
Abstract
Fluorescence in situ hybridization to DNA fibers (Fiber-FISH) is a high-resolution, wide-ranging physical DNA mapping method that finds increasing application in the study of pathological gene rearrangements. Here we present experiments designed to understand the nature of the discontinuous FISH signal patterns seen after Fiber-FISH. Use of a novel cisplatin-based chemical labeling method enabled us to produce intact biotinlabeled cosmid target DNA molecules. We monitored by immunofluorescence the fate of such cosmid targets during denaturation and hybridization. The same cosmid DNA labeled with digoxigenin by nick-translation was used to analyze the FISH probe signal distribution in a different color. The probe signals proved to be a subset of the target signals remaining after denaturation and hybridization. We argue that the discontinuity of probe signals in Fiber-FISH is mainly caused by loss of target DNA and limited accessibility due to in situ renaturation and attachment. Furthermore, we conclude that FISH sensitivity is determined by hybridization efficiency and not the ability to generate sufficient signal from small probes.
D
Typically, large insert clones such as cosmids, bacterial, and yeast artificial chromosomes are used as hybridization probes in Fiber-FISH and specific hybridization to whole genomes or cloned DNA targets is recognized as linear or near-linear stretches of beads-on-a-string fluorescence signals. Factors such as loss and inaccessibility of target DNA due to either in situ renaturation or attachment to the glass substrate, as well as suppression of repeat sequences, have been put forward as contributing to the origin of the typical discontinuous Fiber-FISH pattern (Wiegant et al. 1992; Windle et al. 1995). However, experimental assessment of the factors determining the nature of the pattern is lacking. Here we report experiments aimed at a better understanding of the origin of the beads-on-a-string character of Fiber-FISH signals by monitoring a cloned target DNA in terms of DNA loss and accessibility during the FISH procedure. For this purpose we used a chemical nucleic acid labeling technique based on cisplatin that, unlike nick-translation or random primed labeling, leaves the sugar-phosphate backbone intact (Wiegant et al. 1999). Thus, intact labeled target DNA molecules can be produced for monitoring their fate during Fiber-FISH.
Materials and Methods
Cosmid cAL24 (Florijn et al. 1995) was modified to about 10% with biotin-ULS according to the manufacturer's instructions (Kreatech Diagnostics; Amsterdam, The Netherlands). Biotin-ULS-labeled cosmids were subsequently immobilized on aminosilane-coated slides using the receding meniscus technique (Bensimon et al. 1994). The immobilized DNA was analyzed microscopically before denaturation for biotin stainability in a three-layer detection scheme consisting of streptavidin-Texas Red (Vector; Burlingame, CA) biotinylated goat-anti-streptavidin (Vector), and streptavidin-Texas Red (Florijn et al. 1996). After denaturation, the biotinated target cosmids were hybridized with the same cosmid DNA labeled by nick-translation with digoxigenin-dUTP, after which the biotin and digoxigenin moieties were detected with Texas Red and fluorescein, respectively. The Texas Red detection of biotin was as above. For the detection of digoxigenin the following three-layer detection scheme was used: mouse anti-digoxin (Sigma; St Louis, MO), rabbit antimouse-FITC (Sigma), goat anti-rabbit-FITC (Vector) (Florijn et al. 1996).
Fiber-FISH results were imaged by digital fluorescence microscopy according to Vrolijk et al. (1996). Because of shearing, a considerable fraction of the cosmid molecules was linear. Only linear molecules were imaged.
Results
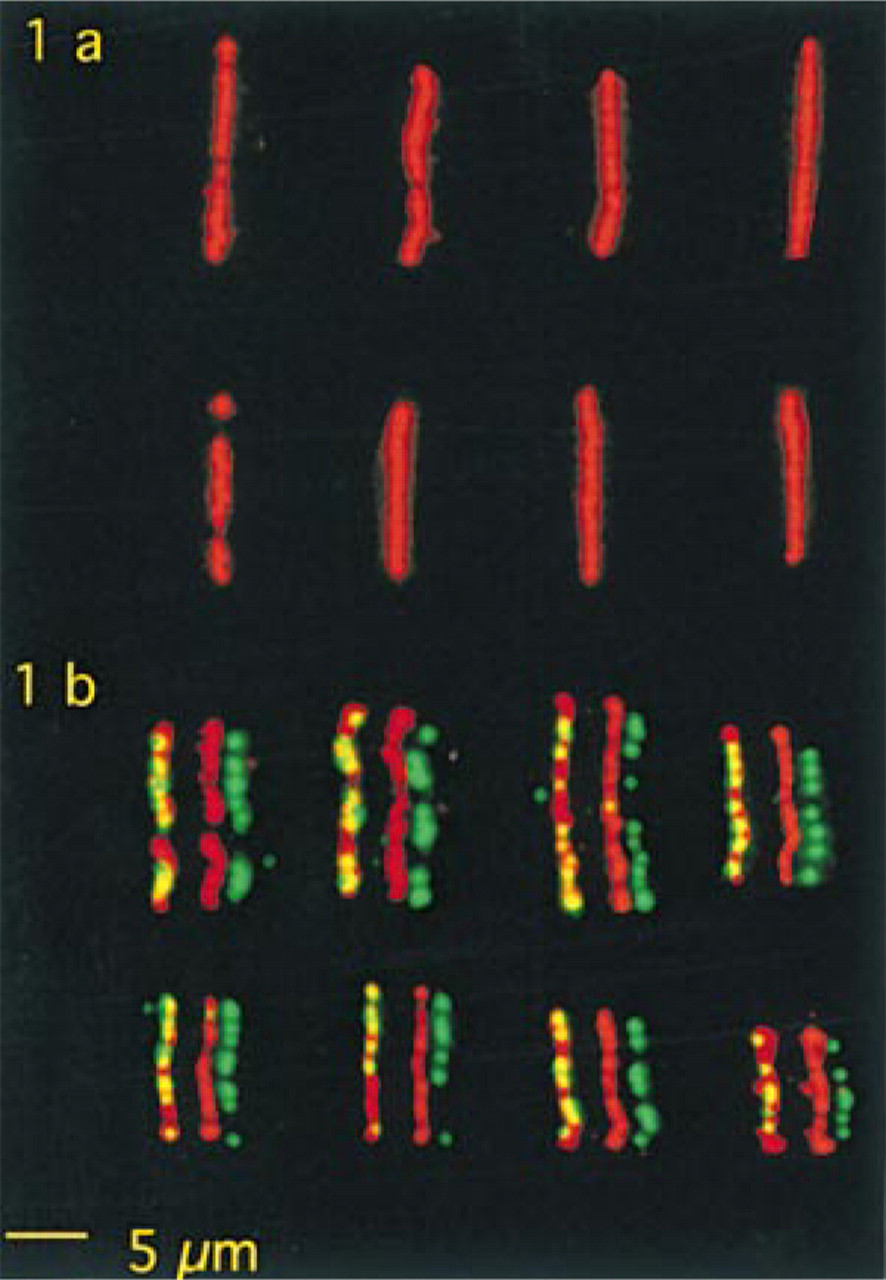
Figure 1a shows images of biotinated target cosmid molecules before denaturation and hybridization and Figure 1b of target and probe cosmid signals after denaturation and hybridization. The overall fluorescence intensities of denatured and hybridized target cosmid molecules are much lower than those of undenatured targets. After denaturation alone, signal distribution and intensities were similar to those of target cosmids that were denatured and hybridized (not shown).
Discussion
Because it is unlikely that inaccessibility for biotin detection reagents will decrease with denaturation, the reduction in the signal intensities of the target cosmids after denaturation can be accounted for by DNA loss.
When the distribution of the hybridization signals is compared with that of the target (Figure 1b), it is clear that there are stretches of target signals not bearing any hybridization signal and that there is no consistent signal distribution pattern. Because the present homologous cosmid DNA hybridizations were conducted without unlabeled Cot-1 DNA, it can be concluded that the typical beads-on-a-string Fiber-FISH pattern seen in Figure 1b does not originate from suppression of repeats but rather from technicalities in the FISH protocol. It should be noted, however, that in Fiber-FISH to whole genomes using large insert clones as probes in the presence of Cot-1 DNA for suppression of repeats, the sizes of tandemly repeated sequences may be such that they can contribute to the appearance of gaps. Consistent appearance of large gaps with Fiber-FISH to whole genomes has indeed been reported (Windle et al. 1995).
The fact that the hybridization signals are a subset of the remaining target signals strongly indicates that, next to DNA loss, accessibility of target DNA is a main determinant of hybridization efficiency. This inaccessibility may be due to the process of in situ renaturation or to physical attachment to the glass surface per se.
We conclude that the relative scarcity of the hybridization signals, as seen in these experiments, is a consequence of DNA loss during denaturation, in situ renaturation of target, and limited reagent accessibility due to attachment. This implies that a target fragment has a random chance of obtaining a FISH signal. This randomness explains the absence of consistent beads-on-string signal patterns in the present experiments. As noted previously, suppression of repeat sequences may contribute to the appearance of gaps in Fiber-FISH to genomic DNA.
(
Previously, we have detected sub-kb targets by genomic Fiber-FISH and have noted a decrease in efficiency of detection as target sizes became smaller. Nevertheless, the FISH intensities of the smallest target tested (202 bp) were detected with an efficiency of about 30%, indicating that smaller sequences should be detectable as such, even with the standard indirect FISH techniques used (Florijn et al. 1996). A similar Fiber-FISH efficiency has been reported recently for ESTs of about 300 bp using highly sensitive tyramide-based detection protocols (Horelli-Kuitunen et al. 1999). It is concluded from these observations that it is hybridization efficiency that determines FISH sensitivity rather than the ability to generate sufficient signal from small probes.
It is noteworthy that FISH with cosmid probes to metaphase and interphase cells leads to single signals at the target sites with intensities comparable to those of a single signal in a cosmid Fiber-FISH track, indicating that, because of a higher level of inaccessibility, hybridization efficiencies in interphase cells and chromosomes are much lower than those of Fiber-FISH, perhaps by as much as a factor of 10.
Footnotes
Acknowledgements
We thank Kreatech Diagnostics for the biotin-ULS reagent.