
Abstract
We used three anti-H monoclonal antibodies (MAbs) specific for H Type 1, H Type 2, and H Type 3/4 antigens to investigate the distribution of H Type 1–H Type 4 chains of the ABO(H) histo-blood group in the human respiratory system. Strong staining of H Type 1 chain and weak staining of H Type 2 chain were observed in mucous cells of submucosal glands of bronchial epithelium, which were dependent on the secretor status. No H Type 3/4 chains were detected in mucous cells. Serous cells of submucosal glands of respiratory system showed no staining by three anti-H antibodies. H Type 1 and H Type 3/4 antigens were detected heterogeneously in apical surfaces of bronchial epithelium from secretors but not from nonsecretors. In contrast, basal cells of bronchial epithelium expressed H Type 2 irrespective of the secretor status, probably regulated by the H gene. Some alveolar Type II cells contained only H Types 3/4, which were dependent on the secretor status, whereas alveolar Type I cells had no H antigens. Our results indicated that different cell types in respiratory epithelium expressed different types of carbohydrate chains of histo-blood group antigens under the control of the H or the Se gene.
A
The epitope of the H antigen, the precursor of A, B, LeY, and Leb antigens of ABO and Lewis histo-blood groups, is carried by at least four different types of internal carbohydrate backbones (Type 1, Galβ1–3Glc-NAcβ1-R; Type 2, Galβ1–4GlcNAcβ1-R; Type 3, Galβ1–3GalNAcα1-R; and Type 4, Galβ1–3GalNAc-β1-R) (Clausen and Hakomori 1989). However, much less is known about the distribution and the amount of each type of chain of ABH antigens in tissues. We have immunohistochemically determined the differences in the distribution of H Type 1, H Type 2, and H Type 3/4 antigens in human submandibular gland, using specific monoclonal antibodies (MAbs) (Liu et al. 1998, 1999). The distribution of ABO and Lewis antigens in many normal or tumor tissues has been investigated by lectin or immunohistochemical methods (Mazzuca et al. 1982; Yuan et al. 1985; Cossu et al. 1990; Ito et al. 1990; Maritgnone et al. 1993; Liu et al. 1998, 1999). However, the distribution of Type 1–Type 4 chains of blood group antigens in human respiratory epithelium is not yet reported (Alvarez-Fernandez, and Carretero–Albiñana 1991; Bals and Welsch 1997).
The aim of this study was to investigate the cell type distribution of H Type 1, H Type 2, and H Type 3/4 substances in the human respiratory system by an immunohistochemical method using three anti-H MAbs specific for H Type 1 chain (1E3), H Type 2 (3A5), and H Type 3/4 chains (MBr1). These data are useful for a better understanding of the biology of secretory cell types and of several pathological conditions that are characterized by altered secretion.
Materials and Methods
Twenty-two human lung tissue specimens were obtained from autopsies in our department. The ABO and Lewis histo-blood groups of tissue donors were assessed by routine blood agglutination methods. The secretor status was determined according to the Lewis phenotypes. Individuals with Le(a + b –) or Le(a + b +) phenotype were classified as nonsecretors, while those with Le(a–b +) phenotype were classified as secretors. This study included six samples with blood group A, six with blood group B, and 10 with blood group O; 12 of these individuals were secretors and 10 were nonsecretors.
Deparaffinized serial sections (3 μm) of 10% formalin-fixed, paraffin-embedded human lungs were pretreated with 3% hydrogen peroxide for 30 min to quench endogeneous peroxidase activity. Then the pretreated sections were immunostained using a streptavidin–biotin complex immunoperoxidase method as described previously (Liu et al. 1998, 1999). Controls without the first antibody showed negative staining.
Commercial anti-A or anti-B can not discriminate Type 1–Type 4 carbohydrate chains. Therefore, a panel of specific anti-H MAbs was used in this study. MAb 1E3, which was raised after immunization of mixed saliva of blood group O individuals, was a kind gift from Drs. Ken Furukawa and Shin Yazawa (Department of Legal Medicine, Gunma University School of Medicine). Although anti-H 1E3 has been demonstrated to be reactive with H Type 1–H Type 4 synthetic trisaccharides (Nakajima et al. 1993), we demonstrated that anti-H MAb 1E3 is reactive predominantly with H Type 1 and H Type 2 antigens in situ in human submandibular glands (Liu et al. 1998; Liu et al. 1999). In addition, we demonstrated that anti-H 1E3 became specific for H Type 1 (Fucα1–2Galβ1–3GlcNAcβ1-) after absorption with red cell ghosts (Liu et al. 1998). We used red cell ghost-absorbed MAb 1E3 throughout all experiments in this study. The anti-H MAb 3A5, raised against membranes of red blood cells from blood group O, has been demonstrated to be specific for H Type 2 structure (Fucα1–2Galβ1–4GlcNAcβ1-) (Liu et al. 1996). The anti-H MBr1, specific for H Type 3/4 structures (Galβ1–3GalNAc-) (Menard et al. 1983; Bremer et al. 1984; Adobati et al. 1997), was a kind gift from Dr. Maria I. Colnaghi (Division of Experimental Oncology E; National Institute for Cancer Research, Milan, Italy). The MBr1 was raised against cell membranes of the breast cancer cell line MCF7. Anti-H MBr1 is unable to discriminate H Type 3 (Fucα1–2Galβ1–3GalNAcα-) from H Type 4 (Fucα1–2Galβ1–3GalNAcβ-). Ulex europaeus agglutinin I (UEA-I) conjugated with horseradish peroxidase (EY Laboratories; San Mateo, CA) was used to compare with staining by anti-H antibodies. Anti-factor VIII (Dako; Carpinteria, CA) and anti-SP-A (Dako) were also used to identify alveolar Type I and Type II cells, respectively.
Results
In this study, we used specimens mainly from blood group O individuals to avoid an effect of metabolic conversion of H carbohydrate chains to A and B chains.
Figure 1 shows staining of submucosal glands of bronchial epithelium from O secretors by anti-H MAbs 1E3, 3A5, and MBr1. MAb 3A5, specific for H Type 2 antigen, strongly stained capillary endothelial cells and erythrocytes (Figure 1B), whereas no staining was observed with erythrocyte-absorbed anti-H 1E3 (Figure 1A), indicating that MAb 1E3 was specific for H Type 1 chain under certain conditions. In secretors, the cytoplasm of mucous cells of submucosal glands of bronchial epithelium was strongly stained with 1E3 (Figure 1A) and UEA-I (Figure 1D), or by anti-A and anti-B (not shown), and was weakly stained with 3A5 (Figure 1B), but not with MBr1 (Figure 1C). In nonsecretors, no staining of mucous cells with any of the anti-H MAbs and with UEA-I was observed (not shown). Serous cells did not react with any of the anti-H MAbs, in contrast to serous cells of secretory acini of the submandibular gland, which were strongly stained by MAb MBr1 (Liu et al. 1999).
The bronchial epithelial cells showed heterogeneous staining with the anti-H MAbs according to cell type. Luminal surfaces of bronchial epithelium were labeled heterogeneously by anti-H 1E3 and MBr1, depending on the secretor status (Figure 2A and 2C). However, the cytoplasm of columnar cells and cuboidal cells of bronchial epithelium completely lacked positive staining by 1E3 and MBr1. Anti-H MAbs did not stain a majority of goblet cells, but a few goblet cells from secretors reacted with 1E3 and MBr1. The contents of bronchioles were strongly stained with 1E3 and MBr1 and weakly with 3A5 (Figure 2B and 3B), suggesting that carbohydrate chains of mucins consisted of various type of H-active chains. Cytoplasm of basal cells from both secretors and nonsecretors was labeled with 3A5 (Figure 2B and 2D) or with anti-A or anti-B (not shown), corresponding to the ABO blood groups. However, UEA-I did not stain basal cells (not shown), suggesting that the substances positive for anti-H 3A5 and for UEA-I were different, as reported previously (Liu et al. 1998).
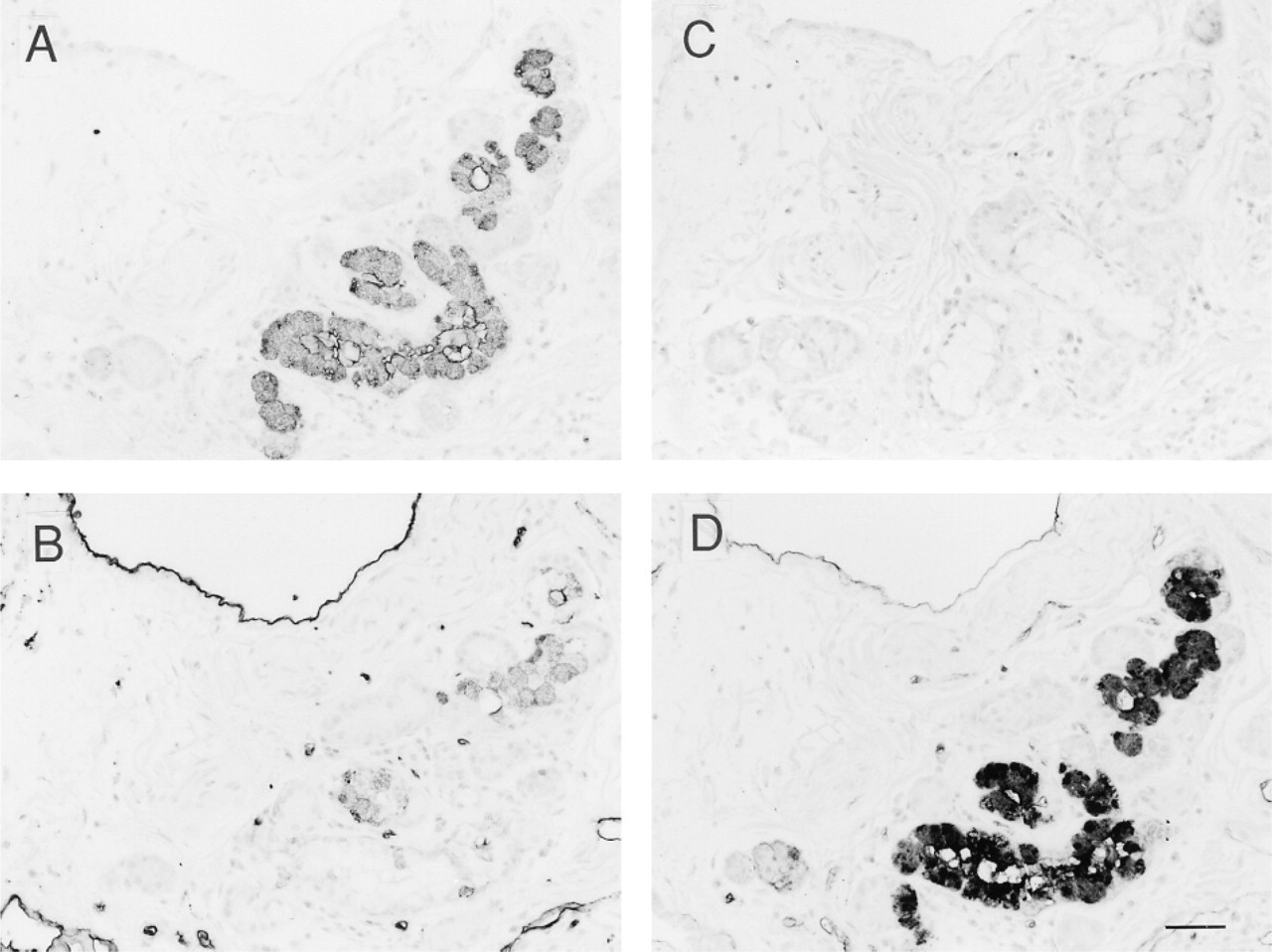
Immunostaining of serial sections of submucosal glands of bronchial epithelium from a blood group O secretor with monoclonal anti-H antibodies, 1E3, 3A5, and MBr1, and lectin UEA-I. Mucous cells from secretors were stained strongly by 1E3 (
Type I cells (squamous alveolar cells) of alveoli from both secretors and nonsecretors did not react with any of the anti-H antibodies (not shown). However, about 15% of alveolar Type II cells (great alveolar cells) reacted with MBr1 only from secretors but not from nonsecretors (Figure 4). The MBr1-positive cells were stained with anti-SP-A (not shown). All results are summarized in Table 1.
Discussion
Although many studies have investigated the distribution of ABO and Lewis antigens in normal and malignant tissues of the digestive system, studies on the distribution of these antigens in the respiratory system have been few. Hirohashi et al. (1984) and Alvarez–Fernandez and Carretero–Albiñana (1991) demonstrated the presence of H-like antigen in bronchial mucous cells and bronchial goblet cells using UEA-I lectin, which was previously believed to be specific for the H Type 2 chain. Recently, Bals and Welsch (1997) reported similar findings using commercial anti-H (probably H Type 2-specific) and UEA-I. However, we demonstrated that the specificity of UEA-I was broader than previously imagined (Mazzuca et al. 1982), because we found a significant difference in reactivity between MAb anti-H 3A5 and UEA-I in basal cells of bronchial epithelium and in human submandibular gland, as reported previously (Liu et al. 1998). Accordingly, there was no study to focus the distribution of the Type 1–Type 4 carbohydrate chains in the respiratory system. The only study focused on Type 2 carbohydrate chain was that of Zenita et al. (1988), in which they found the expression of LeY (Type 2) antigen in lung cancer tissues from all patients examined. In the present study, we examined the distribution of H Type 1–4 carbohydrate chains of the ABO(H) system in the respiratory system using anti-H MAbs specific for H Type 1, H Type 2, and H Type 3/4. We demonstrated the expression of H Type 1 and H Type 3/4 antigens in apical surfaces of bronchial epithelium and of H Type 1 and H Type 2 in mucous cells of bronchial submucosal glands from secretors but not from nonsecretors. Although we demonstrated previously the presence of H Type 3/4 in serous cells of secretory acini of the submandibular gland (Liu et al. 1999), serous cells of bronchial submucosal glands showed no staining by anti-H MBr1. These results suggest that the embryogenetic mechanism of evolution of serous cells of the airways is different from that of the submandibular gland and pancreas (Ito et al. 1990). Basal cells of bronchial epithelium expressed the H Type 2 antigen independent of the secretor status (Figure 2), in contrast to the results of Bals and Welsch (1997), in which they found positive staining of basal cells from only secretors by commercial anti-H MAb and UEA-I. We found only a few goblet cells expressing H Type 1 and H Type 3/4 chains. Mazzuca et al. (1982) suggested differences in the glycosylation abilities of mucous glands and goblet cells, using various lectins.
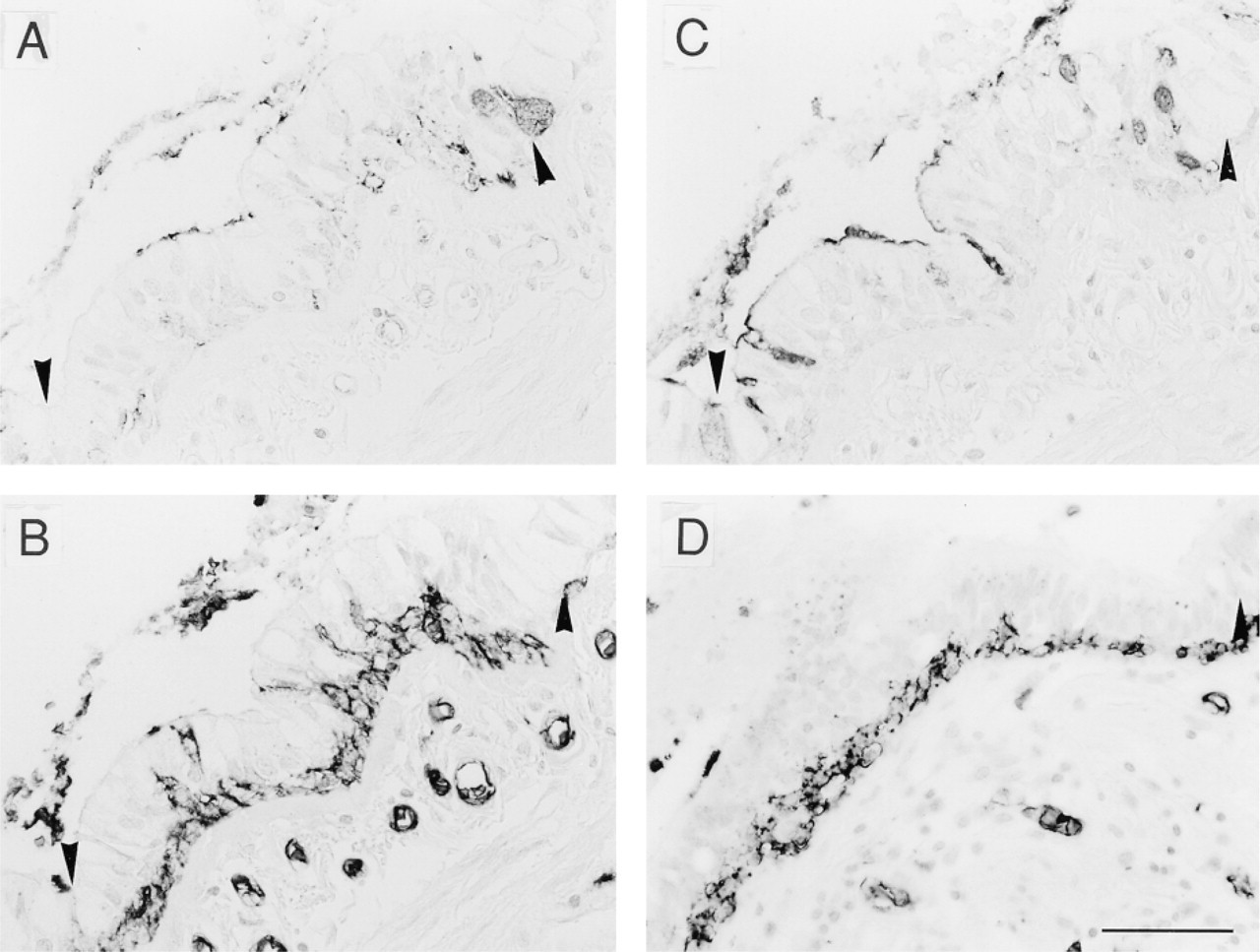
Immunostaining of serial sections of bronchial epithelium from blood group O with anti-H MAbs 1E3, 3A5, and MBr1. Luminal surfaces of bronchial epithelium from secretors were labeled by anti-H 1E3 (
Although the ABO histo-blood group was identified 100 years ago and has contributed enormously to transfusion medicine, its function is still an enigma. However, there is increasing evidence that the remarkable diversity of carbohydrate chains of respiratory mucins allows many interactions with microorganisms and may be an important factor in maintaining the sterility of the respiratory tree (Ramphal et al. 1989, 1991; Lambin et al. 1991). Many complexed carbohydrates were also found in human milk and were suggested to play a role as antipathological agents for breast-feeding infants (Newburg 1999). Carbohydrate chains of human respiratory mucins are responsible for 70–80% of the weight of glycoproteins, and major sugars of mucins consist of fucose, galactose, N-acetylglucosamine, N-acetylgalactosamine, and N-acetylneuraminic acid, with a small amount of mannose (Lambin et al. 1991; Sangadala et al. 1992), suggesting that most of carbohydrate chains are O-linked carbohydrate chains. Accordingly, the composition of sugars is markedly changed by genetic variation of glycosyltransferases. A Lewis-negative nonsecretor individual with blood group O may have a reduced amount of fucose, galactose, and N-acetylgalactosamine. Here, we described the distribution of well-characterized Type 1–4 carbohydrate chains of the ABO system as one of the markedly diverse carbohydrate chains in the respiratory system. In addition to genetic background, pathological conditions also alter the composition of glycoconjugates of mucins. A remarkable diversity of carbohydrate chains has been observed in mucins from patients with cystic fibrosis (Roussel et al. 1975; Ramphal et al. 1989), chronic bronchitis (Feldhoff et al. 1979; Lhermitte et al. 1991), or bronchiectasis (Van Halbeek et al. 1988). Although the association of chronic obstructive pulmonary disease with the α1-anti-trypsin gene has been established, the associations of pulmonary diseases and lung function with other genetic risk factors, including glycosyltransferases for synthesis of histo-blood group antigens, were also suggested (Springer 1970; Raza et al. 1991; Ramphal et al. 1991; Kauffmann et al. 1996; Scharfman et al. 1996; Sandford et al. 1997; Weimann et al. 1998). However, the associations of blood group antigens with chronic obstructive pulmonary disease are controversial. There are some studies that did not detect these associations and some that showed contradictory results (Abboud et al. 1982; Sandford et al. 1997).
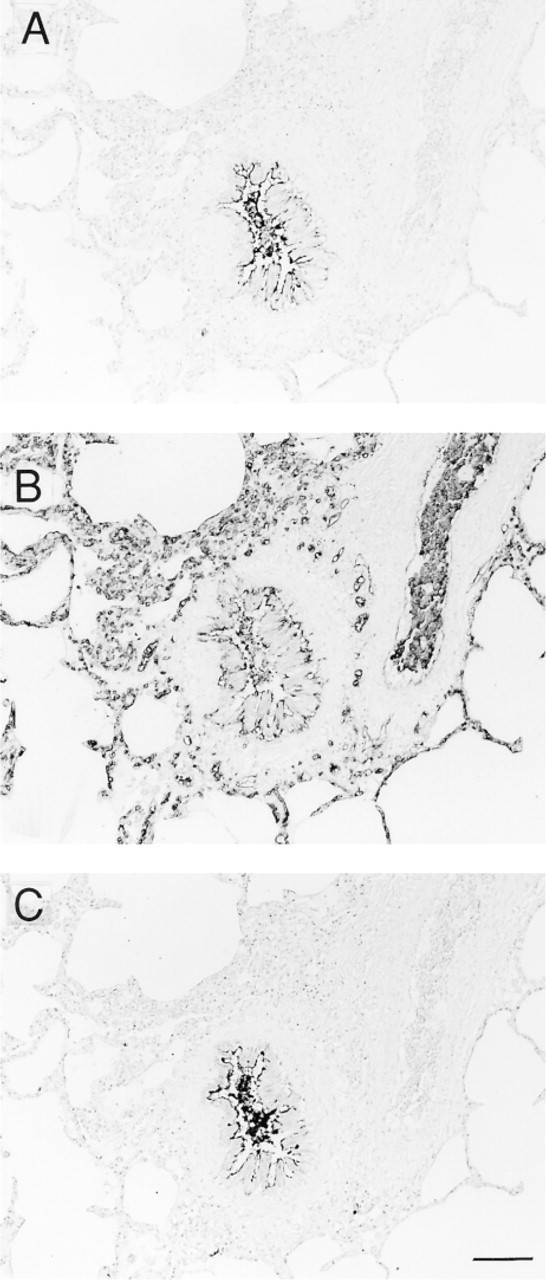
Immunostaining of serial sections of bronchioles from a blood group O secretor with anti-H MAbs. Contents of bronchioles were strongly stained with 1E3 (
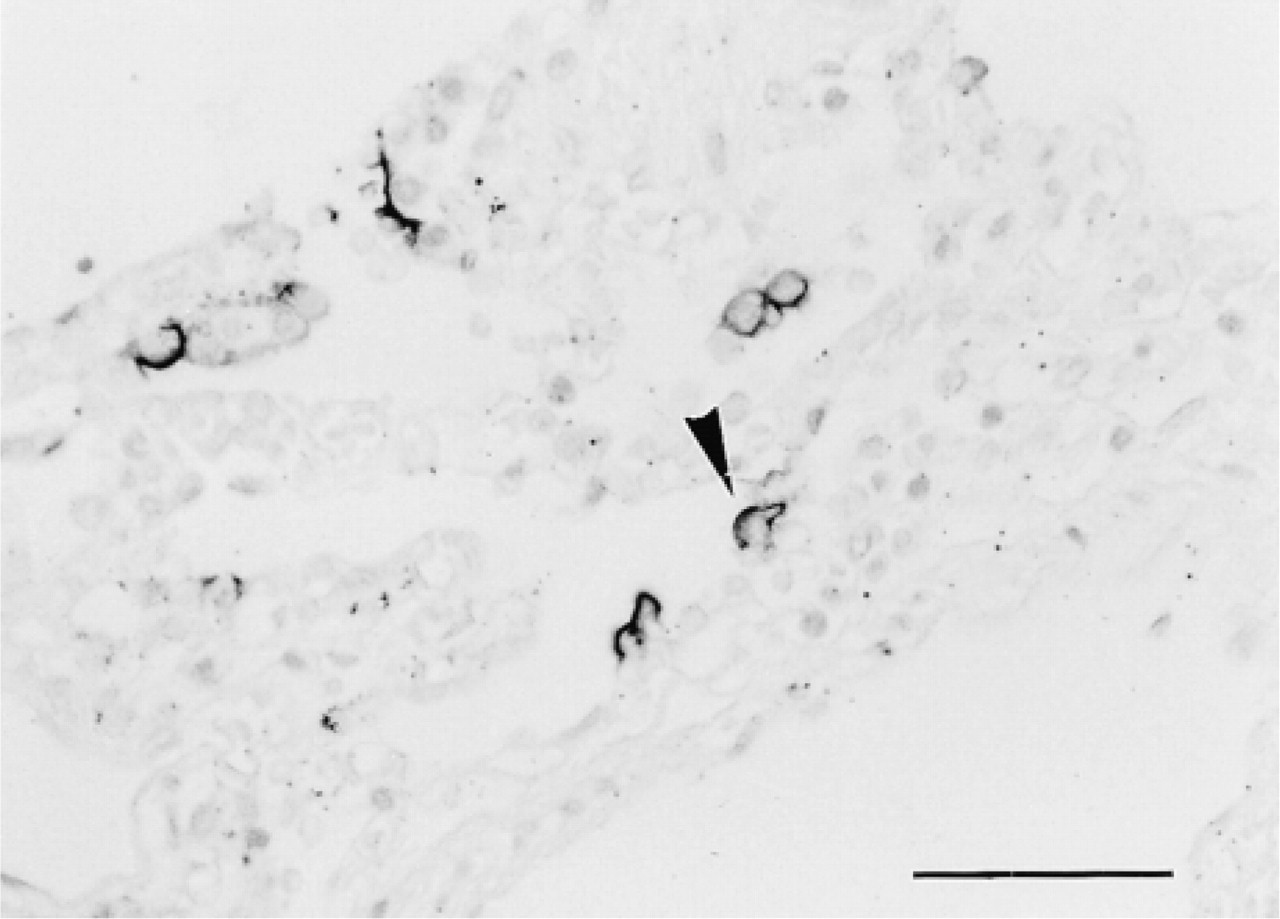
Immunostaining of a section of alveoli from an O secretor with anti-H MAb MBr1. Type II cells (great alveolar cells) were stained with MBr1 but not with other anti-H antibodies. Bar = 50 μm.
It is known that many carbohydrate structures are recognized by microorganisms (Plotkowski et al. 1993). Accordingly, the remarkable diversity of mucin carbohydrate chains would provide many ways for the interactions of sugars and microorganisms as part of the defense mechanisms of the respiratory system. Several potential carbohydrate sites have been identified for microorganisms. Ramphal et al. (1991) have studied binding of P. aeruginosa to a Gal-GlcNAc moiety that is a major component of glycoconjugates of mucins, and have found that the Type 1 (Galβ1–3GlcNAc) and Type 2 (Galβ1–4GlcNAc) disaccharide units were receptors for P. aeruginosa. There are also many reports regarding the secretor phenotype and infections. Some reports describe the disadvantage of the nonsecretor phenotype in bacterial infections (Blackwell et al. 1986a, b), but Raza et al. (1991) reported the advantage of nonsecretors for respiratory viral infections. Kauffmann et al. (1996) also found the disadvantage of Lewis-negative nonsecretors for lung function and asthma after epidemiological examination of a cohort of 228 coal miners.
The immunohistochemical distribution of ABH antigens in human lung
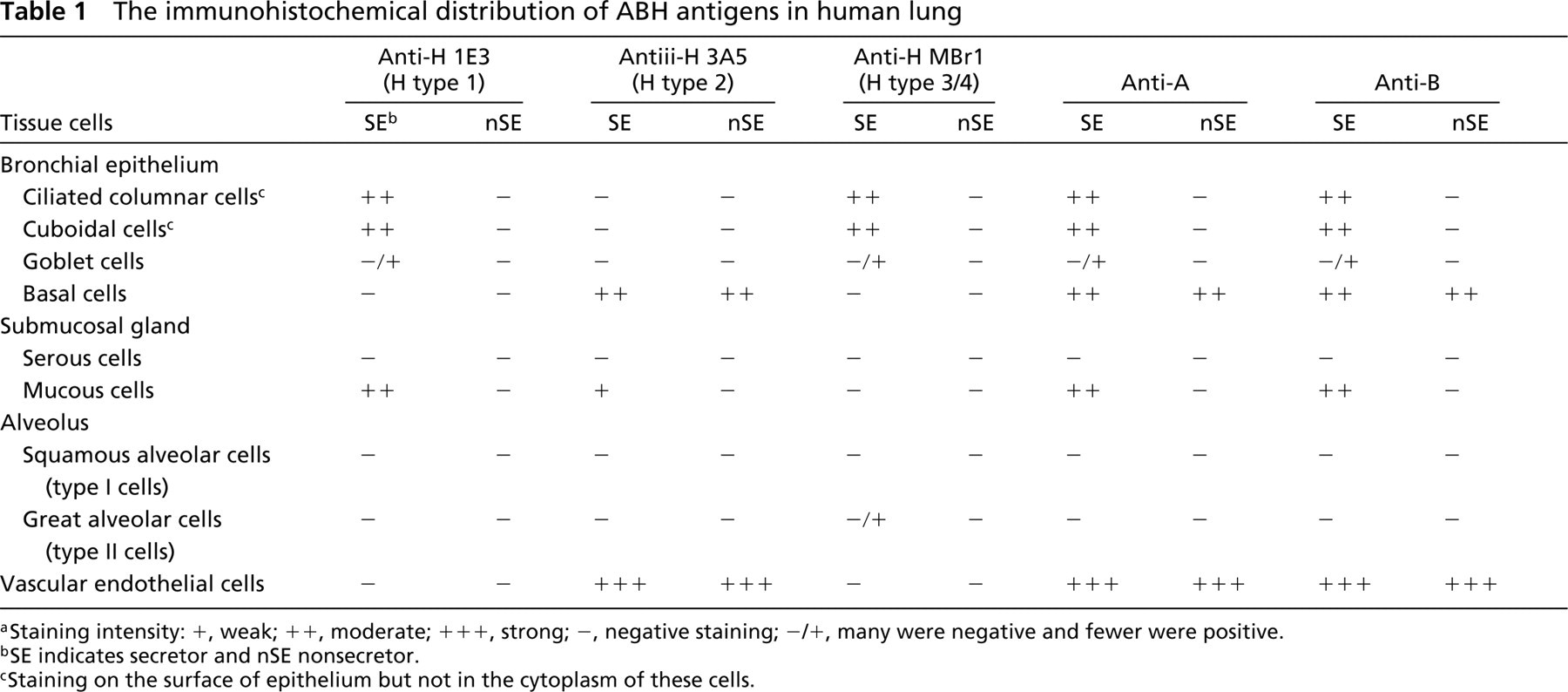
aStaining intensity: +, weak; + +, moderate; + + +, strong; –, negative staining; –/+, many were negative and fewer were positive.
bSE indicates secretor and nSE nonsecretor.
cStaining on the surface of epithelium but not in the cytoplasm of these cells.
It is plausible that carbohydrate chains of mucins play an important role in binding of pathogens in the respiratory organs. However, further studies are required to clarify the relation between carbohydrate structures and binding of each pathogen or in pulmonary diseases.
Footnotes
Acknowledgements
Supported in part by a grant-in-aid for Scientific Research from the Ministry of Education, Science, Culture and Sports of Japan.
We thank Drs Ken Furukawa and Shin Yazawa (Department of Legal Medicine, Gunma University School of Medicine) and Dr Maria Ines Colnaghi (Division of Experimental Oncology E; National Institute for Tumor, Milan) for the kind gifts of the anti-H 1E3 and the anti-H MBr1 monoclonal antibodies, respectively. We also thank Mr Shigeo Kamimura and Ms Yasuko Noguchi for technical assistance.