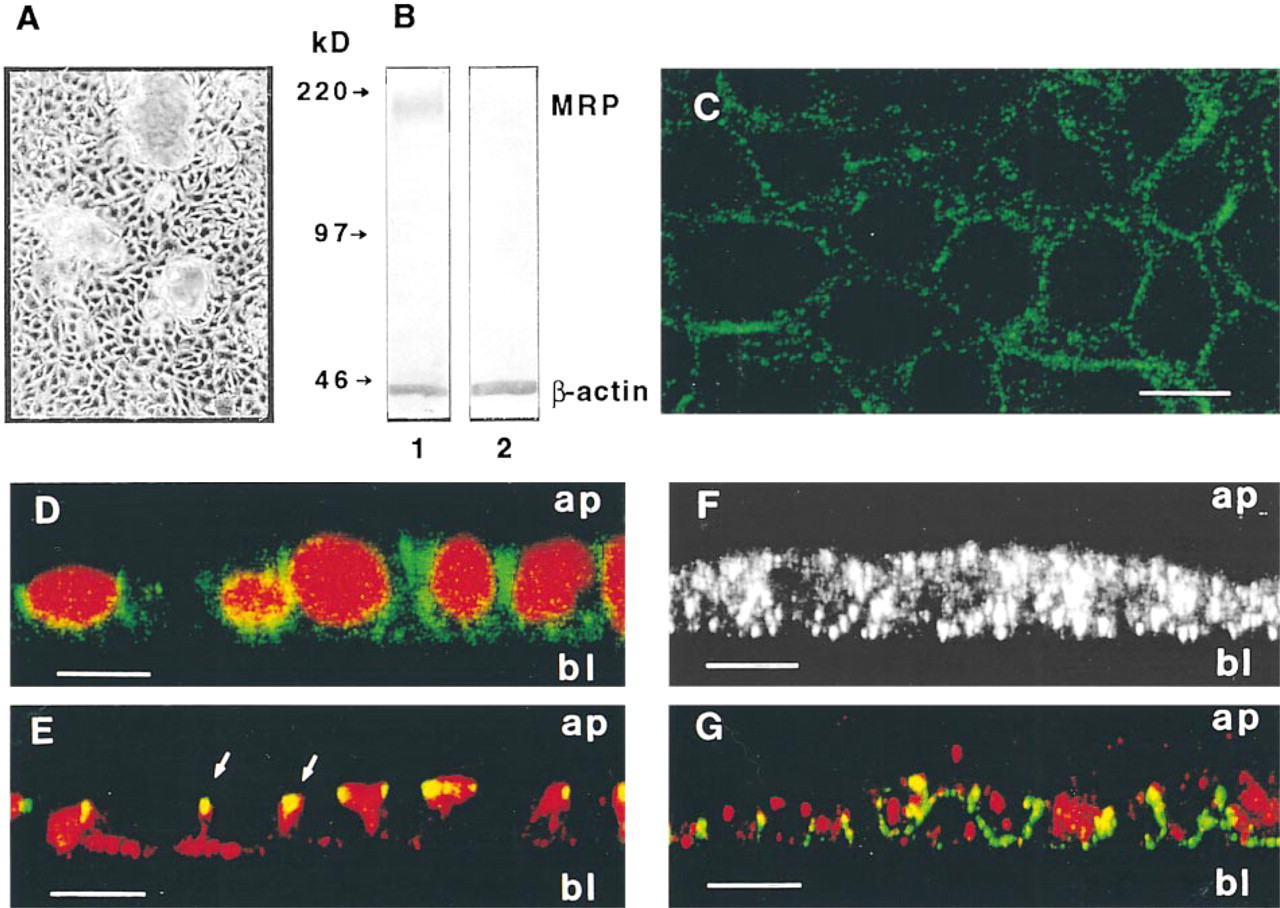
Detection of MRP in cultured intestinal crypt cells from mice. (
MRP is detected in the basolateral membranes of both cultured mouse m-ICcl2 intestinal crypt and distal mpkDCT cells, as well as in the apical membrane of one of the subtypes of DBA-positive intercalated cells. Consistent with the in vivo data, Western blots showed that MRP was more abundant in cultured renal distal tubule cells than in cultured intestinal crypt cells. This confirms the results obtained in intact tissues and provides further evidence that these immortalized cells derived from transgenic mice have retained most of the features typical of their parent cells (Bens et al. 1996; Duong Van Huyen et al. 1998). These particular cultured epithelial cell models are therefore potentially useful tools for further studies on the function of MRP.
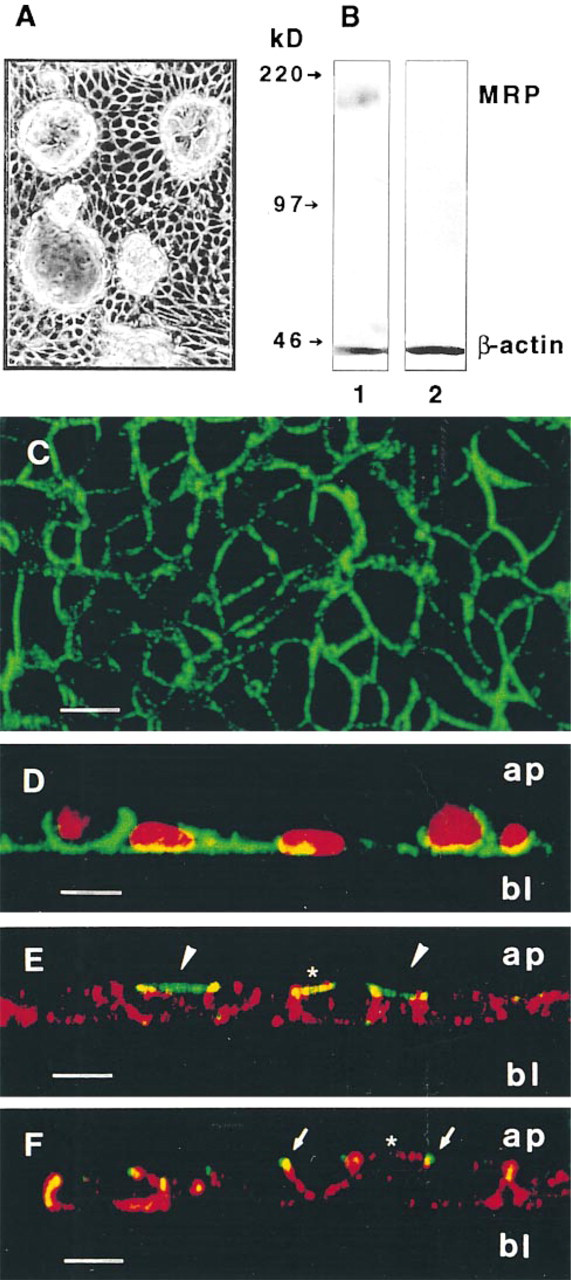
Detection of MRP in cultured distal tubule cells from mice. (
Previous studies have suggested that MRP does not have the same distribution as Pgp in the kidney. Evers et al. (1996) showed that MRP is located in the basolateral membrane domain of pig kidney LLC-PK1 epithelial cells transfected with the human MRP cDNA.
Recently, Brechot et al. (1998) also showed that MRP, unlike the apical location of the cystic fibrosis transmembrane conductance regulator (CFTR) and MDR1 proteins, is restricted to the basal surface of ciliated human bronchial cells and is present in the entire circumference of the plasma membrane from nonpolarized basal cells (lacking tight junctions) of the human bronchial epithelium (Brechot et al. 1998). Flens et al. (1996) first reported the cellular distribution of MRP in human normal and tumor tissues but did not investigate in detail the distribution of the protein in the tissues examined in the present study. They reported that frozen and paraffin sections of normal tissues had mainly a granular MRP cytoplasmic staining, contrasting with the cytoplasmic and plasma membrane MRP staining observed in a variety of human tumor sections. On the basis of these observations, Flens et al. (1996) concluded that MRP could be involved in the transport of compounds into the intracellular compartments of normal cells and acted as a plasma membrane drug efflux pump in tumor cells. It is also possible that the different patterns of MRP staining in normal and tumor cells are due to differences in the amount of MRP, because there is frequently more MRP in tumors and multidrug-resistant cancer cells than in normal tissues or drug-sensitive cells (Flens et al. 1994; Zaman et al. 1994). Evers et al. (1996) have clearly shown that LLC-PK1 cells lacking MRP had MRP protein in the basolateral plasma membrane after being transfected with human MRP cDNA. The fact that we found that MRP was mainly in the plasma membrane from certain specialized intestinal and renal nontumor cell types suggests that these particular epithelial cells normally contain large amount of MRP.
Pgp, MRP, and MRP2, belonging to the superfamily of ABC transporters, are involved in the efflux of cytotoxic drugs (Endicott and Ling 1989; Gottesman and Pastan 1993; Jedlitschky et al. 1994; Lautier et al. 1996), but the particular intestinal and renal distribution of MRP raises questions about its physiological significance. MRP, like Pgp and MRP2, may play a part in the defense of cells against natural toxic compounds (Higgins 1992). Paneth cells and the crypt cell population delimiting the crypt-villous junction are secretary by nature. Therefore, why is MRP in the basolateral membranes and not in apical membranes? To our knowledge, no Pgp has been found in the apical membranes of crypt cells, including Paneth cells. The function of the MIM-1/39 antigen secreted by crypt cells is also unknown, but Paneth cells are involved in the mucosal defense barrier. They apically release granules containing a variety of antimicrobial products, including defensins that are secreted in the intestinal lumen (Ouellette et al. 1989; Ouellette and Selsted 1996). The MRP in the basolateral membranes of Paneth cells and the surrounding crypt cells may produce and transport leukotriene-glutathione conjugates and/or reinforce the role of defensins via basolateral extrusion of natural or exogenous toxic proteins, thus maintaining a favorable environment for the proliferation and differentiation of the main intestinal cell types (absorptive enterocytes and goblet and enteroendocrine cells) (Cheng and Leblond 1974; Potten and Loeffler 1990).
MRP transports cytotoxic drugs and glutathione-conjugated substrates (Leier et al. 1994; Evers et al. 1996). Its presence in the distal parts of the renal tubule raises the question of whether this membrane transporter has other physiological functions. It has been suggested that MRP is involved in regulating ion transporters and channels. ABC transporters are believed to regulate the activity of CFTR and volume-regulated Cl− channels (Loe et al. 1996). A chloride conductance regulator (EBRC) thought to be a AMP-activated Cl− channel, belonging to the ATP binding protein family, is present in the basolateral membranes of TAL, DCT, and CCD cells of rabbit kidney (van Kuijck et al. 1996). Therefore, the presence of MRP in these kidney cells indicates that several ABC transporters can be expressed within the cells of the late segments of the renal tubule, but their functions remain to be determined.
The distal segments of the renal tubule are also the main sites of action of corticosteroid hormones (Rossier and Palmer 1992). Cultured human embryonic kidney cells transfected with the MRP or the mrp (the murine homologue of MRP) cDNA, actively transport leukotriene C4, 17 β-estradiol, and 17β
Footnotes
Acknowledgments
Supported by INSERM and by grants from the Association pour la Recherche sur le Cancer and the European Community (BIO4-CTT 960052). K-C Peng holds an INSERM postdoctoral (Poste vert) fellowship supported by the Conseil Régional d'Ile de France.
We thank F. Paraf for providing the human tissues. We thank R. Calvert, M. Knepper, S. Robine, and D. Louvard for generously providing us with valuable antibodies, and Y. Pilatte (INSERM U129, Créteil, France) for advice on performing Western blotting. We thank S. Roger and P. Disdier for photographic work and Ms J. Knight and Dr O. Parkes for editing assistance.