
Abstract
Studies on the effects of pulmonary toxicants on the lung often overlook the fact that site-specific changes are likely to occur in response to chemical exposure. These changes can be highly focal and may be undetected by methods that do not examine specific lung regions. This problem is especially acute for studies of the conducting airways. In this study, differential gene expression of secreted proteins in the lung by different methods of collection (whole lung, gross airway microdissection, and laser capture microdissection) and by airway levels (whole lobe, whole airway tree, proximal airways, airway bifurcations, and terminal bronchioles) was examined. Site-specific sampling approaches were combined with methods to detect both gene and corresponding protein expression in different lung regions. Differential expression of mRNA by both airway level and lung region was determined for Clara cell secretory protein, calcitonin gene-related peptide, uteroglobin-related protein 2, surfactant protein A, and surfactant protein C. Therefore, for maximal enrichment of mRNA and maximal ability to identify changes in mRNA levels in the diseased state or in response to chemical exposure, it is critical to choose the appropriate airway region and sample collection method to enrich detection of the transcript(s) of interest. (J Histochem Cytochem 58:1107–1119, 2010)
Keywords
T
The majority of lung toxicity studies investigate global, lung gene expression changes in response to chemical exposure; the whole lung is homogenized without regard for microenvironmental regions and their contribution to the cellular response. Although this approach can yield valuable data, many pulmonary toxicants have site-specific effects that can be diluted or undetectable using this approach. The purpose of this study is to evaluate gene expression of a select group of genes for secreted proteins involved in chronic airway diseases by both conventional whole lung lobe analysis and by newer, more site-selective approaches [gross airway microdissection and laser capture microdissection (LCM)] to compare and quantify differential gene expression. The site of secreted protein synthesis is especially important to determine because these proteins can move far from their site of origin.
Secreted proteins in the lung are abundant, but distribution varies for both sites of synthesis and sites of action. For this study, members of the secretoglobin family [SCGB1A1, also known as Clara cell secretory protein (CCSP), and SCGB3A1, also known as uteroglobin-related protein 2 (UGRP2)], components of lung surfactant [surfactant protein A (SP-A) and surfactant protein C (SP-C)], and neuropeptide calcitonin gene-related peptide (CGRP) were investigated because of their importance in normal lung function and homeostasis. Furthermore, many of these genes have either gene/protein alterations or deficiencies in lung disease. For example, CCSP, a member of the secretoglobin family also known as CC10, is a 10-kDa protein with anti-inflammatory properties, which is regarded as a Clara cell marker in laboratory animals because mature Clara cells express abundant levels of this protein and its pattern of expression parallels postnatal Clara cell maturation (Singh et al. 1990; Cardoso et al. 1993; Singh and Katyal 1997; Coppens et al. 2009). Moreover, CCSP has clinical implications in human airway disease–-it has been shown that smokers, asthmatics, and patients with lung cancer have significantly decreased CCSP expression in serum and lavage fluid (Shijubo et al. 2003). Hence, site-specific investigation of changes in CCSP expression in response to toxicant exposures may give further insight into the role of CCSP in the diseased lung.
CGRP is a neuropeptide secreted from pulmonary neuroendocrine cells (PNECs) present in the airway epithelium. CGRP is substantially enriched in airway bifurcations and in neuroepithelial bodies (NEBs) along the main axial path of the conducting airways (Peake et al. 2000; Reynolds et al. 2000). NEBs have CGRP-expressing PNECs, and co-localized injury-resistant CCSP-expressing cells are involved in epithelial regeneration and repair after epithelial injury (Reynolds et al. 2000; Hong et al. 2001). Because of the specific localization of CGRP and its role in cell proliferation and PNEC hyperplasia (Stevens et al. 1997), pulmonary toxicants affecting bifurcations and terminal bronchioles could have detrimental consequences on epithelial repair. Detection of site-specific effects may be overlooked by conventional methods of sample collection for gene expression studies as conducting airways comprise a small percentage of the whole lung volume and CGRP is highly expressed only in a subset of these conducting airways.
UGRP2 is a member of the secretoglobin family (member SCGB3A1) present predominantly in extrapulmonary trachea and bronchi in terminally differentiated epithelial cells. UGRP2 is thought to be involved in maintenance of the terminally differentiated airway phenotype (Porter et al. 2002). In addition, UGRP2 mRNA is regulated by multiple cytokines and is potentially involved in inflammation, immunologic responses, and diseases, such as asthma (Yamada et al. 2005). In humans, UGRP2, also known as high in normal-1, is a candidate tumor suppressor gene and is silenced in various human cancers including lung, breast, and prostate cancers (Krop et al. 2001, 2004).
Surfactant proteins, part of the pulmonary surfactant that lines the air passages in the lung, are phospholipid-associated proteins involved in regulating lung volume and surface tension and are present in Type II cells and alveolar lining (Turell 2008). SP-A, one of the four surfactant-associated proteins, is also expressed in non-ciliated bronchiolar epithelial cells and is involved in host defense systems against pathogens, such as mycobacterium (Lopez et al. 2009) and viruses (van Iwaarden et al. 1991; Benne et al. 1995). In contrast, SP-C is produced primarily by Type II cells in the alveolar compartment and is a hydrophobic membrane protein that is involved in surfactant film formation (Weaver and Conkright 2001) and used in the treatment of neonatal respiratory distress syndrome (Merrill and Ballard 2003).
This study examines gene and protein expression patterns for five secretory proteins of the lung. These secretory proteins were selected because they represent a high-abundance protein for the entire conducting airway tree (CCSP), a low-abundance protein with focal expression in the airway tree (CGRP), a protein that is more abundant in proximal airway regions (UGRP2), and two surfactant proteins with differential compartmental expression between airways and alveoli (SP-A and SP-C). The goal of this study was to demonstrate the enrichment that different methods can afford for defining site-specific gene expression in the lung and to confirm the compartmental-specific gene expression using new methodology.
Materials and Methods
Animals and Treatments
Adult male NIH Swiss mice (10 weeks of age) were purchased from Harlan Laboratories (Hayward, CA) and housed in a high-efficiency particulate air-filtered laminar flow cage rack with a 12 hr light/dark cycle and allowed free access to food and water. Animals were given a 1- to 2-week acclimation period before experiments were conducted and were housed in American Association for Accreditation of Laboratory Animal Care-approved facilities. Animal use protocols were approved by the University of California-Davis Institute for Animal Care and Use Committee. Five to six animals were used for each analysis. Mice were administered corn oil by IP injection at a concentration of 10 μl/g body weight once daily for 7 days. This dosing regimen was chosen as control scenario to precede future studies where chemicals are administered IP in a corn oil vehicle. On day eight, mice were anesthetized with an overdose of Fatal Plus (Vortech Pharmaceuticals; Dearborn, MI) and euthanized by exsanguination.
Tissue Preparation
For collection of whole lung and intrapulmonary airway tree samples, the trachea was cannulated and the lungs were removed and inflated to capacity with RNAlater (Ambion/Applied Biosystems; Foster City, CA) through the tracheal cannula. The lungs were stored at 4C for 24 hr and then moved to −20C until microdissection and RNA isolation could be performed. For whole lung analysis, the right cranial lobe was separated from the remaining lung and processed for RNA isolation. For airway analysis, airways were microdissected from the surrounding parenchyma of the left lobe. This approach allows airways as small as terminal bronchioles to be isolated from the mouse lung (Van Winkle et al. 1996; Baker et al. 2004). All the intrapulmonary bronchi and bronchioles (airways within the lung lobe) were dissected in this manner and pooled for further analysis. RNA isolation from both whole lung and airways was carried out using the RNeasy Plus Mini kit (Qiagen; Valencia, CA) using gDNA eliminator columns.
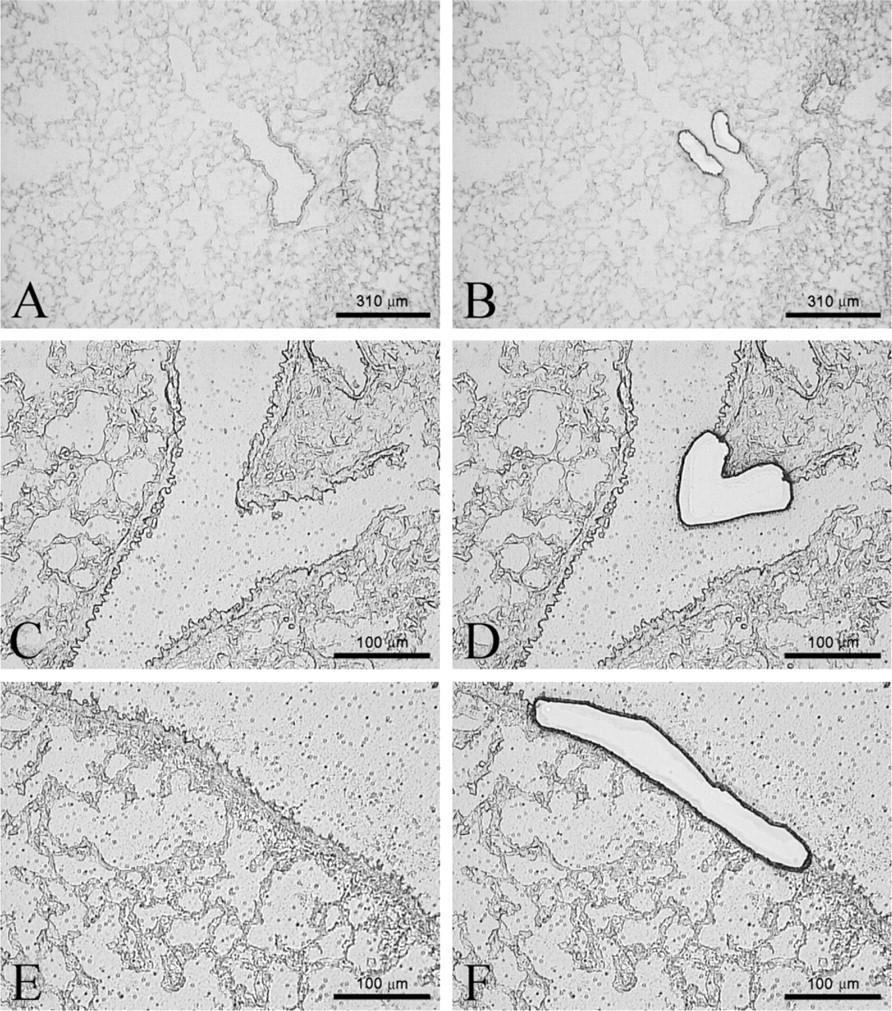
Airway levels captured by laser capture microdissection (LCM). Representative airways before laser capture and post-laser capturing of mouse terminal bronchioles (
Lungs for LCM were prepared by cannulating the trachea and then removing the lung en bloc from the chest cavity. The lungs were inflated to capacity through the cannula with a 50/50 v/v solution of Tissue-Tek O.C.T. compound (Sakura Finetek Europe B.V.; Alphen aan den Rijn, The Netherlands) in RNAse-free PBS. The lobes were cut and placed into cryomolds with excess O.C.T. and cork backing and frozen in 2-methylbutane over liquid nitrogen. Samples were stored at −80C. The cryostat was cleaned with 100% ethanol, and tools and surfaces in contact with the tissue were cleaned with RNase Away (Molecular BioProducts; San Diego, CA) to prepare for cryosectioning. Ten μm sections were collected by cryostat sectioning at −20C onto Leica PEN-Membrane slides (Leica Microsystems; Wetzlar, Germany). Two sections were collected on the same slide. Forty μl of prechilled RNAlater-ICE (Ambion/Applied Biosystems) was pipetted directly onto the slide, and the slide was stored flat in a prechilled slide box at −20C overnight. The next day, slides were rinsed in a Copeland jar of RNAse-free water for 4 min and then desiccated for 10 min/until dry at room temperature. Cells were captured using the Leica LMD6000 Laser Capture Microdissection Unit (Leica Microsystems) by cutting the region of interest into the cap of a 500 μl RNase-free microfuge tube with 25 μl of Buffer RLT Plus (Qiagen). Slides were processed one at a time, taking care that each slide was at room temperature for less than 30 min to protect RNA integrity.
Description of TaqMan Gene Expression Assays (Applied Biosystems)
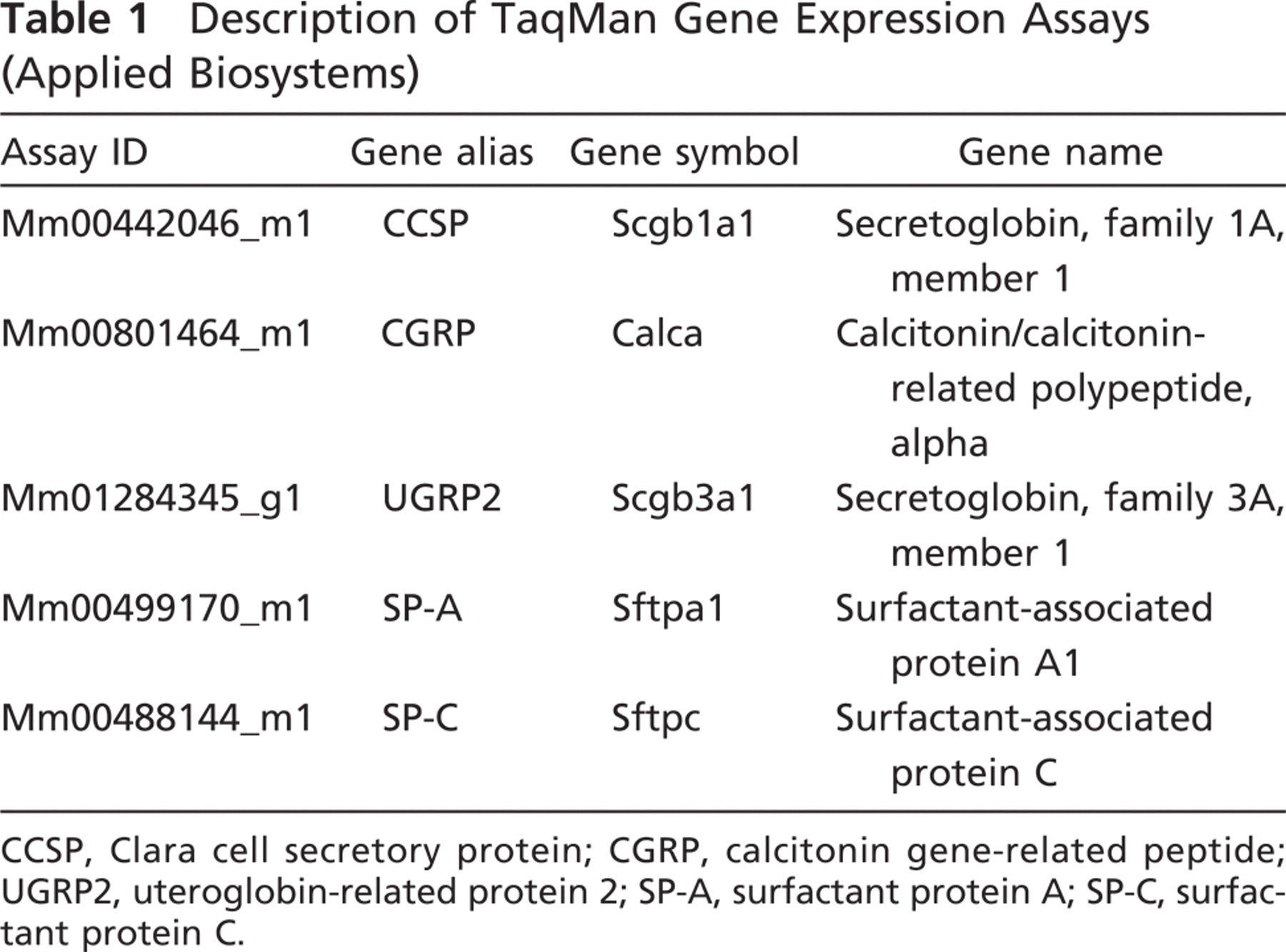
CCSP, Clara cell secretory protein; CGRP, calcitonin gene-related peptide; UGRP2, uteroglobin-related protein 2; SP-A, surfactant protein A; SP-C, surfactant protein C
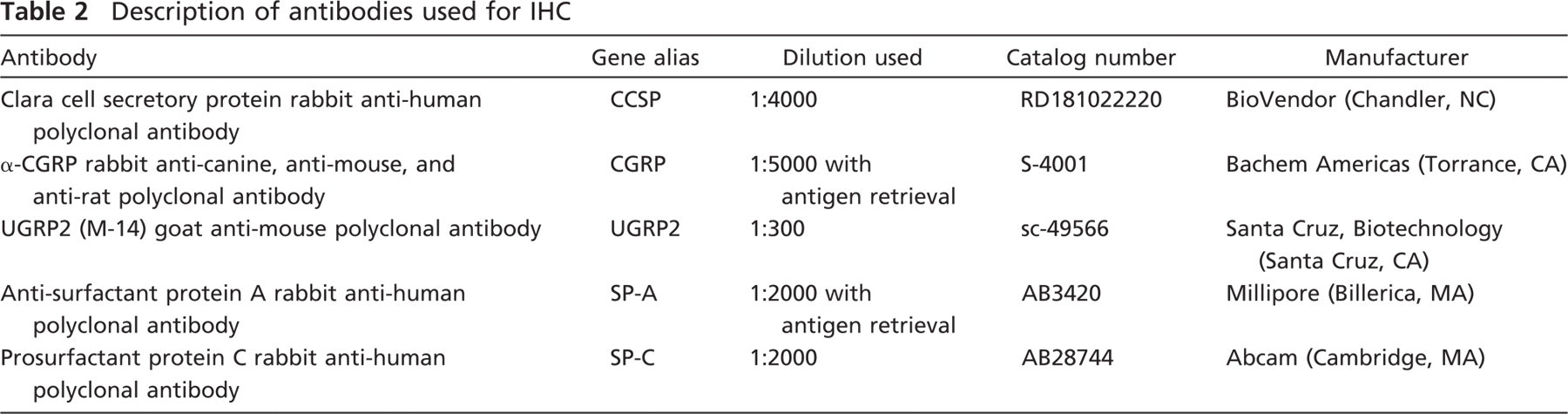
Description of antibodies used for IHC
For this study, terminal bronchioles (TB), airway bifurcations (BF), and mid-level bronchioles, defined as airway levels between main bronchi and distal bronchioles (Lawson et al. 2002), were captured (Figure 1). For each airway level per animal (n = 5–6), sampling was very robust (3–4 tissue faces per section, 2 sections per slide, 10 slides) and areas ranging from 200,000 to two million μm were collected. All TB, BF, and mid-level bronchioles were collected for each section on each slide until two million μm2 were collected and sampling ceased. For each animal and each airway level, the entire sample was collected in a single cap when all LCM sampling occurred in a single session. If a break occurred during capture, remaining sample was collected into a new cap and the contents were later pooled for further analysis. When sample collection was complete, the tube was centrifuged at 13,000 rpm for 2 min. RLT buffer was added to bring the total volume to 75 μl. The sample was frozen at −80C and stored or immediately processed for RNA isolation using the RNeasy Plus Micro Kit (Qiagen) using gDNA eliminator columns.
Samples for IHC were prepared by cannulating the trachea, removing the lungs from the chest cavity, and perfusing through the tracheal cannula for 1 hr at 30 cm constant pressure with 1% paraformaldehyde. The lungs were stored at 4C in 1% paraformaldehyde for 24 hr. The left lobe was sliced into cross sections and embedded in paraffin. Blocks were sectioned (4 μm) onto glass poly-
Methods Validation
To validate our method for RNA preservation before the laser microdissection step, frozen sections were prepared on 10 plain glass slides. Sections were coated with RNAlater-ICE overnight as described earlier, and a razor blade was used to scrape the tissue off the slide. RNA was isolated from the sample. To optimize sample handling methods post-laser capture, mouse lung airway epithelium was captured by LCM from five Leica PEN-membrane slides at two block faces per slide sectioned at 20 μm thickness. Total RNA yield was measured by NanoDrop 1000 (Thermo Scientific Nanodrop Products; Wilmington, DE), and data on comparative quality was determined using a 2100 Bioanalyzer (Agilent Technologies; Santa Clara, CA).
Quantitative Real-time RT-PCR
RNA was reverse transcribed into single-stranded cDNA using the High Capacity RNA-to-cDNA kit (Applied Biosystems) according to the manufacturer's suggested protocol. RT-PCR was carried out in the StepOnePlus Real-Time PCR System (Applied Biosystems). Each reaction contained 2 μl cDNA, 1 μl TaqMan Gene Expression Assay (list of genes assayed and assay ID numbers can be found in Table 1), 10 μl TaqMan Fast Universal PCR Master Mix, No AmpErase uracil-n-glycosylase, and 7 μl of RNase-free water, as recommended by the System software. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as a housekeeping gene. All samples were run in triplicate.
Immunohistochemistry
Paraffin sections were immunostained with rabbit anti-CCSP, anti-CGRP, anti-SP-A, and anti-SP-C and goat anti-UGRP2 (see Table 2). The optimal antibody concentration that gave positive tissue staining with minimal background staining was determined separately for each antibody using a series of dilutions. The avidin–biotin peroxidase method in the form of a Vector ABC kit (Vector Laboratories; Burlingame, CA) was used following the manufacturer's instructions. Antigen retrieval using a decloaking chamber (Biocare Medical; Concord, CA) was carried out for SP-A only. Slides were incubated in 1× AR10 solution (BioGenex Laboratories; San Ramon, CA) in the decloaking chamber for 2 min at 123C, 10 sec at 85C, and then cooled to room temperature. Antigen retrieval using 0.01 M citrate buffer (pH 6.0) was carried out for CGRP (boiled for 15 min and then cooled for 10 min). DAB enhanced with nickel chloride was used as the chromagen with the exception of CGRP where aminoethyl carbazole was used. All tissues to be compared were processed and stained in the same immunochemical run. Tissues were not counterstained except when stained for CGRP where hematoxylin was used. A negative control slide with either preimmune serum or PBS was substituted for the primary antibody for each primary antibody. Images were captured on an Olympus BH-2 microscope (Olympus American; Center Valley, PA). Images were cropped and arrows were inserted using Adobe Photoshop (Adobe Systems; San Jose, CA). No other image manipulation was performed.
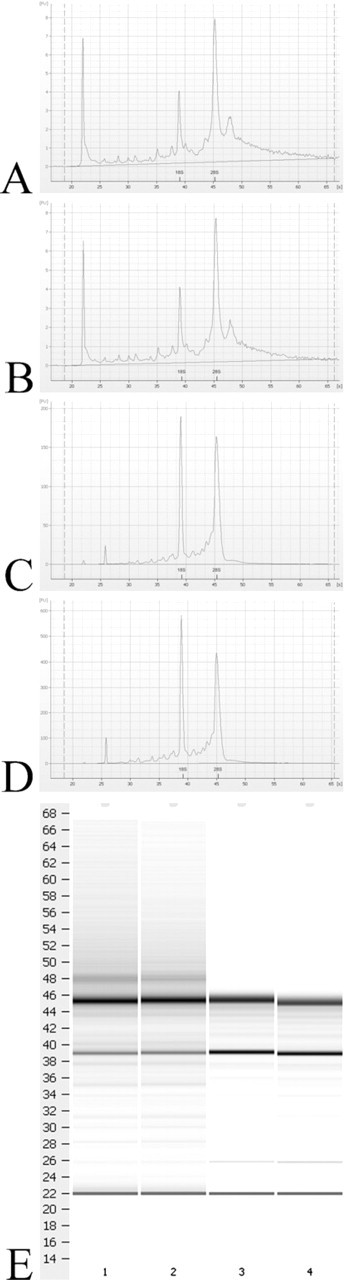
Agilent Bioanalyzer scans of representative ethanol-fixed LCM sample (
Statistical Analysis
Fold change of gene expression in microdissected airways from five–six animals per time point was calculated using the comparative CT(2–ΔΔCT) method as described previously by Applied Biosystems and Livak and Schmittgen (2001). Briefly, for each sample, CT (cycle threshold) value for the reference gene was subtracted from the CT value for the gene of interest to yield a ΔCT value. Next, the ΔCT value from the calibrator group (whole lung) for each gene was subtracted from each individual ΔCT for the same gene. This was the ΔΔCT value. The 2–ΔΔCT value was then determined for each sample and was reported as fold changes relative to whole lung. Groups are reported as mean fold change ± 1 SD. Statistical calculations were carried out using StatView (SAS; Cary, NC). Results for each gene were analyzed by ANOVA with Scheffe's test. p Values less than 0.05 were considered significant.
Results
Methods Development
Samples taken before LCM were assessed by scraping the slides to establish a baseline yield of RNA at this phase of sample processing. The resulting yield was 4.5 μg total RNA with a 260/280 ratio of 2.1 using RNAlater-ICE. Incorporation of RNAlater-ICE produced both high quality and high yields. Furthermore, slides treated with RNAlater-ICE rather than ethanol took less time both to prepare and to laser. Slides fixed with ethanol took an average of 11 min to fix, rinse, and dessicate and 13 min per slide to laser vs 8 min to prepare and 5 min per slide to laser when fixed with RNAlater-ICE. Increased laser times were due to the need to recut an average of 73% of the selected regions on the ethanol-fixed slides, whereas only 22% of the regions on the RNAlater-ICE slides required recutting. This was at similar power, aperture, and speed setting (average of 47, 12, and 10, respectively). Increasing power to achieve a lower recutting average was attempted for the ethanol-fixed slides, but the width of the laser at higher power settings destroyed large amounts of tissue surrounding the area of interest. An alternative method attempted was to incorporate RNAlater into the inflation step for RNA stabilization, but this was unsuccessful as blocks prepared in this manner were not fully frozen at −20C likely due to the high salt content of this solution.
Optimization of the sample handling methods was confirmed by Bioanalyzer comparing RNA quality from (1) the new method incorporating RNAlater-ICE on tissue sections, (2) ethanol fixation of LCM sections, (3) gross airway microdissection, and (4) whole lung homogenate of tissue inflated and stored in RNAlater. No significant degradation of the LCM samples with either method was found, and RNA integrity numbers were 7.9 or above for all methods tested (Figure 2). This demonstrates that the method for RNA stabilization in frozen tissue is highly effective and that RNA can be isolated from laser-captured tissue with high quality and without significant degradation as described in Materials and Methods.
TaqMan gene expression analysis of each sample was also carried out, and amplification plots and CT (cycle where threshold was reached) values for GAPDH and peroxiredoxin 6 were detected at >99% assay efficiency. CT values for GAPDH were also evaluated to validate its use as a housekeeping gene in the experiments. cDNA from airway-only samples and from whole lung samples were compared by using RT-PCR. When the same input amount of cDNA was used per reaction (100 ng/μL), CT values were 22.895 ± 0.135 for airways and 22.218 ± 0.189 (means ± SD) for whole lung samples, with no significant difference in CT values, indicating no significant differences in GAPDH expression between the airway and parenchyma compartments. Thus, GAPDH was considered acceptable as a housekeeping gene for the study.
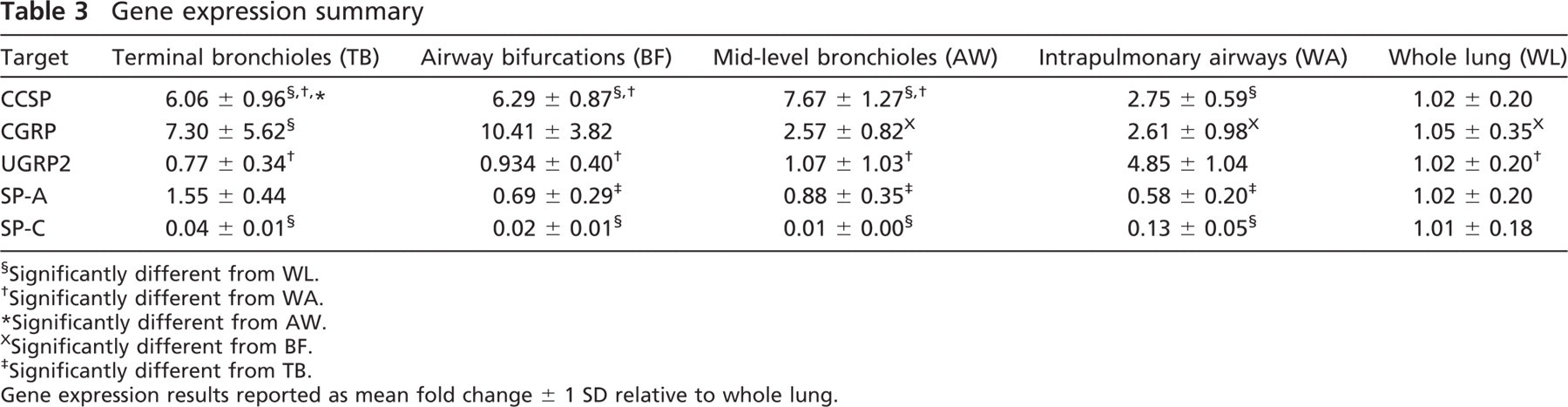
Gene expression summary
Significantly different from WL.
Significantly different from WA.
Significantly different from AW.
Significantly different from BF.
Significantly different from TB.
Gene expression results reported as mean fold change ± 1 SD relative to whole lung.
CCSP Gene and Protein
CCSP mRNA expression was significantly more abundant in LCM and airway microdissection at all airway levels (whole airway, p = 0.0019; terminal bronchioles, p < 0.0001; mid-level bronchioles, p < 0.0001; airway bifurcations, p < 0.0001) relative to whole lung (Table 3). All airway levels obtained from LCM (terminal bronchioles, mid-level bronchioles, and airway bifurcations) were significantly increased from the whole airway tree (p < 0.0001), and within LCM samples, mid-level bronchioles were significantly increased from terminal bronchioles (p = 0.0048). Airway microdissection increased the detection of CCSP by almost 3-fold, and LCM increased the detection of CCSP mRNA by over 6-fold at all airway levels (Figure 3A). CCSP protein was consistently expressed in the Clara cells of the conducting airways in all airway generations, from proximal intrapulmonary bronchi to the most distal terminal bronchioles, and was not detected in the alveoli (Figures 3B–3D).
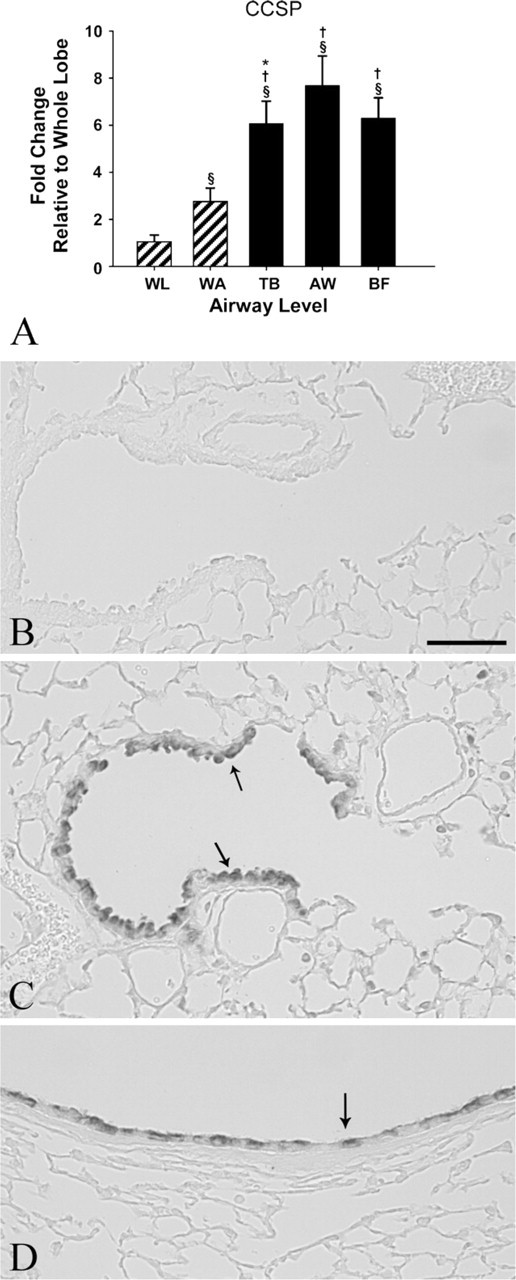
Relative gene and protein expression of Clara cell secretory protein (CCSP). (
CGRP Gene and Protein Expression
Expression of CGRP mRNA was enriched in airway bifurcations 10-fold over the whole lung and 8-fold over the whole airway tree and mid-level bronchioles (p < 0.0001, p = 0.0003, and p = 0.0008, respectively). Additionally, LCM sampling of terminal bronchioles showed a significant 7-fold increase over whole lung (p = 0.0024; Table 3, Figure 4A). CGRP protein expression was detected sparsely in terminal bronchioles and mid-level airways (Figures 4C and 4E) and in discrete clusters of airway epithelial cells at airway bifurcations (more proximal airway generations had greater frequency of having CGRP-positive cells) (Figure 4D).
UGRP2 Gene and Protein Expression
mRNA expression of UGRP2 was significantly increased by almost 5-fold in the whole airway tree compared with whole lung, terminal bronchioles, and airway bifurcations and by 3-fold over mid-level bronchioles (p < 0.0001) (Table 3, Figure 5A). UGRP2 protein expression was enriched throughout the proximal airway epithelium being the most abundant in the trachea, present in lesser quantities in the larger intrapulmonary proximal airways, and highly localized to the apical surface of individual airway epithelial cells. Staining in the smaller distal airways and terminal bronchioles was sparse to undetected (Figures 5C–5E).
SP-A Gene and Protein Expression
mRNA expression of SP-A was significantly elevated in terminal bronchioles compared with the whole airway tree, mid-level airways, and airway bifurcations (p < 0.0001, p = 0.0023, and p = 0.0003, respectively) (Table 3). Furthermore, SP-A levels were greater in the whole lung compared with the whole airway tree, although it was not statistically significant (Figure 6A). SP-A protein expression was detected with a similar distribution and abundance as mRNA throughout the airway tree (Figures 6C–6D).
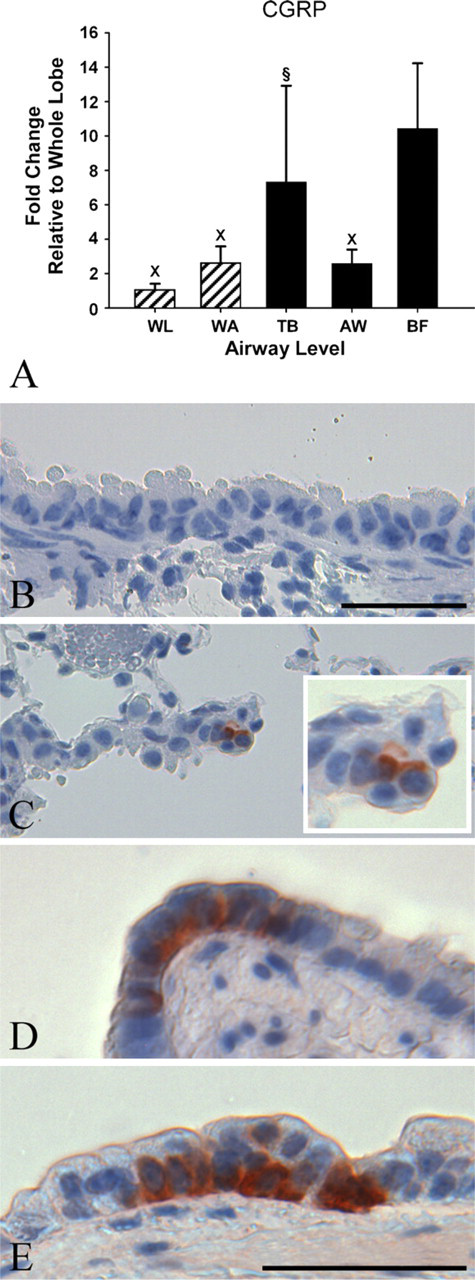
Relative gene and protein expression of calcitonin gene-related peptide (CGRP). (
SP-C Gene and Protein Expression
Detection of SP-C mRNA was significantly and highly enriched in the whole lung compared with all other lung compartment approaches (p < 0.0001) (Table 3, Figure 7A). Immunohistochemical detection of the pro-SP-C protein was localized to the parenchyma Type II cells (Figure 7C).
Discussion
This study demonstrates that mRNA expression differs by lung region (including airway level) and the method and site of sample collection for CCSP, UGRP2, CGRP, SP-A, and SP-C can alter the detection of mRNA. These results are indicative of the relative utility of different microdissection approaches for studying site-specific gene expression in the lung. Our hope is that a thorough description of these approaches and demonstration of their utility will enhance research into site-specific changes in the lung. These methods are especially critical for the study of air pollutants, such as particulate matter or oxidant gasses, which have site-specific deposition and toxicity patterns in the tracheobronchial airways. These approaches can be utilized to study airway epithelial injury and repair responses in the lung as well as mechanisms of lung development because these events are highly airway level and lung micro-environment specific.
It has been demonstrated that mouse lungs can be inflated with RNAlater and microdissected grossly to isolate different airway levels for gene expression analysis (Baker et al. 2004). Storage of tissue samples in RNAlater has been shown to be the most effective method of preserving RNA integrity, even superior to flash freezing in liquid nitrogen (Keating et al. 2008). Because of the ease of use and the advantages for integrity of lung RNA, instillation of or immersion in RNAlater is now routinely used to preserve respiratory tract samples (Wagner et al. 2009). However, because LCM necessitates sectioning of tissue onto a slide, frozen cryosections or paraffin sections are typically used depending on the downstream applications. For gene expression analysis of samples obtained by LCM, it is widely accepted that frozen tissue yields better quality RNA than formalin-fixed, paraffin-embedded tissue, although morphology is less preserved (Mora et al. 2002). In a previous study, fixation in 70% ethanol improved morphology and provided good RNA extraction over other fixation and embedding methods (Goldsworthy et al. 1999). However, the results from this study revealed that ethanol fixation resulted in tissue that was more difficult to cut with laser on the Leica LMD6000 platform and there were also concerns with room temperature stability of these samples during the laser capturing process.
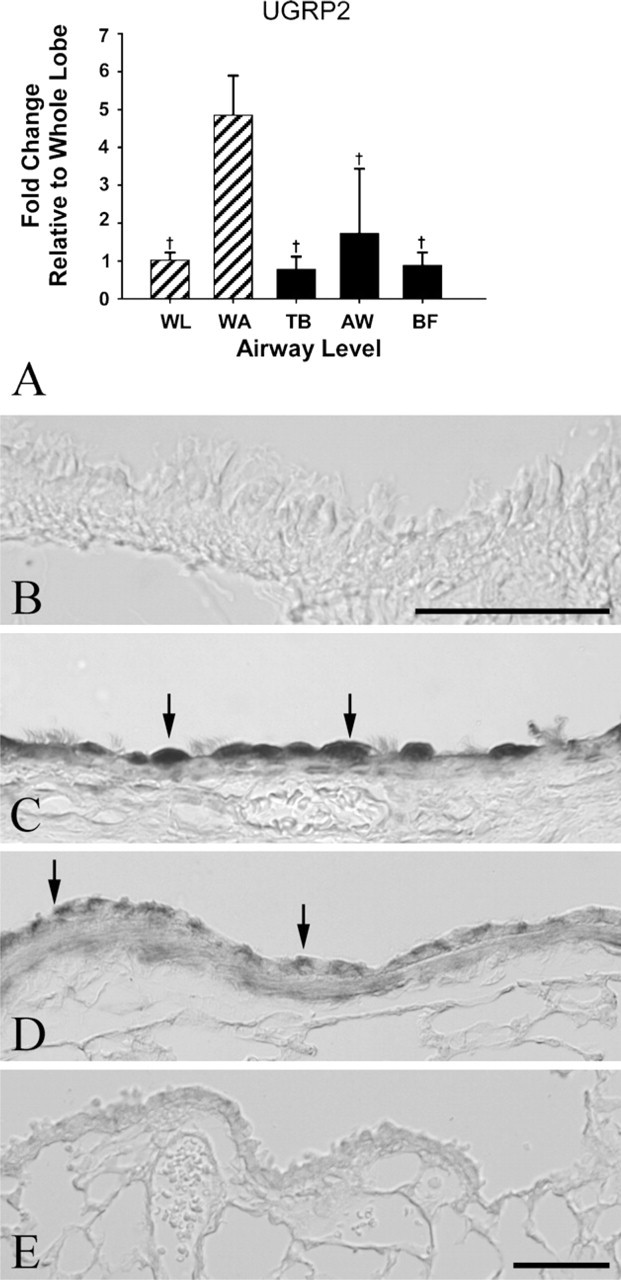
Relative gene and protein expression of uteroglobin-related protein 2 (UGRP2). (
This study describes an improvement over current approaches for laser capture of frozen tissue section that enhances RNA stabilization in frozen tissue sections: use of RNAlater-ICE for RNA preservation after cryosectioning. This method provided consistently good RNA yield and RNA integrity. Further, this approach saved time during sample preparation and during laser capture compared with conventional methods using ethanol fixation. This approach allows sufficient yield for analysis of numerous genes from a single sample without the amplification step that is common in other studies (Betsuyaku and Senior 2004; Adair-Kirk et al. 2008; Klee et al. 2009).
CCSP gene expression was enriched in all airway samples compared with the whole lung. This is not unexpected due to the localization of CCSP to the Clara cells of the conducting airways. However, the enhanced fold change detected in the LCM samples (6-fold over whole lung and 3-fold over whole airway tree) and the ability to detect significant differences within the LCM samples by sublocation with the airway tree (terminal bronchioles from mid-level bronchioles, p = 0.0048) demonstrates the benefit of using LCM to detect changes in mRNA expression in research involving diseases of the distal lung. In human studies, CCSP levels in serum and lavage fluid serve as biomarkers of lung disease and lung injury (Shijubo et al. 1997; Broeckaert et al. 2000). In a transgenic mouse model used to study lung tumors arising from Clara cells, CCSP expression was found to be diminished in tumor cells (Hicks et al. 2003). In mouse models of lung toxicity to naphthalene, a polycyclic aromatic hydrogen compound metabolically activated by cytochrome P450s in the Clara cells of the conducting airways, toxicity to the Clara cells was distributed in a site-specific manner depending on route of exposure and dose resulting in focal losses of the CCSP-expressing cell population (Warren et al. 1982; Plopper et al. 1992; Van Winkle et al. 1995). CCSP is involved in the response of the lung to oxidant stress; CCSP-knockout mice have increased cellular injury compared with wild-type mice when exposed to hyperoxia or ozone (Johnston et al. 1997). With toxicants and disease states that affect specific regions, populations, or cell types, use of new methodology such as LCM for site-specific sampling has given additional insight into cellular changes that occur in the lung, but has been limited by low yields, the labor-intensive nature of these studies, and the need to amplify the mRNA obtained. This study advances the field by applying new approaches that enhance yield.
The increased mRNA expression of CGRP in airway bifurcations (10-fold over whole lung) is supported by previous studies that describe a population of CGRP-labeled cells at airway branch points (Peake et al. 2000; Reynolds et al. 2000). However, the 7-fold increase observed in the terminal bronchioles has not been previously reported. This is especially interesting because terminal bronchioles were not examined in previous studies that investigated the role of CGRP-expressing cells in cellular proliferation and epithelial repair in response to naphthalene exposure, a cytotoxicant that disproportionately targets terminal bronchioles (Stevens et al. 1997; Peake et al. 2000). It is not known if a similar result would also be obtained for other site-specific toxicants, such as 4-ipomeanol and coumarin, both of which target Clara cells in terminal bronchioles via cytochrome P450 activation to reactive chemical intermediates that cause cellular toxicity (Boyd 1977; Born et al. 1998). In this study, the increased variability in the terminal bronchioles compared with airway bifurcations suggest that although bifurcations consistently express CGRP, mRNA expression of CGRP in terminal bronchioles is not consistent and this may have implications for epithelial repair and regeneration in response to cytotoxic exposure targeting the terminal bronchioles. This is supported by IHC data showing sparse CGRP staining in the terminal bronchioles. It is possible that the airway epithelium in the terminal bronchiole generates CGRP that is secreted and migrates to other sites in the airway. Because of the pattern and specificity of CGRP expression in the lung and the proposed functions of CGRP in airway regeneration, site-specific approaches such as LCM to determine alterations in CGRP gene expression in response to exposure or disease state increase the ability to define and detect these changes.
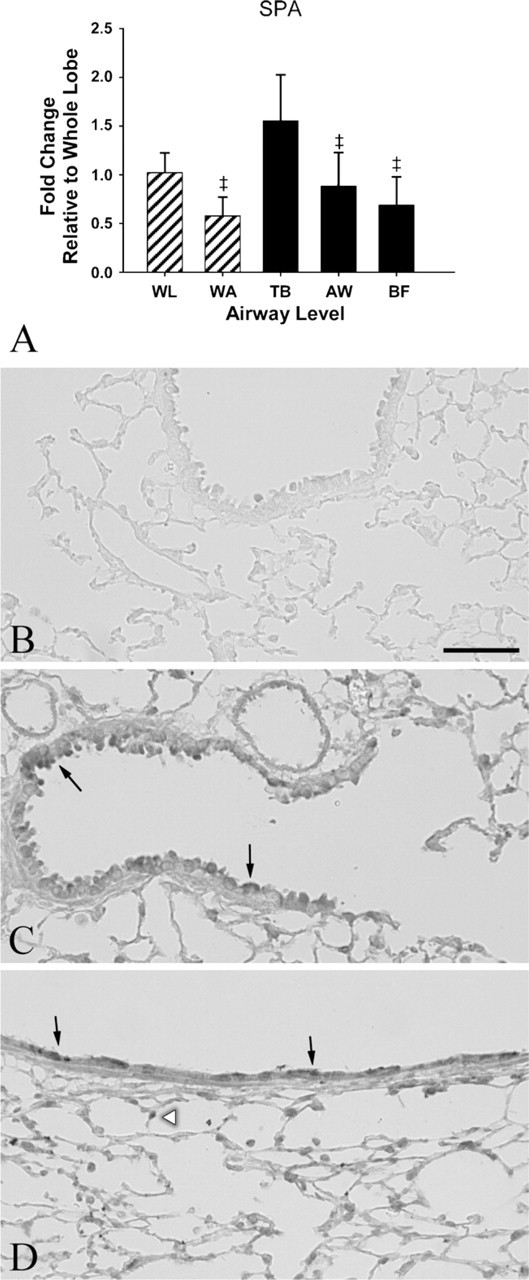
Relative gene and protein expression of surfactant protein A (SP-A). (
Detection of significantly elevated mRNA expression of UGRP2 in the whole airway tree (5-fold) but not in the terminal bronchiole and airway bifurcations supports the previously published reports of UGRP2 expression being localized to proximal airways (Porter et al. 2002; Reynolds et al. 2002). LCM of mid-level bronchioles introduced an increase in variability of UGRP2 mRNA detection compared with all other samples. This is likely due to the broad definition of mid-level bronchiole in our sampling, resulting in a larger and more variable sampling region. Perhaps, a more stringent definition of airway levels to include a range of airway generations or sampling of airways in the presence of smooth muscle would decrease the variability observed. However, because the methods used to map airway generations require microdissection of tissue before embedment, we were unable to adapt those methods for LCM. Immunohistochemical data showing sites in the trachea, lobar bronchus, and intrapulmonary proximal large conducting airways with positive staining for UGRP2 and decreased staining in a proximal to distal manner support the gene expression data. This suggests that airway microdissection of the trachea and proximal intrapulmonary conducting airways would likely result in maximal enrichment of UGRP2 message.

Relative gene and protein expression of surfactant protein C (SP-C). (
SP-A is the major protein in surfactant and is involved in host defense in the lung (Kingma and Whitsett 2006; Serrano and Perez-Gil 2006; Linke et al. 2009). It is also expressed in some non–small cell lung carcinomas, reduced or absent in other pulmonary diseases, such as fibrosis and respiratory distress syndrome, and its localization and detection in tissue may aid in the diagnosis and prognosis of these conditions (Goldmann et al. 2009). This study shows enrichment of SP-A mRNA expression in laser-captured terminal bronchioles when compared with other airway levels examined. Although enrichment is only 2-fold greater than the whole airway tree and not significantly different from the whole lung, this could potentially be important as terminal bronchioles can be targets of inhaled toxicants and infectious agents. SP-A-knock out and SP-A-deficient mice are found to be more susceptible to certain pathogens than their wild-type counterparts (LeVine et al. 2002; Ledford et al. 2009; Linke et al. 2009) and thus site-specific approaches to changes in gene expression of SP-A may give insight into pathogenesis of lung disease.
SP-C mRNA expression was found to be 25- to 100-fold less in microdissected and LCM-sampled airways than in the whole lung, which is consistent with the literature stating SP-C is transcribed exclusively by Type II cells in the lung (Mulugeta and Beers 2006). The ability to detect very small amounts in the LCM samples may come from small amounts of parenchyma below the basement membrane or behind the cells visualized and captured on the LCM. Variance between all airway levels and methods was also very small. Deficiencies and mutation in the SP-C gene, SFTPC, have been found to contribute to interstitial lung disease, neonatal respiratory failure, and increased susceptibility to viral infection (Cameron et al. 2005; Soraisham et al. 2006; Glasser et al. 2009). Because of the specificity in mRNA expression of SP-C in the Type II cells, a whole lung- or parenchyma-specific approach to tissue isolation would be an appropriate sampling scheme. The same recommendation could be made for chemicals such as 3-methylindole, which targets Type I alveolar epithelial cells (Durham and Castleman 1985).
Variability within samples is always a concern, and in some cases, LCM increased the variability of relative mRNA detected. We do not believe that the increase in variability observed diminishes the strength and utility of LCM use. Earlier morphometric studies (Hyde et al. 1990) indicate that 70% of all variance in animal experiments is due to animal/animal variance. Airway-to-airway variability is most likely depending on the location in the axial path and lobe. Thus, perhaps the variability observed in this study was in part due to a combination of these factors, but this is likely the case with any site-specific methodology.
In summary, this study demonstrates the relative utility of different approaches to defining lung mRNA expression and newly quantifies the gene expression pattern of several secreted proteins in the conducting airways of the mouse lung. Both LCM and gross airway microdissection have advantages compared with each other and are superior for detection of airway-specific proteins compared with a whole lung homogenate. Gross airway microdissection of RNAlater-inflated tissues is faster to achieve sufficient sample and enriches the detection of mRNA in the airway epithelium of the conducting airways, but is less precise than the laser capture approach. The limitation of laser capture is the time and effort it entails relative to the amount of the sample that can be obtained for analysis without amplification. With the diversity of approaches available with both advantages and limitations to each, knowing the cell type of interest or regions affected in an experimental exposure or diseased state will aid in choosing the appropriate method to enrich detection of changes in gene expression.
Footnotes
Acknowledgements
This work was supported in part by a National Institute of Environmental Health Sciences training grant ES007059 (to KMS), NIH R01 ES012720, TRDRP 14RT-0132, and FAMRI. Although the research described in the article has been funded in part by the United States Environmental Protection Agency through grant RD-83241401–0, it has not been subject to the Agency's required peer and policy review and therefore does not necessarily reflect the views of the Agency and no official endorsement should be inferred.
We thank Dr. Suzette Smiley-Jewell for editing the manuscript. The following core facilities at University of California Davis were used for this project: Cellular and Molecular Imaging at the Center for Health and the Environment, The UC Davis Cancer Center Gene Expression Resource, and the UC Davis Genome Center Expression Analysis Core.