
Abstract
Sensitivity to light was investigated 5 and 8 h after consumption of an amino acid drink which contained L-tryptophan (balanced amino acid condition: 19 controls and 22 migraine sufferers) or which produced a short-term reduction in brain serotonin synthesis by omitting L-tryptophan (tryptophan depletion condition: 16 controls and 16 migraine sufferers). Migraine sufferers reported more intense nausea, headache, glare- and light-induced pain than controls. In addition, glare- and light-induced pain were greater in the tryptophan depletion condition than in the balanced amino acid condition, in both migraine sufferers and controls. Eight hours after the amino acid drink, after participants had completed tests of pain sensitivity and motion sickness provocation, tryptophan depletion augmented headache in migraine sufferers and aggravated nausea in migraine sufferers and controls. These findings suggest that a reduction in brain synthesis of serotonin intensifies photophobia and other migrainous symptoms and thus might contribute to the pathogenesis of migraine.
Introduction
Serotonin (5-HT) has long been implicated in the pathogenesis of migraine (1). Susceptibility to migraine is associated with chronically low concentrations of plasma serotonin (2) and heightened sensitivity to serotonin agonists (3), possibly due to a disturbance in the metabolism of serotonin (2). In addition, alterations in the sensitivity of serotonergic receptors or turnover of serotonin in the central nervous system (1, 4) could contribute to interictal abnormalities in sensory processing such as attenuated habituation of cortical evoked responses in migraineurs (5). During attacks of migraine, serotonin is released from platelets into blood plasma, possibly mirroring a disturbance in serotonin turnover in the central nervous system (6).
The 5-HT1A,2B/C agonist m-chlorophenylpiperazine provokes migrainous symptoms (7) and headache in migraine sufferers (8). The headache builds up several hours after plasma concentrations and cardiovascular effects peak, apparently during a period of relative reduction in serotonin activity following stimulation of serotonin receptors (8). Drugs such as fenfluramine and reserpine, that release and then deplete endogenous stores of serotonin, also induce headache within a few hours in susceptible people (3, 9, 10). Headache sometimes develops before blood serotonin concentrations decline (9, 10), suggesting that the headache is triggered by neural rather than systemic depletion of serotonin.
Rapid tryptophan depletion produces a short-term reduction in brain serotonin synthesis and has been employed in human studies to investigate the adverse effect of this reduction on mood disorders such as depression (11, 12). In rats, a low-tryptophan diet or tryptophan-free amino acid mixture decreases spontaneous and d-fenfluramine-induced release of serotonin and its metabolite5-hydroxyindoleacetic acid (5-HIAA) into the extracellular fluid of the hippocampus (13, 14). In humans, tryptophan depletion markedly diminishes serotonin synthesis, measured by positron emission tomographic scanning using α-11C-methyl-L-tryptophan as a tracer, both in cortical and subcortical regions of the brain (15). In addition, concentrations of 5-HIAA in the cerebrospinal fluid fall around 30% several hours after tryptophan depletion, indicating that central serotonin turnover declines substantially (16, 17).
Tryptophan depletion does not trigger full-blown attacks of migraine (11, 18), but may heighten susceptibility to migrainous symptoms (18). The effect of tryptophan depletion on susceptibility to motion sickness provoked by optokinetic stimulation was investigated recently in our laboratory (18). In general, vestibular symptoms such as nausea and dizziness were greater in migraine sufferers than in controls during optokinetic stimulation. In addition, tryptophan depletion increased susceptibility to these symptoms in controls, thus raising the possibility that a chronically low concentration of brain serotonin increases susceptibility to vestibular disturbances in migraine sufferers.
The aim of the present study was to determine whether tryptophan depletion would augment photophobia, one of the hallmarks of migraine. During the headache-free interval, bright light and high-contrast striped patterns induce greater discomfort in migraine sufferers than in controls (19–25). Photophobia increases in migraine sufferers during painful stimulation of the face (20, 26) and during migrainous attacks (19, 24, 25). The mechanism of photophobia is unclear, but could involve convergence of visual and trigeminal nociceptive activity in the thalamus or superior colliculus. In patients with ocular disease, irritation or inflammation of structures supplied by trigeminal nociceptive afferents (e.g. the iris, cornea or conjunctiva) contributes to photophobia (27). Photophobia also develops in patients with trigeminal neuralgia and after infection or lesions that involve pain-sensitive structures at the base of the brain (e.g. meningitis, subarachnoid haemorrhage or tumours around the optic chiasm) (28–30). Spatial summation of visual input in the central nervous system contributes to photophobia—the brightness (radiance) of a visual stimulus required to elicit visual discomfort is less for large than for small areas of retinal stimulation (31) and is lower for binocular than monocular stimulation (24, 32). Serotonergic neurons in the dorsal raphe nucleus send axon collaterals to multiple cortical and subcortical trigeminal somatosensory targets (33). A large serotonergic projection from the dorsal raphe nucleus also terminates in the superior colliculus (34), a potential site of visual and trigeminal nociceptive convergence (35, 36). As serotonin generally exerts an inhibitory influence on discharge of visual and nociceptive neurons (34, 37), depletion of serotonin might enhance photophobia.
In a previous study in our laboratory (38), photophobia persisted during a 30-min recovery period after optokinetic stimulation in migraine sufferers but decreased in controls. Hence, photophobia was investigated before and after optokinetic stimulation in the present study, to determine the combined effect of tryptophan depletion, residual nausea and headache on photophobia. Lebensohn and Bellows (27) distinguished between painless glare or dazzle—possibly due to thalamic dysfunction (39)—and pain induced by bright light in patients with painful ocular or intracranial disease. Thus, both glare and light-induced pain were examined in the present study. It was hypothesized that photophobia would be greater in the tryptophan depletion condition than in a balanced amino acid condition and that this would be most evident in the presence of residual symptoms provoked by optokinetic stimulation.
Methods
Subjects
The migraine sample consisted of 32 women and six men aged between 18 and 57 years (mean ± SD 35.1 ± 13.8 years) who met diagnostic criteria (40) for migraine with aura (n = 8) or migraine without aura (n = 30) and who suffered from one or more headache episodes each month. The controls consisted of 26 healthy women and nine men aged between 19 and 59 years (mean age 36.7 ± 14.1 years) without a history of migraine who experienced six or fewer mild headaches per year. The facilitatory effects of tryptophan depletion in these participants on symptoms provoked by optokinetic stimulation have been reported previously (18). Migraine sufferers were tested when they had been free from headache for 9.3 ± 7.5 days and women were tested between menstrual periods. Apart from the oral contraceptive, none of the participants took preventative medicine for migraine or for any other condition.
Each participant provided informed consent for the procedures, which were approved by the Murdoch University Human Research Ethics Committee.
Procedures
The procedures were carried out in a temperature-controlled room, maintained at 23 ± 1°C. Participants had a light protein-free breakfast without caffeine and reported to the laboratory around 08.00 h. The synthesis of serotonin depends on dietary intake of tryptophan, the amino acid precursor of serotonin. Ingestion of a tryptophan-free mixture of amino acids provokes a large and rapid fall in plasma tryptophan by stimulating protein synthesis in the liver. In addition, competition between tryptophan and other large amino acids for transport across the blood–brain barrier restricts the entry of tryptophan into the brain, thereby decreasing the synthesis of serotonin in brainstem projection neurons (the main source of serotonin in the central nervous system) (12). Thus, to accomplish rapid tryptophan depletion, participants consumed capsules and a drink containing 5.5 g
Sensitivity to light was investigated approximately 5 h and 8 h after the amino acids were ingested. During the 3-h interval between the two tests of light sensitivity, participants completed a range of neurophysiological and sensory tests that aimed to investigate the effect of tryptophan depletion on symptoms and signs of migraine. In particular, blink reflexes to electrical stimulation of the supraorbital nerve were recorded before and during limb and cranial pain induced by ice (unpublished observations), followed by optokinetic stimulation that involved a visual illusion of movement created by watching black and white stripes move past for 15 min or until vomiting was imminent (18). The second test of light sensitivity began approximately 20 min after termination of optokinetic stimulation, when some participants were still experiencing residual symptoms of motion sickness. The intensity of nausea and headache was rated before and after each test of light sensitivity on scales where 0 corresponded to 'no sensation at all' and 10 to 'extreme'.
During each test of light sensitivity, the participant sat approximately 80 cm in front of a 50-W halogen globe located behind a translucent plastic screen. Electrical power to the halogen globe was controlled by a computer to produce three levels of screen luminance (90 cd/m2, 4100 cd/m2 and 19 800 cd/m2 measured with a Photo Research Spectra spotmeter, Model PR1500; Chatsworth, CA, USA). Twenty seconds after light onset, the participant rated glare- and light-induced pain between 0 and 10, where 0 corresponded to 'not at all' and 10 to 'extreme'. The participant then waited for 30 s in darkness before the next trial began. Each level of light intensity was presented three times in random order.
Data reduction and statistical approach
Mean glare and pain ratings for each level of light intensity were investigated separately in 2 × 2 × 2 × 3 [Group (migraine, control) × Condition (tryptophan depletion, balanced amino acids) × Test (first, second) × Light Intensity (dim, moderately bright, bright)] analyses of variance. Similarly, nausea and headache ratings were investigated separately in Group × Condition × Test × Pre-Post (before vs. after each test) analyses of variance. The multivariate solution was used for factors with more than two levels because the multivariate approach does not require assumptions of sphericity of the variance–covariance matrix to be met (41). The criterion of statistical significance was P < 0.05 and only significant main effects and interactions are reported.
Results
Ratings of glare and light-induced pain
As shown in Fig. 1, glare ratings were greatest for bright light [main effect for Light Intensity: F(2,68) = 365.9, P < 0.001] and increased from before to after optokinetic stimulation [main effect for Test: F(1,69) = 17.4, P < 0.001]. Glare was greater in migraine sufferers than in controls both before and after optokinetic stimulation [main effect for Group: F(1,69) = 4.03, P < 0.05] and was greater throughout the experiment for participants in the tryptophan depletion condition than in the balanced amino acid condition [main effect for Condition: F(1,69) = 6.76, P < 0.05].
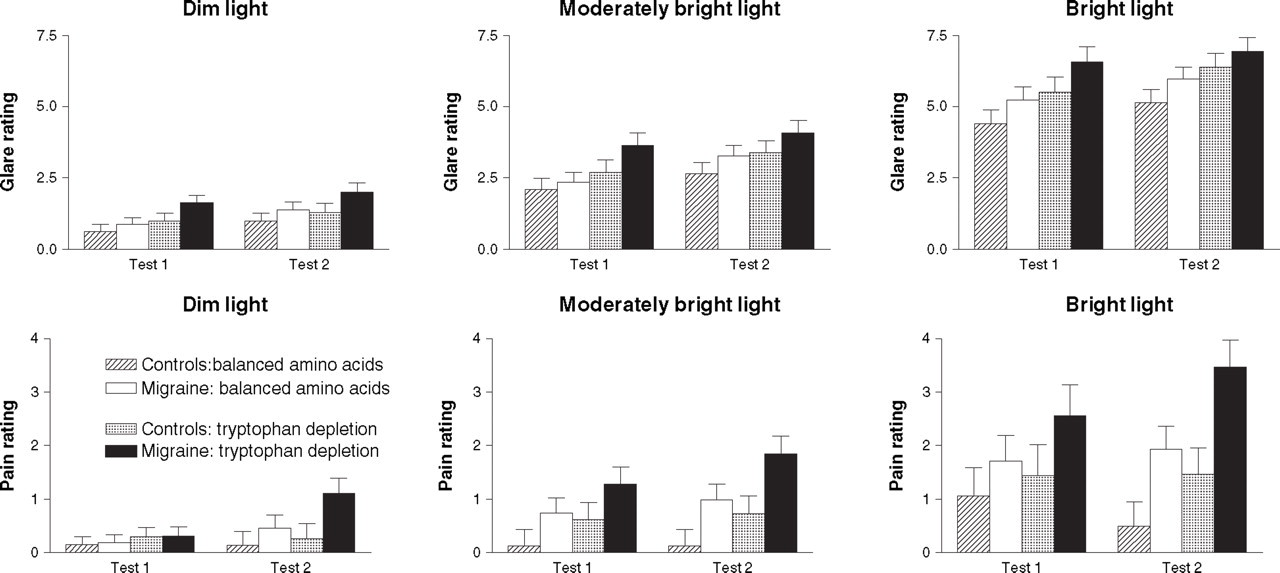
Ratings of glare and light-induced pain (±SEM) in migraine sufferers and controls in the tryptophan depletion and balanced amino acid conditions, during exposure to dim light, moderately bright light, and bright light before (test 1) and after optokinetic stimulation (test 2).
Ratings of light-induced pain also were greatest for bright light [main effect for Light Intensity: F(2,68) = 27.3, P < 0.001] (Fig. 1). Light-induced pain was greater in migraine sufferers than in controls both before and after optokinetic stimulation [main effect for Group: F(1,69) = 7.48, P < 0.01], particularly at the brighter intensities of light [Group–Light Intensity interaction: F(2,68) = 3.69, P < 0.05] (Fig. 1). Light-induced pain increased after optokinetic stimulation in migraine sufferers but not in controls [Group–Test interaction, F(1,69) = 6.89, P < 0.05] and was greater throughout the experiment for participants in the tryptophan depletion condition than in the balanced amino acid condition [main effect for Condition: F(1,69) = 4.19, P < 0.05].
Nausea and headache before and after each test of light sensitivity
Nausea was greater in migraine sufferers than in controls throughout the experiment [main effect for Group, F(1,69) = 11.9, P < 0.001] Fig. 2). Nausea remained relatively stable in migraine sufferers and controls in the balanced amino acid condition, but increased after optokinetic stimulation for participants in the tryptophan depletion condition [main effect for Test, F(1,69) = 11.6, P < 0.001; Condition–Test interaction, F(1,69) = 5.01, P < 0.05]. Nausea did not increase during the tests of light sensitivity (neither the Pre–Post main effect nor interactions that included this factor achieved statistical significance).

Nausea ratings (±SEM) in migraine sufferers and controls in the tryptophan depletion and balanced amino acid conditions, before (□) and after each test of light sensitivity (▪). Nausea increased during the interval between the first and second test of light sensitivity for migraine sufferers and controls in the tryptophan depletion condition (∗P < 0.05).
In general, headache ratings were low throughout the experiment and the headaches did not meet diagnostic criteria for migraine. Nevertheless, headache was greater in migraine sufferers than in controls [main effect for Group, F(1,69) = 25.9, P < 0.001] and increased during the tests of light sensitivity in migraine sufferers but not in controls [Group × Pre–Post interaction, F(1,69) = 10.5, P < 0.01] (Fig. 3). In addition, headache increased after optokinetic stimulation [main effect for Test, F(1,69) = 4.62, P < 0.05], notably in migraine sufferers in the tryptophan depletion condition [Group × Condition–Test interaction, F(1,69) = 5.29, P < 0.05] (Fig. 3).

Headache ratings (±SEM) in migraine sufferers and controls in the tryptophan depletion and balanced amino acid conditions, before (□) and after each test of light sensitivity (▪). Headache increased during the interval between the first and second test of light sensitivity for migraine sufferers in the tryptophan depletion condition (∗P < 0.05).
Discussion
Ratings of glare- and light-induced pain were greater in the tryptophan depletion condition than in the balanced amino acid condition, in both migraine sufferers and headache-free controls. In addition, tryptophan depletion augmented headache in migraine sufferers and enhanced nausea in migraine sufferers and controls after they had completed tests of pain sensitivity and motion sickness provocation. As tryptophan depletion lowers levels of serotonin in the brain (13–17) but not in the systemic circulation (18), these findings suggest that a reduction in brain synthesis of serotonin aggravates symptoms of migraine.
Irrespective of tryptophan depletion, migraine sufferers reported more intense nausea, headache, glare and light-induced pain than participants who rarely or never suffered from headache. These findings concur with many previous reports (18–25) and indicate that symptoms of migraine develop readily in migraine sufferers during the headache-free interval. Whether this is a cause or effect of the migraine predisposition is uncertain. However, it seems reasonable to speculate that interictal hyperexcitability of the trigeminal system in migraine sufferers (42–44) increases vulnerability to headache and possibly also to photophobia (20, 26).
Sensitivity to light during attacks of migraine forms part of a global sensitivity to noise, odours, taste and touch. This implies a common central cause of sensory hypersensitivity (e.g. disruption to inhibitory serotonergic modulation of sensory input) (45). In the present study, ratings of glare and light-induced pain were greater in the tryptophan depletion condition than in the balanced amino acid condition. One potential explanation for this finding is that serotonergic neurons with multiple projections (e.g. from the dorsal raphe nucleus to the thalamus and superior colliculus) (33, 34) inhibit both the painless and painful components of photophobia.
In another part of this study, tryptophan depletion enhanced symptoms of motion sickness (nausea, dizziness and a sense of spinning) in control participants to levels approaching those of migraine sufferers (18). To determine whether residual symptoms of motion sickness would augment effects of tryptophan depletion on photophobia in the present study, the second test of light sensitivity began approximately 20 min after termination of the optokinetic stimulus that induced motion sickness. Nausea and headache were greater after than before optokinetic stimulation for migraine sufferers in the tryptophan depletion condition, but remained unchanged in the balanced amino acid condition. Similarly, nausea was greater after than before optokinetic stimulation for controls in the tryptophan depletion condition. As nausea was minimal in controls before optokinetic stimulation (mean rating 0.2 ± 0.1 on a 0–10 scale) (18), tryptophan depletion apparently inhibited the resolution of motion sickness. Photophobia increased after optokinetic stimulation—glare increased in migraine sufferers and controls and light-induced pain increased in migraine sufferers. In addition, both in migraine sufferers and controls, glare- and light-induced pain were greater during the tests of light sensitivity in the tryptophan depletion condition than in the balanced amino acid condition. Taken together, the findings suggest that the migraine predisposition, tryptophan depletion and residual symptoms of motion sickness augment photophobia. Clearly, these effects are not entirely independent of each other, because both the migraine predisposition and tryptophan depletion augmented residual motion sickness.
In previous studies in our laboratory, head pain was found to aggravate nausea and photophobia in migraine sufferers (20, 26, 46); conversely, nausea aggravated headache and scalp tenderness (38, 47). In addition, visual stimulation that provoked photophobia in migraine sufferers also heightened scalp tenderness (20, 48). Thus, symptoms of migraine appear to build upon each other during the headache-free interval, perhaps in a vicious circle. Although a reduction in brain serotonin synthesis might contribute to this synergistic process, some other factor must also be involved because tryptophan-depleted controls remained headache free and photophobia did not increase over the course of the present study, despite an increase in nausea. In contrast, headache increased during visual stimulation in migraine sufferers and photophobia increased in line with increases in headache as the experiment progressed. The link between headache and photophobia in migraine sufferers is consistent with many other observations that implicate trigeminal nociceptive involvement in photophobia (27, 30).
One limitation of the present study is that the sample size was too small to compare effects of tryptophan depletion in migraine sufferers with and without aura. The effect of serotonin depletion with para-chlorophenylalanine, a tryptophan hydroxylase inhibitor, on spreading depression of neural activity in the cerebral cortex (a model of the migraine aura) was investigated recently in rats (49). Serotonin depletion increased the number of waves of spreading depression induced by topical application of potassium chloride on the parietal cortex. Moreover, expression of c-fos-immunoreactivity in neurons in the trigeminal nucleus caudalis was greater ipsilateral than contralateral to the cortical spreading depression, and was greater bilaterally in serotonin-depleted rats than in controls. These findings suggest that cortical application of potassium chloride evokes activity in trigeminal nociceptive neurons and that serotonin depletion facilitates this activity. Thus, it would be interesting to determine whether tryptophan depletion alters cortical activity (indexed, for example, by cortical evoked responses) and whether this response differs between migraine sufferers with and without aura.
In conclusion, the present findings suggest that a reduction in brain synthesis of serotonin intensifies nausea, headache and photophobia. Hence, correcting this deficiency could prevent these symptoms from escalating before and during migrainous attacks and, in the longer term, reduce vulnerability to migraine. Tryptophan depletion could be a useful tool for investigating the effect of new treatments that target central serotonin turnover or that aim to block symptoms of migraine.
Footnotes
Acknowledgements
This study was supported by grants from the National Health and Medical Research Council of Australia and the UK Migraine Trust. The author thanks Anna Granston, Kath McLean and Kaye Anderson for their expert research assistance. Plasma tryprophan was assayed at the Australian Proteome Analysis Facility under the direction of Dr Bernie McInerney.