
Abstract
It has been recently noticed that dust originating from deserts can be transported to other continents by the atmosphere and has an adverse effect on public health, such as increased asthma attacks. Dust originating from the Saharan Desert could initiate a series of reactions upon contact with cloud water and results in the formation of reduced iron (Fe2+), oxalate and various basic amino acids. We aimed to evaluate whether the simulation of Saharan dust-containing atmospheric conditions could trigger the trigeminovascular system. Freely moving rats incubated within simulated atmospheric conditions containing (i) Saharan dust, (ii) Co60 gamma ray-treated Saharan dust (sterilized) and (iii) dust-free air, were investigated for the presence of c-fos expression in trigeminal nucleus caudalis (TNC) and for NOx (nitrate+nitrite) levels in blood samples. Atmospheric samples were analysed for microorganisms. Saharan dust-containing atmospheric conditions induced c-fos expression in nociceptive neurons within TNC. The number of c-fos+ neurons in superficial lamina of TNC was significantly higher in the Saharan dust group (32.9 ± 5.3, P = 0.0001) compared with dust-free air (11.02 ± 2.7) or Co60-treated Saharan dust groups (15.01 ± 2.4). An increase in NOx levels was detected in blood samples of rats exposed to Saharan dust-containing atmosphere. This study has revealed an unknown environmental factor as a possible trigger for headache. It is the first time that transport of Saharan dust with the atmospheric air stream has been documented to be able to trigger the trigeminovascular system in animals. Further studies are needed to explore the mechanisms and molecules that mediate the nociceptive effect and to guide new treatment strategies.
Introduction
Migraine seen in approximately one-fifth of the population is one of the most debilitating diseases, deeply affecting the quality of life and leading to loss of work due to headache and accompanying symptoms. Migraine headache is a neurovascular disorder in which the attacks could be triggered by internal and external factors in genetically susceptible individuals. One of the environmental factors is weather conditions, and studies have revealed that 7–43% of patients are susceptible to atmospheric changes (1–3). Several studies have shown that atmospheric conditions can affect well-being and, particularly, headache/migraine is reported to be most frequent symptom (61%) (4, 5). Atmospheric weather changes including, pressure, humidity, wind and temperature were all implicated as a potential trigger for headache, though a definite link could not be demonstrated. Low barometric pressure and low temperature have been associated with increased pain intensity in humans as well as experimental animals (6, 7). Emergency department visits for headache are correlated with air pollution-related increases in levels of SO2 (sulphur dioxide), NO2 (nitrogen dioxide), CO (carbon monoxide) and PM 2.5 (particulate matter < 2.5 μm) (8). The effect of Saharan desert dust-laden atmospheric conditions on headache has not been implicated before.
It is known that 1–1.5 billion tons of dust are thrown from the Saharan desert and scattered to the atmosphere each year (9). Besides closer continents such as Europe and Asia, Saharan desert dust effects transatlantic regions, primarily the Caribbean and America, including the USA and beyond. Satellite observations have revealed that there is frequent transport of Saharan dust as pulses in various directions, including Turkey, and regional studies have confirmed that the main path of the dust-carrying winds has a westerly (87%) or southwesterly (61%) component (i.e. from North Africa) (10). Studies performed in the east of the Mediterranean have confirmed that 20 million tons of Saharan and African dust are transported to Anatolia each year (11–13). A typical Saharan dust episode affecting Anatolia as recorded by satellite is given in Fig. 1.

A typical Saharan dust transport episode affecting Anatolia Black Sea Caucasus and beyond, as recorded by satellite. Image courtesy of MODIS Rapid Response Project at NASA/GSFC.
Dust particles have a spectrum of 0.01 to hundreds of microns of particulate size. The Environmental Protection Agency in the USA has determined standards for air-borne dust, and dust is classified as PM10 (particulate matter up to 10 μm) and PM2.5. Particles up to 10 μm can accumulate in the airway and particles up to 4 μm can affect pulmonary function by gas exchange.
Particles < 2.5 μm have the most important effect on human health. These particles by-pass the filtration mechanism of the lung and carry infective agents directly to lung tissue and the subepithelial environment, which causes reduction of functional capacity by surrounding internal tissue (14, 15). It was shown that various microorganisms can be carried a long distance by simply sticking to dust. Griffin and Gadd (15, 16) have recently shown that dust-borne microorganisms and other associated constituents may also play a significant role in regional human and ecosystem health. Health studies, on the other hand, have mainly concentrated on the risk of exposure to particulate matter of anthropogenic vs. that of crustal origin (8). Geographical regions such as the Middle East and the Caribbean are impacted by desert dust storms and known to have had some of the highest incidence of asthma during the last two decades, which has coincided with an increased flow of Saharan dust towards the region (17, 18).
Studies dealing with intercontinental dust transport issues have thus far concentrated either on the transport mechanisms, size and chemical composition of dust, or recently on its microorganism composition. It has also been shown that desert top soils are also laden with viable and diverse prokaryote communities (19–21). On average, one gram of topsoil contains 107–109 prokaryotes (22) compared with a mere 1011 stars in our galaxy.
Saydam and Senyuva (23) have shown that the bacteria and fungi content of Saharan dust has an essential role besides its specific clay and iron mineralogy. Significant alterations take place along the intercontinental transport route of the dust veil upon contact with cloud water. The microorganisms found in the dry dust are activated when exposed to cloud water and form oxalate as an osmosolute within 15 min. Thus, Saharan dust can produce its own oxalate indirectly, after being exposed to cloud and water (23). This may be the reason for the observation and suggestion of in-cloud processes as the principal pathways to formation of dicarboxylic acids (16, 24, 25).
Following the in-cloud formation of oxalate, bacteria and fungi attach themselves onto the clay minerals by forming iron oxalate. If solar irradiation along the transport route is above some threshold energy level, charge is transferred from ligand to metal and iron oxalate is metabolized by decarboxylation, resulting in one mole of reduced iron, one mole of carbon dioxide and one mole of carbonyl radical, which can trigger yet another recycling mechanism to dissolve with another surface, oxygen or Fe3+ (26). Stochiometrically reduced iron is unstable, therefore it tends to be utilized immediately, and the surrounding veil of bacteria and fungi are ready to utilize iron in its reduced form. Recent studies have further shown that in rain water, organic nitrogen is associated with atmospheric dust originating from the Sahara desert. Although amino nitrogen contributed minimally to organic nitrogen in total (approximately 2% of total organic nitrogen in aerosols), the individual amino acids contributing about 75% of amino nitrogen were indicative of biological organisms such as bacteria (10). Thus, we can conclude that, given sufficient sunlight, cloud transporting Saharan dust would be an accessible medium for proliferation of microorganisms by providing bioavailable iron, organic nitrogen and simple amino acids such as glycine, proline and valine, which are demonstrated in Saharan desert dust-enriched rain.
Although there have been some controversial studies, low atmospheric pressure, wind and temperature are often correlated with headache attacks (27–31). These studies have provided trivial explanations for the triggering mechanism of weather conditions. Saharan dust in atmospheric air has been a recently discovered subject with a significant impact on public health. Until now, its role as a headache trigger has not been investigated, and this is the first study to test the hypothesis that ‘Saharan dust in atmospheric air triggers migraine headache’.
Material and methods
The study was performed in Gazi University Neuropsychiatry Centre Neuroscience Laboratory after ethics committee approval had been obtained from Gazi University Animal Studies Ethical Committee. Male Wistar rats weighing 200–250 g were used. Animals were kept in a special compartment avoiding light from outside and exposed to 12 h daylight, 12 h dark with 18–21°C fixed temperature.
Preparation of dust
Saharan desert soil was brought by C.S. from around Tozeur city south of Tunisia, where there was a proper resource of soil with the potential of atmospheric transport to Turkey. The soil was sifted through a fine sieve to obtain a dust fraction < 200 μm. To test ‘the possible impact of organic composition of desert dust as a triggering factor of migraine headache’ hypothesis, part of the desert dust was treated with Co60 gamma irradiation for 2 days before the experiment to sterilize the dust by non-destructive methods avoiding alteration of clay mineralogy as described before by Saydam and Senyuva (23). Saharan dust pulse waves are variable in the regional atmosphere, being in the range of 0.5–326 μg/m3 (13), which is compatible with concentrations chosen for simulated atmospheric conditions.
Dust-containing experimental atmospheric conditions
The experiment was arranged to simulate the atmospheric conditions of dust in the cloud, so 100 mg to 200 g sieved dust was put into cylindrical measuring cups and distilled sterilized water was added to 2 l. This blend was mixed with air for 24 h and irradiated with a 500-W tungsten lamp to imitate solar radiation. Following the irradiation and mixing, air was turned off and the mixture was allowed to settle for 2 h. Approximately 300 ml of supernatant solution was transferred as mist through a sonic nebulizer at a rate of 100–150 ml/h into the atmosphere, where freely moving rats were incubated. Animals were kept under these stimulated atmospheric conditions for 2 h.
We attempted to quantify solid material suspended in the air through sonification of various dust-containing solutions, although there is no known system to measure the quantity of solid material suspended in 100% humidified air. For this purpose, a hole was opened and 200 ml/min air was sucked into an iced water bath to obtain diffusion of vapour into water. This mechanism was operated for at least 2 h and when water increased by 50% in the container, the diffusion process was stopped. This liquid was filtrated through weight-stabilized 0.45 μm pore size cellulose acetate paper and weighed. The experiment was first performed with distilled water and the results were taken as baseline. However, no significant weight difference was found between baseline and experimental conditions containing Saharan dust. That result indicated that experimental conditions did not bring an extra burden of suspended solid material, and simulated Saharan dust-laden atmospheric conditions were considered as containing particles probably < 0.45 μm.
Experimental groups
Experiments were performed under three different environmental conditions. The Saharan dust group (n = 21) was obtained by incubation of 50 mg to 100 g/l dust mixed with water kept under the light simulating solar radiation for 12 h. Co60-treated Saharan dust (sterilized) was prepared by irradiating desert dust (100 g/l) with Co60 for 2 days to kill live microorganisms and sterilize the dust as described by Saydam and Senyuva (23). Thereafter, Co60-treated Saharan dust was processed similar to Saharan dust (n = 9).
As a control, dust- and bacteria-free air (control) (n = 8) was generated by introducing air filtered through a water-based filter. The ambient air was pumped through 20 l distilled sterilized water through airstones. It was assumed that while the bubbles were rising towards the surface the vortex created within the bubbles would provide enough surface area and time for any particles to be transferred to water and leaving dust- and bacteria-free air behind. This atmosphere was assumed to be free of any ambient atmospheric particulates of any size fraction. Prior to experiments, all animals were kept in such created dust-free atmospheric conditions, then transferred to each experimental environment. The control group was kept in such an atmosphere till the end of incubation. During the course of Saharan dust irradiation, dust- and bacteria-free air was also used to stir the dust water mixture.
C-fos immunohistochemistry
Animals were anaesthetized with a lethal dosage of tiopenthal sodium after 2 h incubation in the simulated atmospheric conditions. They were sacrificed and perfused transcardially by heparinated saline followed by 4% 0.1 M paraformaldehyde solution. Brain and brainstem were prepared for c-fos immunohistochemistry as described before (32). Sections at obex level (obex ± 0.5 mm), lower than obex brainstem sections [between obex −0.5 mm and −3 mm, trigeminal nucleus caudalis (TNC)] and upper cervical sections (between obex −3 mm and −5 mm) were evaluated for c-fos immunoreactivity in TNC.
Nitrate + nitrite (NOx) assay
Blood samples were obtained to determine plasma nitrate + nitrite (NOx) levels (Griess method) as a marker of nitric oxide (NO) production. NOx levels were obtained from an enzyme-linked immunosorbent assay (ELISA) reader by vanadium chloride (VCl3)/Griess assay. First, 0.3 M 500 μl sodium hydroxide (NaOH) was mixed with 500 μl plasma. After incubation for 5 min at room temperature, 500 μl 10% zinc sulphate (ZnSO4) was added for deproteinization. This mixture was then centrifuged at 18 228 g for 5 min and supernatants were used for the assays. Nitrate standard solution was serially diluted. After loading the plate with samples (100 μl), addition of vanadium III chloride (VCl3) (100 μl) to each well was rapidly followed by addition of Griess reagents, sulphanilamide (SULF) (50 μl) and N-(1-naphtyl) ethylenediamide dihydrochloride (50 μl). After incubation (usually 30–45 min), samples were measured at 540 nm using an ELISA reader (33).
Microbiological analysis
The specimens for microbiological assays were obtained from dust-containing aerosols when Saharan desert dust (or sterilized Saharan desert dust with cobalt) was mixed with water and left for incubation under solar light. Samples for microbiological analysis were taken only from the 100 g/l concentration of Saharan dust group. Control specimens were also obtained from the same setting (without any dust content). Specimens were taken from those solutions twice, the first at 1 h and the second at 24 h (just before animals were exposed to it as an aeorosol). Microbiological specimens (100 ml) were taken and transferred to blood agar, eosin methylene blue (EMB) agar, Sabouraud dextrose agar and chocolate agar and incubated for 72 h before evaluation. Air specimens were taken via special air samplers designed for microbial sampling at a fixed suction rate for 1 min and cultured onto Sabouraud dextrose agar plates to detect fungal growth. Each specimen was examined for aerobic bacteria and fungi. Serial 10-fold dilutions were made, and 0.1-ml aliquots were plated on the following agar media: blood agar, chocolate agar, EMB agar and Sabouraud dextrose agar. Bacterial culture plates were incubated aerobically at 37 oC for 2 weeks. Sabouraud dextrose agar plates were incubated at room temperature for 4 weeks to detect fungal growth. Bacterial and fungal isolates were identified to the genus level by colony morphology and microscopic examination. Some could not be identified and were therefore expressed in terms of Gram-staining properties. The results were reported in terms of the colony-forming units (CFU) per ml of dust-contaminated water or CFU of air sampled from the system.
Statistics
Results were evaluated with
Results
Brain stem c-fos
No significant difference in behaviour was observed among experimental groups, although rats exposed to Saharan dust seemed to exhibit more face-grooming behaviour (34), which might be related to perception of pain and anxiety.
c-fos+ cells in brainstem sections including TNC (lamina I and II) in each experimental group are shown in Fig. 2. Brainstem sections between obex and C2 of freely moving rats in dust-free group contained few c-fos+ neurons, which were scattered in all laminae and not clustered in one particular area. Saharan dust groups consisted of four different dust concentrations: 50 mg/l, 500 mg/l, 10 g/kg and 100 g/l. When compared with the dust-free group, the number of c-fos+ neurons were higher in the Saharan dust group (Fig. 2) and they were concentrated in nociceptive laminae I and II (P = 0.0001). c-fos+ neurons were bilaterally symmetrical and their distribution was similar to cortical spreading depression (CSD)-induced c-fos cell distribution in TNC, and they were located prominently in ventral part where the ophthalmic branch of the trigeminal nerve makes synapses (data not shown).
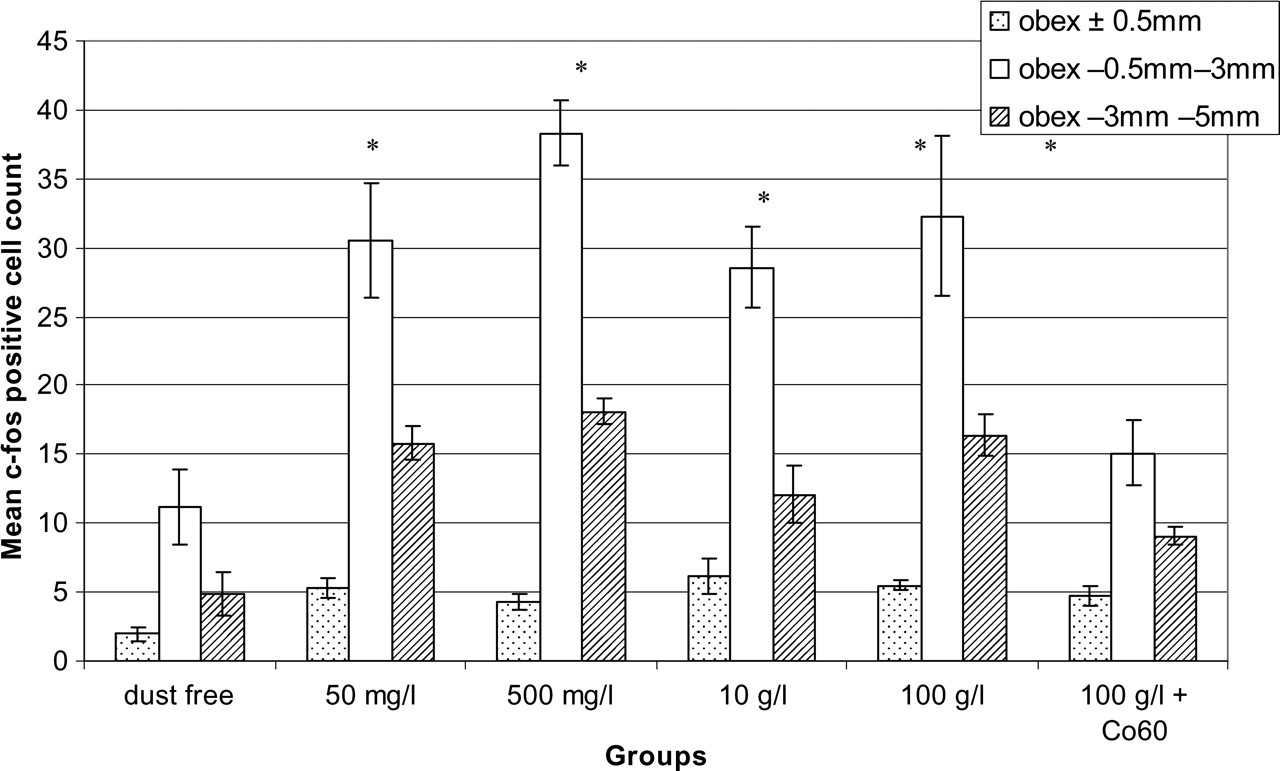
The comparison of mean c-fos+ cells in brainstem sections in experimental groups. c-fos expression in nociceptive superficial laminae (I and II) in trigeminal nucleus caudalis (TNC) (between obex −0.5 mm and −3 mm) and upper cervical sections (between obex −3 mm and −5 mm) is significantly increased in animals exposed to Saharan dust-laden atmospheric conditions. The number of c-fos+ neurons did not significantly change with different concentrations of Saharan dust. Sterilization of high concentration of Saharan dust with Co60 resulted in decreased c-fos expression in TNC, implying that the same dust without microorganisms did not activate nociceptive neurons (∗P < 0.05).
To eliminate the effect of microorganisms within Saharan dust, sterilization with Co60 was performed 24 h previously. This process was applied only to the highest concentration of Saharan dust (100 g/l). Number of c-fos+ neurons in TNC was reduced in the group that received Saharan dust sterilized with Co60.
When c-fos+ cell numbers of the Saharan dust group were compared with the Co60-treated Saharan dust or dust-free group, significant difference was found between TNC sections (P = 0.0001) and upper cervical sections (Fig. 2); however, c-fos activation around the obex level in the Saharan dust group did not show any significant difference from that of the Co60-treated Saharan dust or dust-free air group (P = 0.743, P = 0.676, respectively). c-fos cell numbers were not significantly different in obex, TNC and upper cervical sections among different concentrations of Saharan dust groups (P > 0.05).
Cerebral cortex c-fos
In freely moving animals, c-fos immunoreactivity was very low and found to be scattered, particularly in somatosensorial areas in the cerebral cortex. c-fos immunoreactivity did not differ significantly among Saharan dust- and dust-free groups. Furthermore, c-fos expression indicating CSD (dense and diffuse c-fos activation in all laminae of cerebral cortex) was not detected in any rat brain subjected to Saharan dust.
NOx results
The plasma levels of nitrate + nitrite (NOx) were significantly higher in the Saharan dust group (17.7 ± 11.71 μM/l) compared with the Co60-sterilized dust group (6.7 ± 3.62 μM/l, P = 0.047, Table 1).
Microorganisms detected (CFU/ml) at 1 and 24 h in simulated atmospheric conditions, plasma NOx level and c-fos+ cells in experimental groups
Microbiological analysis
In the Saharan dust group, 1250 CFU/ml and 20 000 CFU/ml microorganisms were detected at 1 h and 24 h, respectively (Table 1). In dust-free group, 110 CFU/ml and 2000 CFU/ml and in the Co60-sterilized group, 125 CFU/ml and 210 CFU/ml colonies were recognized at 1 h and 24 h, respectively (Table 1). The number of microorganisms in the Saharan dust group was significantly higher than in both the dust-free and Co60-sterilized groups. The names of various microorganisms found in Saharan dust samples are listed in Table 2.
Genera of bacteria and fungi found in Saharan dust samples

Discussion
The present study provides evidence for the first time that an unidentified factor existing in the atmosphere could activate the trigeminovascular system and trigger headache. It is very unfamiliar concept that Saharan desert dust transported in the atmosphere could reach many continents and countries around the globe, leading to epidemic health problems such as allergic reactions, asthma attacks and possibly headache (14, 15). Changes in weather conditions are among the well-known triggering factors for headache, particularly for migraine (27–31, 35). Microorganisms carried in the atmosphere along with Sahara desert dust have gained attention lately and been associated with some environmental health problems. Transport of Saharan dust in atmospheric air to some extent reminds us that other triggering factors may be transported in the same manner. By demonstrating the capability of a novel atmospheric factor in triggering headache the presented study provides a new concept.
The headache phase of migraine is characterized by activation of the trigeminovascular system, leading to neurogenic inflammation in the dura mater (36–38), although it is not known exactly how triggering factors activate the trigeminovascular system and initiate headache. The cells in Sp5C lamina I and II receive direct synaptic input from primary afferent fibres about transmission of pain in trigeminal receptive area. Expression of immediate early gene c-fos in second-order neurons in TNC occurs when the trigeminovascular system is activated (39, 40).
In the present study, c-fos immunoreactivity in TNC, a validated method for trigeminovascular system activation, was used (39). Saharan desert dust-containing air induced significant c-fos expression of brainstem TNC neurons. The noxious effect was further strengthened by the observation that the distribution of c-fos immunoreactivity is prominent in the nociceptive superficial lamina of TNC and upper cervical sections particularly in the topographic area where the ophthalmic branch of the trigeminal nerve ends. c-fos expression was symmetrical and there was no lateralizing finding. We also detected little other neuronal c-fos activation in other brainstem areas and in the cerebral cortex. Even in the dust-free group, which was not exposed to Saharan dust, very little c-fos expression was detected. This is not an unexpected finding, since animals were awake, going about daily activities (walking around the cage, eating, fighting with other rats) and subject to many stimuli.
Identifying the triggering factor, whether it is Saharan dust itself, accompanying microorganisms or other factors, is very important. According to our results, it is less likely that Saharan dust is the sole triggering factor, since the same Saharan dust following sterilization with Co60 did not activate the trigeminovascular system. Microbiological analysis revealed a significantly higher number of microorganisms in the Saharan dust group at both 1 h and 24 h of sampling. There was no statistical difference between the Co60-sterilized Saharan dust and the dust-free groups regarding microorganisms. Therefore, we suggest that microorganisms or by-products of microorganisms in the Sahara desert dust rather than dust itself could be related to the activation of the trigeminovascular system.
The aim of the light source used was to simulate the atmospheric environment and support microorganisms to multiply and create products such as oxalate and Fe2+, carboxyl radicals that can easily react with biological structures exposed to sunlight. Therefore, a rich environment of dust-associated microorganisms and their by-products was obtained. Saydam and Senyuva (23) have shown that Co60-sterilized Saharan dust has a significantly reduced capacity to produce Fe2+ and oxalate. Co60 gamma radiation source was especially preferred to obtain sterilization without harming clay mineral instead of many wet sterilization techniques. Soil samples irradiated in that way were not able to produce reduced iron. These results indicate that the presence of bacteria and fungi is the controlling step during the production of reduced iron in Saharan soil samples.
Recent data have further supported Saydam and Senyuva's findings and presented new evidence for the behaviour of desert soil in cloud (10, 23). Mace and colleagues collected rain and aerosol samples from Saharan air and measurements of total organic nitrogen and calcium, magnesium, sulphate and nitrate were found strongly correlated with atmospheric dust. They found that rain and aerosol organic nitrogen quantities were 13% and 17% of total organic nitrogen, respectively, and the ratio of free amino acids (glycine, arginine, proline and valine) contributing to total amino nitrogen was as high as 75% (10). The latter finding gains importance, as the existence of amino acids such as glycine, proline and valine is indicative of biological organisms such as bacteria in the medium (41, 42).
We further included particle size investigation in our study, although it was not possible to correlate the triggering effect directly to the presence of microorganisms. Studies performed with various calibrated filtration systems revealed that the air that a rat inhaled did not contain particles > 0.45 μm sonically. Microorganisms such as bacteria and fungi are significantly larger than 0.45 μm, therefore we assume that these microorganisms may not be directly involved in triggering pain. It must be kept in mind as a limitation of this method that the fraction of bacteria and fungi cannot be definitely determined by the weight-determining sonic system. The most difficult step is determining the presence of main contributors in the moist environment of particles. This situation suggests the need for new sampling methodologies. However, we still propose that particles < 0.45 μm and by-products of microorganisms generated during proliferation in the cloud-simulated conditions could be the trigger for headache.
Although the mechanism of trigeminovascular system activation by Sahara desert dust-laden weather conditions remains unknown, several possibilities are likely. Increased allergic and asthmatic attacks during Saharan dust streams in epidemiological studies (14, 15) support the notion that Saharan dust could act on meningeal cells, particularly mast cells, to activate trigeminal nerve fibres through inflammatory molecules. Increased plasma NOx levels in the Saharan dust group may suggest that Saharan dust-induced trigeminovascular activation could be related to NO generation. It has been shown that NO induces calcitonin gene-related peptide synthesis and secretion from trigeminal neurons (43). Increases in systemic NO could mediate generation of prodromal symptoms and headache as shown in the glyceryl trinitrate (GTN) model (44, 45). Administration of GTN, a NO donor, leads to transient systemic NO increase and induces nuclear factor-κβ-mediated inducible nitric oxide synthase expression in meningeal resident macrophages besides inducing cytokines such as interleukin (IL)-1β and IL-6. Dural mast cells are also activated by GTN (46, 47). GTN has been shown to activate central structures as shown by c-fos and cyclooxygenase-2 expression in TNC (48, 49). Therefore, it can be proposed that Saharan dust-containing air could trigger the trigeminovascular system through NO-mediated reactions, and the precise mechanisms involved in this pathway should be explored by further studies. Our data have also revealed that c-fos activation in the TNC is not due to the CSD, as shown by negative cerebral cortical c-fos staining. Saharan desert dust did not induce any CSD in the cerebral cortex of rats. The possibility that simulated conditions had an impact on subcortical and brainstem structures such as periaqueductal grey or rostral ventromedial medulla involved in modulation of nociceptive signals can not be disregarded.
Several limitations of the study warrant mention. First, our laboratory conditions may not have completely and accurately simulated the composition of Saharan dust in the atmosphere. Second, although we were able to show trigeminal nucleus activation by air-borne Saharan soil extracts, this does not necessarily mean that these would trigger migraine headaches in humans.
In the presented study, activation of the trigeminovascular system by Saharan dust and associated microorganisms carried by the air stream has been shown in animals for the first time. Further studies are needed to identify the mechanisms and molecules that mediate that nociceptive effect. Description of a potential headache-triggering effect of an atmospheric factor that remained unnoticed for a long time suggests that the desert dust content of any weather condition besides temperature, pressure, wind and humidity alterations should be investigated in relation to headache in patients. For example, the Chinook days could be accompanied by an increased amount of dust in the air, and the mechanisms possibly shared with Saharan dust might be applied to Chinook wind-triggered migraine headache (30, 35, 50). Clinical prospective studies on the potential triggering effect of Saharan dust-containing weather alterations on patients with migraine and other headaches could provide notification of patients for preventive medication on a daily or seasonal basis.
Footnotes
Acknowledgements
The study was supported by Gazi University (01/2003-19) and SPO State Planning Office (03-K-120 570-2). A part of this work has been presented in International Headache Society 2007 in Stockholm.