
Abstract
The effect of postmenopausal estrogen replacement therapy (ERT) on the risk or severity of cerebrovascular disorders is as yet unclear, and the evidence for flow preservation being a mechanism of estrogen neuroprotection remains elusive. The authors examined whether estrogen-mediated flow-preserving neuroprotective mechanisms, if any, may involve its angiogenic action. This study was conducted using middle-aged (44 weeks) female rats because of the importance of aging in cerebrovascular disease in women. Middle-aged female rats were subjected to sham operation, ovariectomy, or ovariectomy with ERT. The anatomic cerebral capillary morphology showed a significant reduction in the total capillary density in the frontal cortex after ovariectomy. This was associated with marked decreases in protein and gene expression of vascular endothelial growth factor and its angiogenic receptors in cerebral vessels, as demonstrated by immunohistochemistry and in situ hybridization. The expression levels of both estrogen receptor (ER) subtypes, ERα and ERβ, in cerebral vessels were significantly reduced after ovariectomy, but ERβ was more dramatically downregulated as assessed by the ERβ/ERα ratio. These ovariectomy-induced changes were completely prevented by ERT. Vascular endothelial growth factor appears to be a critical regulatory molecule for physiologic cerebral angiogenesis in middle-aged female rats and may play an important role in the flow-preserving neuroprotective action of estrogen through its angiogenic and antiapoptotic properties.
Keywords
A growing body of evidence indicates that the risk of cerebrovascular events in women rises after menopause (Wenger et al., 1993), but the benefit of postmenopausal estrogen replacement therapy (ERT) for cerebrovascular disorders such as stroke is not clear. Some clinical studies do not support the benefit of ERT for cerebrovascular disease (Viscoli et al., 2001; Writing Group for the Women's Health Initiative Investigators, 2002). Estrogen replacement therapy in hypoestrogenic postmenopausal women, however, has been shown to reduce the risk and severity of neurodegenerative decline associated with cerebrovascular disorders (Paganini-Hill, 1995; Schmidt et al., 1996). The protective role of estrogen in brain function has been experimentally verified by using animal models of cerebral ischemia, in which estrogen attenuates cerebrovascular injury (Simpkins et al., 1997; Alkayed et al., 1998; Dubal et al., 1998). Less well understood, however, are the potential mechanisms by which estrogen protects against cerebrovascular events. Estrogen is a pleiotropic hormone that acts beyond the scope of its reproductive function. One of estrogen's most important actions may be as an angiogenic (Losordo and Isner, 2001), in which capacity it could exert an important influence in regulating cerebral microvascular development. Because cerebral microvessels undertake a considerable portion of the total resistance to cerebral blood flow and are important in the coupling of brain metabolism to regional cerebral blood flow (Joo, 1985), the angiogenic action of estrogen could contribute to salvage of neural tissue during cerebral ischemic episodes. Therefore, we examined the influences of in vivo estrogen manipulation on the capillary network of cerebral tissue and cerebrovascular expression of molecules that can be considered to be essential for angiogenesis. Because estrogen regulates angiogenesis through receptor-mediated mechanisms (Losordo and Isner, 2001), we also investigated cerebrovascular expression of the two estrogen receptor (ER) subtypes, ERα and ERβ, in animals of differing hormonal status. Cerebrovascular disorders occur more frequently in older postmenopausal than in young women (Wenger et al., 1993). We thus used middle-aged female rats as an experimental model, which would have important implications for future therapeutic strategies in the setting of cerebrovascular diseases in elderly women.
MATERIALS AND METHODS
Animal models
All experimental procedures were approved by the Hokkaido University School of Medicine Animal Care and Use Committee.
Female Wistar rats, 40 weeks of age, weighing 200 to 230 g, were randomly assigned to two groups. One group of rats was ovariectomized by making a small incision in the lower abdomen and removing both ovaries while the animal was under anesthesia with ketamine (Liu et al., 2001); the other group received only laparotomy and served as intact females. Some ovariectomized rats were given 17β-estradiol (3 μg/d) subcutaneously through an osmotic pump (Model 2ML4; DURECT Corporation, Cupertino, CA, U.S.A.) that was implanted in the back (Liu et al., 2001). The flow rate and concentration of 17β-estradiol in the pump were 2.54 μL/h and 0.17 mg/mL, respectively. Finally, we prepared three groups of 20 animals each. Rats were killed 4 weeks after surgery. On the day of the experiment, a blood sample was collected for the analysis of plasma 17β-estradiol. Plasma 17β-estradiol concentrations were measured by radioimmunoassay (Double Antibody Estradiol; Iatron Laboratories, Tokyo, Japan). Ovariectomized female rats had significantly lower plasma concentrations of 17β-estradiol than intact females (2.3 ± 1.1 versus 12.6 ± 9.3 pg/mL; n = 20 for each, P < 0.05). The low levels of plasma 17β-estradiol in ovariectomized females were completely improved by ERT (16.6 ± 21.0 pg/mL; n = 20, P < 0.01).
After rats were killed by exsanguination under anesthesia with gaseous diethyl ether, the brain was harvested. The brain was immediately frozen in liquid nitrogen and preserved at −80°C. For paraffin sections, the brain was postfixed in 4% paraformaldehyde overnight and processed routinely for paraffin embedding.
Capillary morphology
Eight-micron-thick serial frozen coronal sections of frontal cortex were stained with lectin Griffonia simplicifolia (GSA-B4; Sigma Chemical, St. Louis, MO, U.S.A.). The sections were fixed with acetone, air dried, and placed in phosphate-buffered saline (PBS). After being treated with 3% H2O2 in methanol and washed in PBS, the sections were incubated with GSA-B4 (1:100 dilution in PBS) overnight at 4°C, followed by reaction with streptavidin conjugated to peroxidase (Nichirei Corporation, Tokyo, Japan) and thorough rinse in PBS. For visualization, diaminobenzidine/H2O2 as chromogen was used. To enhance the diaminobenzidine reaction, the sections were rinsed with 0.05 mol/L NaHCO3 (pH 9.6) and then incubated in Diaminobenzidine Enhancing Solution (Vector Laboratories, Burlingame, CA, U.S.A.). Vascular endothelium was stained by the lectin with capillaries appearing as black or dark brown dots. Sections were examined with an Olympus microscope, and counts of capillaries were made in cross sections in 30 fields (117,617 μm2/field) per sample at a final magnification of x400. Any microvessel (defined as any vessel having an internal diameter <100 μm) that apparently had no lumen was considered as a single capillary.
Capillary density was also assessed light-microscopically on 5-μm-thick deparaffinized tissue sections that were immunostained by anti—von Willebrand factor (FVIII) antibody (Dako, Hamburg, Germany). The antibody was made visible by a secondary exposure of the sections to Cy3-conjugated AffiniPure donkey anti—rabbit immunoglobulin G (IgG; Jackson Immunoresearch Laboratories, West Grove, PA, U.S.A.).
Preliminarily, double staining of frozen tissue sections was carried out to discriminate arteriolar and venular capillaries. The double-staining method shows that arteriolar capillaries, whose endothelial cells contain alkaline phosphatase, are stained blue, whereas venular capillaries containing dipeptidylpeptidase IV are stained red (Jesmin et al., 2002). We found very few red-stained cerebral capillaries, suggesting that cerebral capillaries detected by the GS4 lectin method or the method using fluorescent staining for FVIII appear to be largely of the arteriolar type.
Immunohistochemistry and immunofluorescent labeling
Immunohistochemical studies were performed with the following commercially available antibodies: anti—human vascular endothelial growth factor (VEGF) polyclonal antibody (Immunobiological Laboratories, Fujioka, Japan), anti—human fetal liver kinase 1 (kinase domain region [KDR]) polyclonal antibody (Santa Cruz Biotechnology, Santa Cruz, CA, U.S.A.), anti—human fms-like tyrosine kinase 1 (Flt-1) polyclonal antibody (Santa Cruz Biotechnology), anti—human endothelial nitric oxide (NO) synthase (eNOS) polyclonal antibody (Affinity BioReagents, Golden, CO, U.S.A.), anti—human Bcl-2 polyclonal antibody (Santa Cruz Biotechnology), anti—mouse Bax polyclonal antibody (Santa Cruz Biotechnology), and anti—mouse phosphorylated Bad polyclonal antibody (Santa Cruz Biotechnology).
Eight- to 10-μm-thick frozen cryostat serial coronal sections of frontal cortex were fixed in acetone and air dried. Endogenous peroxidase activity was quenched by incubation in 3% hydrogen peroxide for 15 minutes. After being blocked y normal goat serum to prevent nonspecific staining by the secondary antibody, the sections were incubated with primary antibodies overnight at 4°C, followed by exposure to a suitable secondary antibody coupled to horseradish peroxidase. Immunostains were visualized by light microscopy with diaminobenzidine. The specificity of the immunoreaction was evaluated in comparison with the negative control specimen in which nonimmune IgG was used instead of the primary antibodies. Quantification of immunoreactivity by pixel intensity was analyzed using an image-analyzing software (Microcomputer Imaging Device; Imaging Research, St. Catharines, Ontario, Canada). This quantification has been well established in previous reports (Zaman et al., 2001; Jesmin et al., 2002).
After overnight incubation with each primary antibody as previously mentioned, the sections were exposed to the fluorescence secondary antibody, Cy3-conjugated AffiniPure anti—rabbit IgG or fluorescein-conjugated AffiniPure goat anti—rabbit, anti—goat, or anti—mouse IgG (Jackson Immunoresearch Laboratories), for 2 hours according to the manufacturer's instructions. The samples processed without primary antibodies served as negative controls. The coverslips were mounted with Immunon (Thermo Shandon, Pittsburgh, PA, U.S.A.). Immunofluorescent images were observed under Laser Scanning Confocal Imaging System (MRC-1024; Bio-Rad, Hemel Hempstead, U.K.). The intensity of immunofluorescent staining expressed as arbitrary unit was analyzed by free software, NIH Image, produced by Wayne Rasband (National Institutes of Health, Bethesda, MD, U.S.A.). The results obtained from immunofluorescent staining were essentially consistent with those from immunohistochemistry assessed by pixel intensity. Accordingly, the quantitative analysis shown herein is obtained from immunohistochemistry.
Immunohistochemical determination of estrogen receptors
Tissue specimens were fixed in 4% buffered formalin solution, dehydrated, embedded in paraffin, and then sliced into 5-μm-thick serial coronal sections. The sections were deparaffinized and treated for 20 minutes with citrate buffer (10 mmol/L citric acid, pH 6.0) in a microwave oven (750 W) before immunostaining. To prevent nonspecific staining by the secondary antibody, the sections were blocked by nonimmune serum (1% bovine albumin in Tris) for 30 minutes at room temperature. In preliminary experiments, the dilutions of the primary antibodies were titrated in each preparation to achieve optimal nuclear staining with minimal unspecific staining. For identification of ERα and ERβ, we used anti—human ERα monoclonal antibody (Novocastra Laboratories, Newcastle upon Tyne, U.K.), and anti—human ERβ polyclonal antibody (Affinity BioReagents), respectively. For us, these antibodies produced the most prominent robust signal with immunofluorescence. Controls included omission of the primary antibodies, omission of the secondary antibody, adsorption of the primary antibodies with their respective antigen, and cross-adsorption controls. For the latter, the ERα antiserum was adsorbed with the ERβ peptide and then was applied to tissue sections: the ERα immunostaining appeared normal. Similarly, adsorbing ERβ antiserum with the ERα protein did not diminish immunostaining for ERβ. The ratio of ERβ/ERα was calculated by dividing the mean level of ERβ by that of ERα in the same cerebral vessel, as determined by immunohistochemistry.
For double-label immunofluorescent staining of ERα and ERβ, the sections were incubated with ERα mouse monoclonal antibody, followed by Cy3-conjugated anti—mouse IgG, and then incubated with ERβ rabbit polyclonal antibody, followed by fluorescein-conjugated AffiniPure goat anti—rabbit IgG.
In situ hybridization
Twenty-micron-thick serial frozen coronal sections of frontal cortex were prepared and mounted on glass slides precoated with 3-amino-propyltriethoxysilane. The sections were then fixed by 4% paraformaldehyde with 0.25% acetic anhydride in 0.1 mol/L triethanolamine-HCl (pH 8.0). The hybridization procedure was performed as previously described (Mowa and Iwanaga, 2000). The probes were complementary to nucleotide residues 61 to 106 of VEGF complementary DNA (cDNA; accession number AF239170), 541 to 585 of KDR cDNA (accession number U93306), 961 to 1016 of Flt-1 cDNA (accession number D28498), 841 to 885 of eNOS cDNA (accession number X76309), 301 to 346 of ERα cDNA (accession number Y00102), and 45 to 90 of ERβ cDNA (accession number U57439). The oligonucleotides were labeled with 35 S-dATP using terminal deoxyribonucleotidyl transferase. The radiolabeled probes were hybridized to the tissue in a prehybridization buffer for 10 hours at 42°C. The sections were either exposed to Hyperfilm-βmax (Amersham, Little Chalfont, Buckinghamshire, U.K.) for 4 weeks or dipped in Kodak NTB2 nuclear track emulsion and exposed for 4 to 8 weeks. The specificity of in situ hybridization was confirmed by the disappearance of signals when excessive doses of the corresponding nonlabeled (35 S-dATP) antisense oligonucleotides (cold) were added to the labeled antisense oligonucleotides (hot) hybridization fluid. The messenger RNA (mRNA) grains per blood vessel were quantified using an image-analyzing software (Microcomputer Imaging Device), as previously reported (Jesmin et al., 2002).
Statistical analysis
Data are shown as mean ± SD, and the sample number equals the number of animals in each group. Data were analyzed using the Stat View II program (Abacus Concepts, Berkeley, CA, U.S.A.). Means were compared by analysis of variance, followed by the Fisher protected least significance t-test for multiple comparisons. Morphologic data were analyzed with nonparametric statistics (Kruskal-Wallis for analysis of variance designs). Differences were considered significant at P < 0.05.
RESULTS
The capillary morphology obtained from the GS4 lectin method shows that ovariectomy resulted in a remarkable reduction in capillaries in frontal cortex of middle-aged female rats (Fig. 1A). The capillary density determined by the GS4 lectin method showed a 31% reduction after ovariectomy (Fig. 1B). Estrogen replacement therapy for ovariectomized rats significantly improved the capillary density to the intact level (Fig. 1B). The method using fluorescent staining for FVIII (Fig. 1C) gave results comparable to those of the GS4 lectin method.
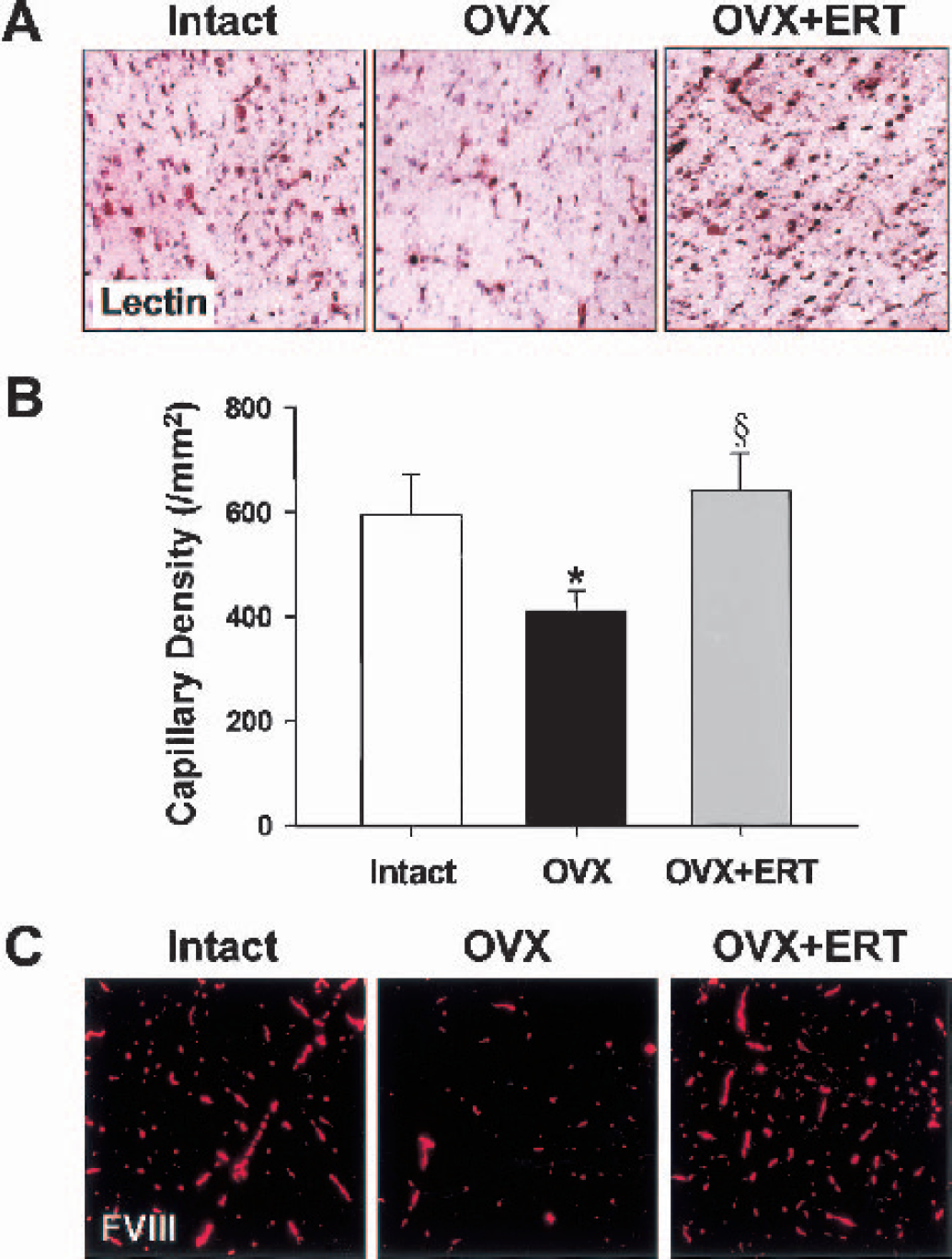
Evaluations of morphologically existing capillaries in coronal sections of frontal cortex from middle-aged female rats that were intact, ovariectomized (OVX), and OVX with estrogen replacement therapy (ERT).
Immunofluorescent staining for VEGF and its angiogenic receptor KDR showed that they were abundantly expressed in microvessels in coronal sections of frontal cortex from intact middle-aged females (Fig. 2A). Both VEGF and KDR were weakly stained after ovariectomy, and the reduced expression levels were completely reversed by ERT (Fig. 2A). As quantitation of immunoreactivity was assessed by pixel intensity, ovariectomy decreased the expression levels of VEGF and KDR to 43% and 44% of the respective intact value (Figs. 3A and 3B). In situ hybridization studies showed that VEGF and KDR mRNAs were also evidently diminished in microvessels in coronal sections of frontal cortex from ovariectomized rats compared with the findings in those from intact rats (Fig. 2B). When the numbers of mRNA grains per cerebral vessel were calculated, VEGF and KDR mRNAs were decreased to 49% and 45%, respectively, of intact animals by ovariectomy (Figs. 3C and 3D). Estrogen replacement therapy significantly increased the expressions of their mRNAs nearly to those of intact animals (Figs. 2 and 3). The expression of another VEGF receptor, Flt-1, which is devoid of angiogenic activities (Ferrara, 2001), was marginally affected by ovariectomy; when quantitation of immunoreactivity was assessed by pixel intensity, the values were 2.97 ± 0.08 for intact rats and 2.89 ± 0.09 for ovariectomized rats (n = 20 for each).
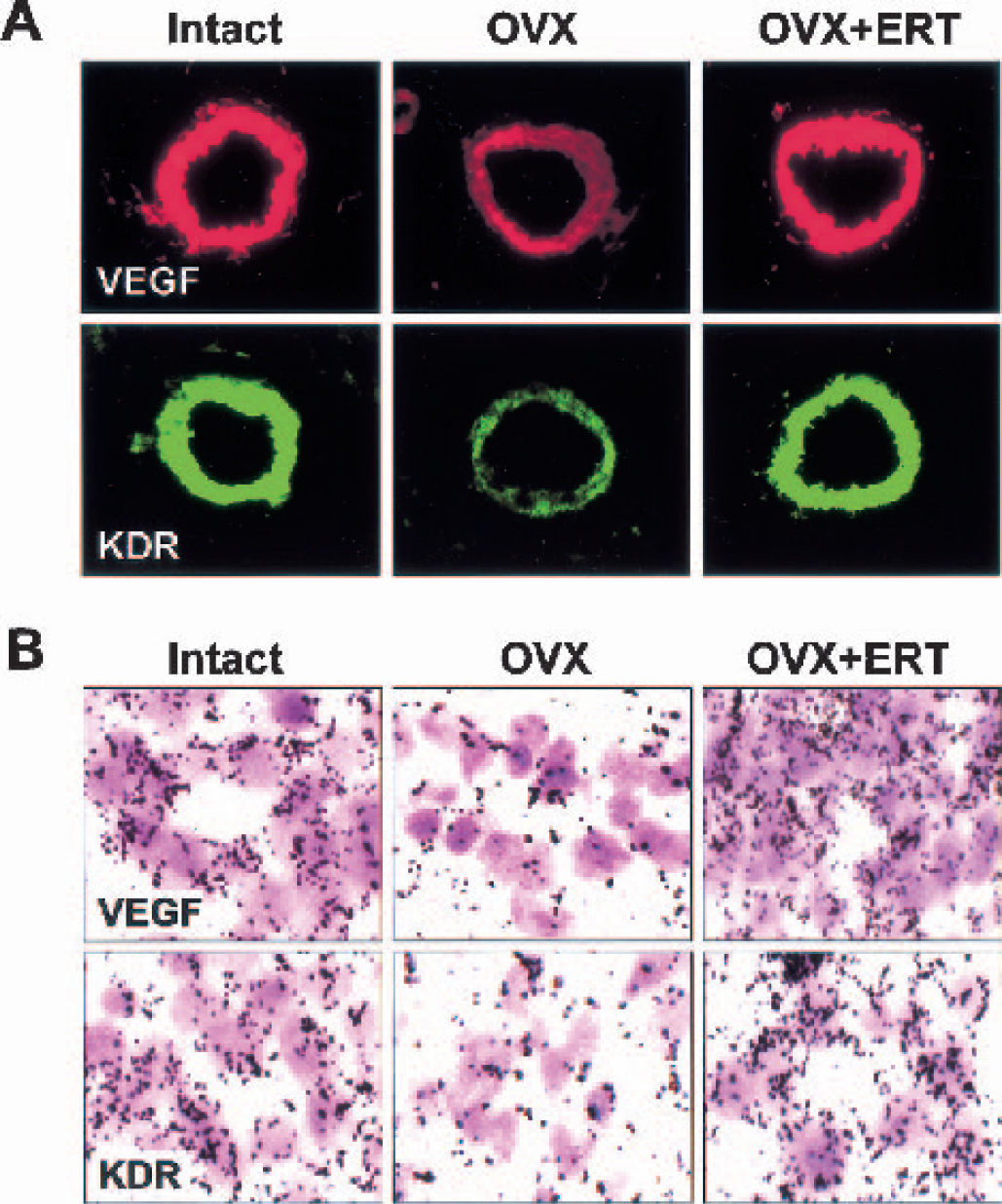
Protein and gene expression of vascular endothelial growth factor (VEGF) and anti—human fetal liver kinase 1 (kinase domain region [KDR]) in coronal sections of frontal cortex from middle-aged female rats that were intact, ovariectomized (OVX), and OVX with estrogen replacement therapy (ERT).
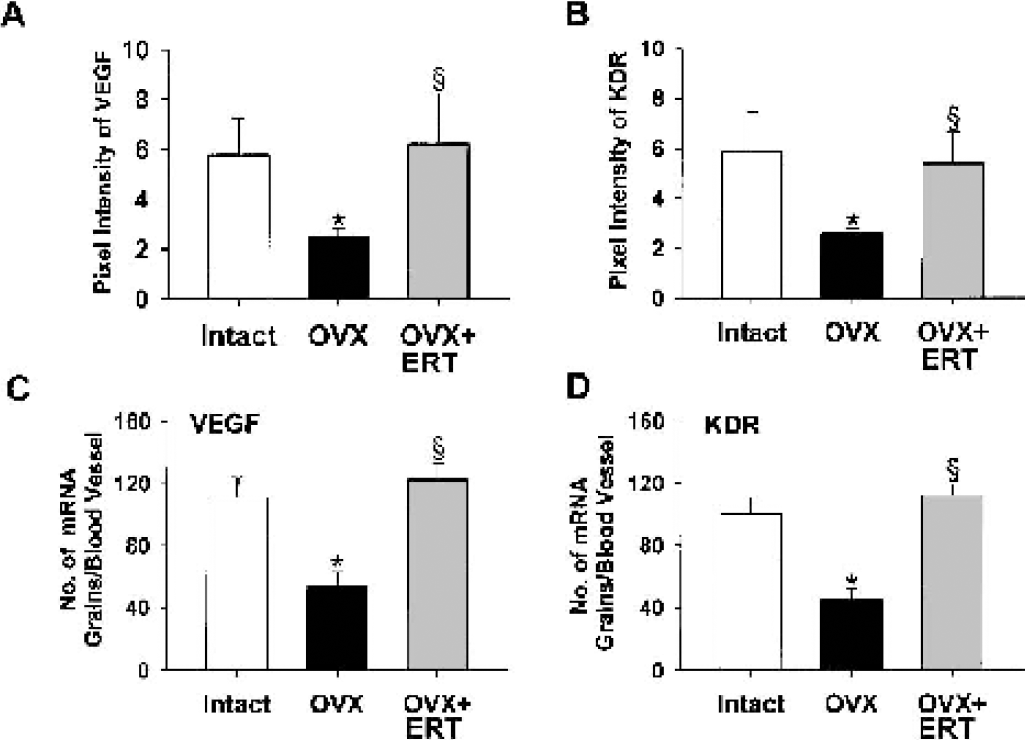
Quantification of vascular endothelial growth factor (VEGF) and fetal liver kinase 1 (kinase domain region [KDR]) proteins and messenger RNAs (mRNAs) in coronal sections of frontal cortex from middle-aged female rats that were intact, ovariectomized (OVX), and OVX with estrogen replacement therapy (ERT).
Immunohistochemical studies showed abundant eNOS protein expression in coronal sections of frontal cortex from intact middle-aged female rats (Fig. 4A). The location of positive staining for eNOS was largely in cerebral vessels, especially the intimal surface. The eNOS protein expression became evidently faint after ovariectomy; the expression was 53% that of intact animals (Fig. 4C). The decrease in positive staining for eNOS protein in cerebral vessels of ovariectomized rats correlated with a decrease in eNOS mRNA, which was obtained from in situ hybridization (Fig. 4B). The results of quantitative analysis showed a 46% decrease in eNOS mRNA expression after ovariectomy (Fig. 4D). Estrogen replacement therapy returned the reduced expressions of eNOS protein and mRNA after ovariectomy to the same levels as intact animals (Fig. 4).
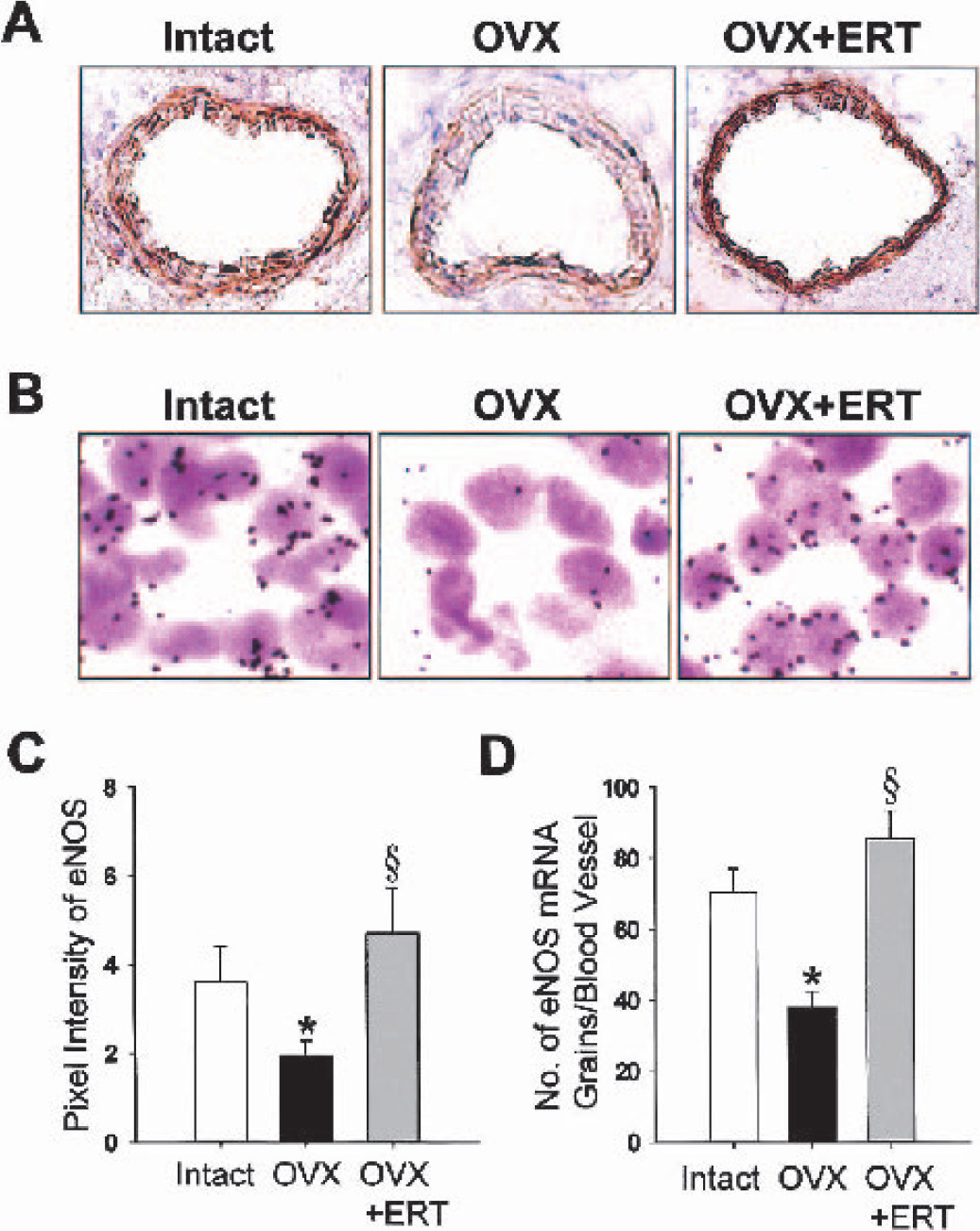
Protein and gene expression of vascular endothelial nitric oxide synthase (eNOS) in coronal sections of frontal cortex from middle-aged female rats that were intact, ovariectomized (OVX), and OVX with estrogen replacement therapy (ERT).
A marked decrease in immunofluorescent staining for Bcl-2, a survival factor that can block both necrotic and apoptotic cell death (Bredesen, 1995), was found in microvessels in coronal sections of frontal cortex from middle-aged female rats after ovariectomy (Fig. 5A). Ovariectomy resulted in a 46% decrease in vascular expression of Bcl-2 protein (Fig. 6A). In contrast, vascular expression of other members of the Bcl-2 family, Bax (Figs. 5B and 6B) and phosphorylated Bad (Figs. 5C and 6C), was not affected by ovariectomy. Estrogen replacement therapy completely prevented the ovariectomy-induced decrease in Bcl-2 protein (Figs. 5A and 6A).
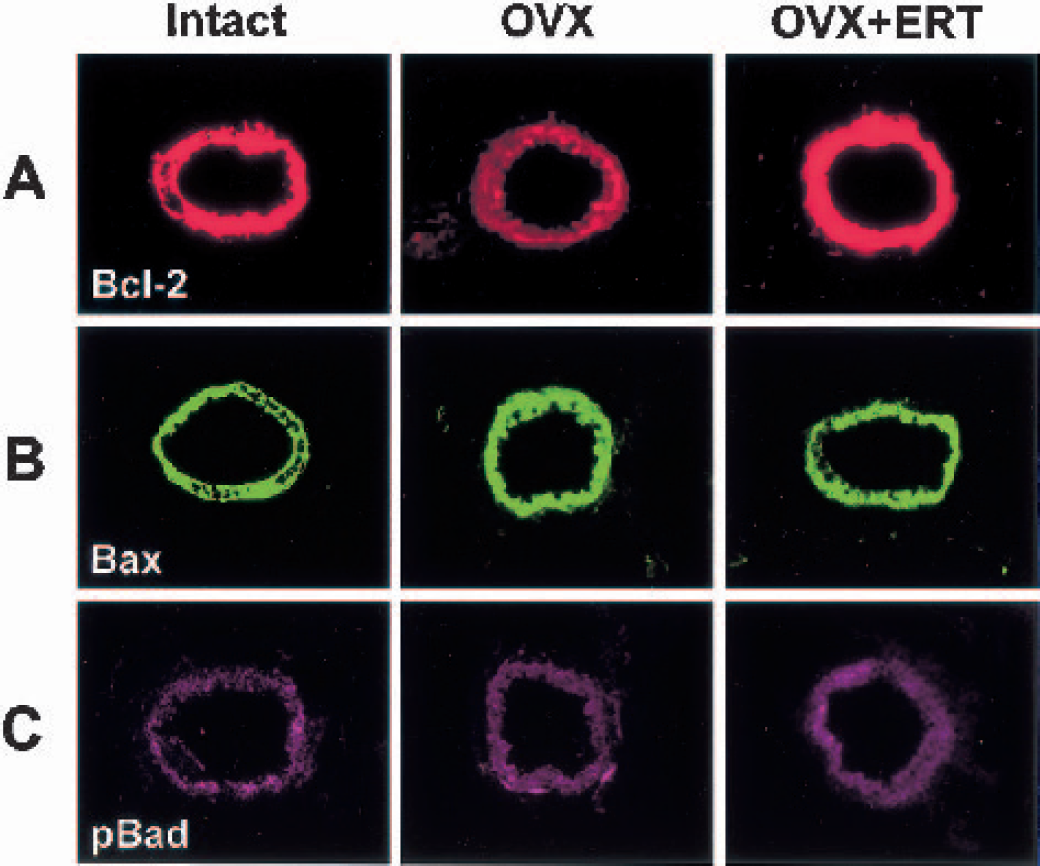
Immunofluorescent findings for Bcl-2
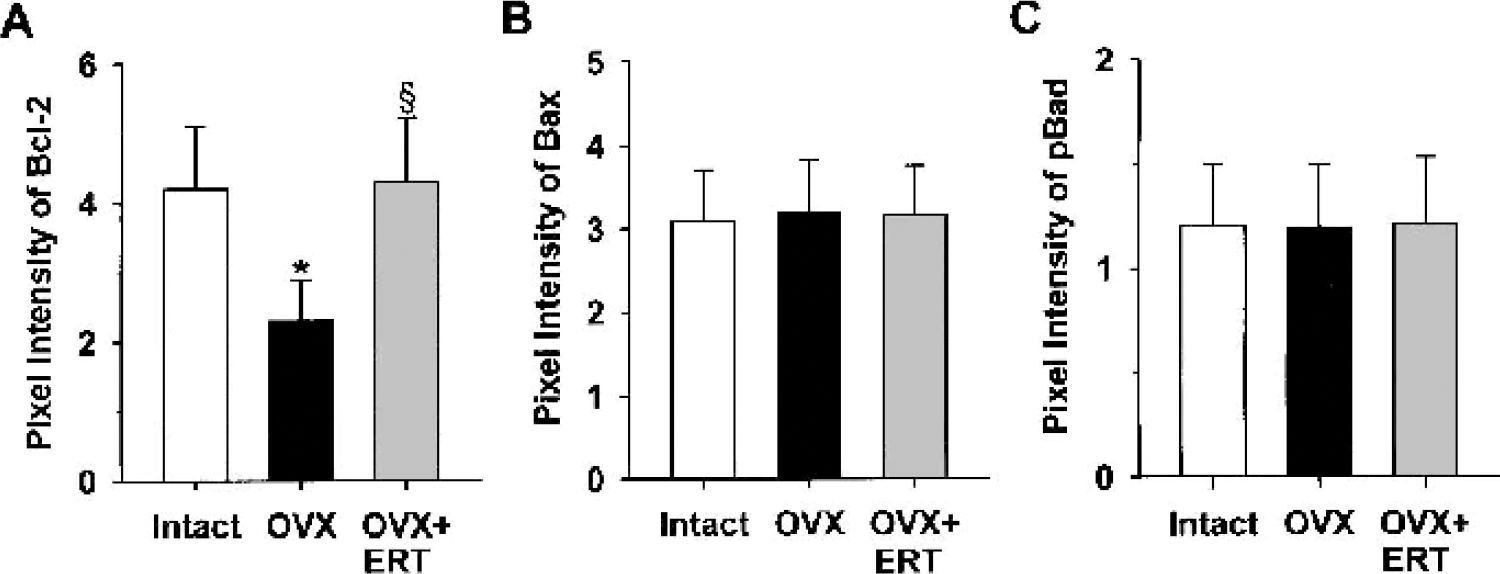
Quantitation of immunoreactivity for vascular Bcl-2
Double-label immunofluorescent staining for ERs in coronal sections of frontal cortex of middle-aged female rats showed that both ERα and ERβ were present in the nuclei of cerebral vessels (Fig. 7A). In intact animals, ERβ was more abundantly expressed than ERa, resulting in the ERβ/ERα ratio of 3.14 ± 0.10 (n = 20). When the animals were subjected to ovariectomy, ERβ expression was diminished more than ERα expression, and the ERβ/ERα ratio was changed to a significantly lower level (2.39 ± 0.09, n = 20, P < 0.01). Estrogen replacement therapy enhanced ERβ expression as seen in intact females and reversed the ERβ/ERα ratio (3.24 ± 0.12, n = 20, P < 0.01). The gene expression of ERs, examined by in situ hybridization (Fig. 7B), showed that the numbers of ERβ mRNA grains per cerebral vessel were about two times higher than those of ERα mRNA (P < 0.01). Although both ERα and ERβ mRNAs declined after ovariectomy, the extent of decline was more marked in ERβ than in ERα mRNA (60% versus 45%, P < 0.01). Estrogen replacement therapy significantly reversed their gene expressions nearly to the levels obtained in intact animals.
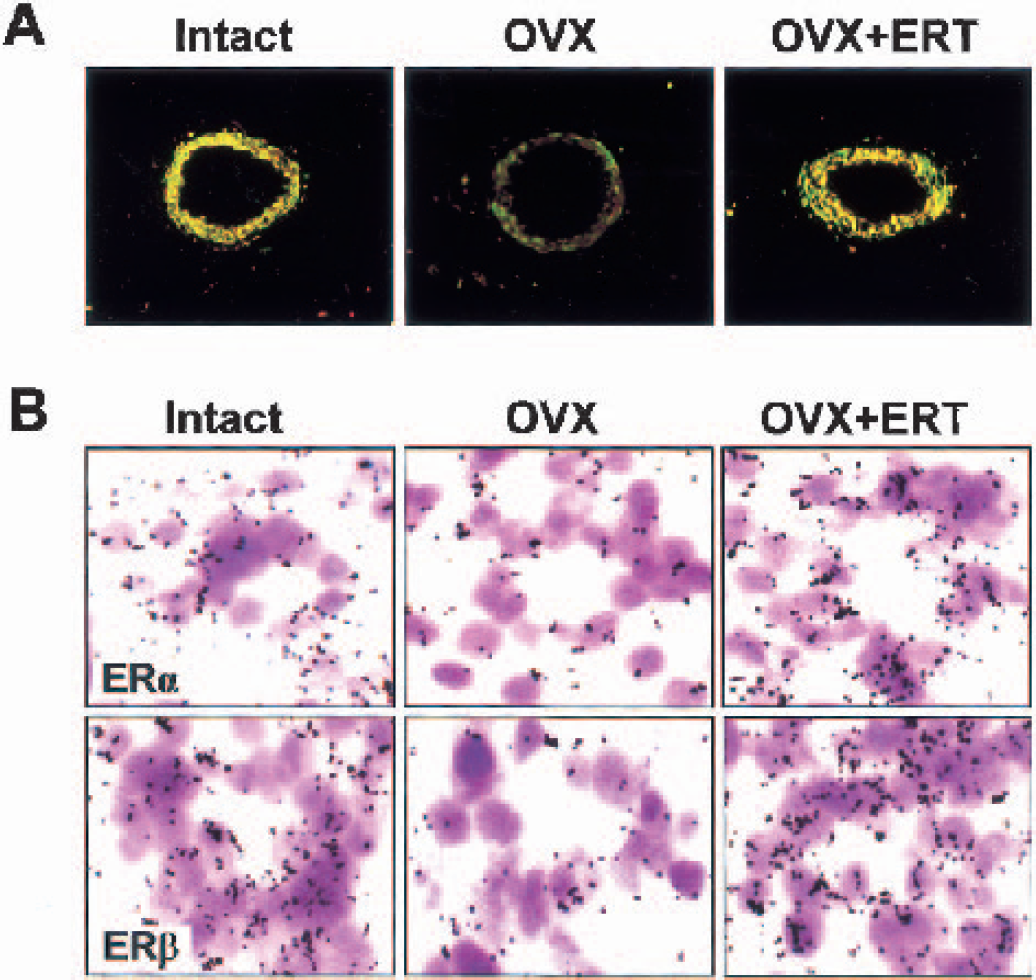
Protein and gene expression of vascular estrogen receptors (ER) in coronal sections of frontal cortex from middle-aged female rats that were intact, ovariectomized (OVX), and OVX with estrogen replacement therapy (ERT).
DISCUSSION
Although the effect of ERT on the risk of cerebrovascular events in postmenopausal women remains controversial, estrogen may confer protection against cerebrovascular disorders via diverse mechanisms, exerting both flow-preserving and other neuroprotective effects (Alkayed et al., 1998). The evidence, however, of flow preservation in animal models of cerebral ischemia is both supportive and unsupportive. Although there is lack of evidence of a beneficial effect of estrogen on cerebral blood flow during the occlusion phase of focal ischemia (Rusa et al., 1999; Carswell et al., 2000), estrogen may improve cerebral blood flow during the reperfusion phase of focal and global ischemia (Hurn et al., 1995; Pelligrino et al., 1998; Shi et al., 1998). Because estrogen plays a potential role in angiogenesis (Losordo and Isner, 2001), we hypothesized that the angiogenic action of estrogen may contribute to its flow-preserving neuroprotective effects. The present study focused attention on the effects of estrogen deprivation and replacement on the regulation of cerebral capillary development in frontal cortex during the period of middle age, when the estrous cycle becomes obscure, as confirmed by vaginal smears (Sakuma et al., 2002). Our experimental model would be appropriate to explore the neuroprotective mechanisms of estrogen because cerebrovascular events are well documented to occur more frequently in elderly women after menopause (Wenger et al., 1993). In addition, it may be stressed that the principal advantage of working in animal models is that the molecular effects of estrogen can be more readily defined. We clearly demonstrate that ovariectomy resulted in a significant reduction in the capillary density in frontal cortex of middle-aged female rats. Estrogen replacement therapy to ovariectomized rats, however, restored the capillary density to levels similar to those of the intact group. To our knowledge, this is the first study showing that estrogen regulates the development of cerebral capillaries in middle-aged females. Interestingly, plasma 17β-estradiol levels in middle-aged female rats (13 pg/mL) are much lower than in young females, which show plasma 17β-estradiol levels of 126 pg/mL at the estrous stage (Liu et al., 2001). Despite the low circulating estrogen levels, further reduction after ovariectomy resulted in a striking decrease in the cerebral capillary density in middle-aged female rats. This suggests that physiologic formation of cerebral capillaries at this age in females can be strongly swayed by subtle changes from critical concentrations of estrogen.
In angiogenesis, a variety of growth factors are released that affect one another, and it has been suggested that the angiogenic action of estrogen is mediated indirectly, via production of VEGF (Losordo and Isner, 2001). We found that the mRNA and protein expression levels of VEGF in cerebral vessels significantly decreased with estrogen deprivation and, with estrogen replacement, increased to levels similar to those seen in the intact group. This finding agrees with the suggestion that estrogen directly regulates VEGF gene transcription (Mueller et al., 2000). We also showed that expression of KDR, a receptor that mediates the angiogenic effect of VEGF (Ferrara, 2001) in cerebral vessels, was markedly downregulated at both mRNA and protein levels after ovariectomy, and ERT prevented the ovariectomy-induced decrease in KDR expression. In contrast, no significant changes were seen in expression of another VEGF receptor, Flt-1, which is devoid of angiogenic activities (Ferrara, 2001), in cerebral vessels of frontal cortex after estrogen deprivation or treatment. Thus, the regulation of VEGF and its angiogenic receptor KDR expressions by estrogen deprivation or treatment suggests that VEGF may participate in stimulating the formation of new capillaries from existing cerebral vessels in middle-aged rats. Furthermore, in the light of the fact that circulating estrogen concentrations are essentially low in middle-aged female rats, the dramatic reductions in VEGF and KDR after ovariectomy implies that subtle changes from critical concentrations of the sex hormone at this age could affect the expression levels of VEGF and KDR.
There is evidence indicating that eNOS acts downstream from VEGF in the angiogenic process (Ziche et al., 1997; Papapetropoulos et al., 1997). In vitro studies have shown that VEGF-induced endothelial cell migration and proliferation can be strongly attenuated by inhibition of eNOS (Shizuoka et al., 1999; Murohara et al., 1999). Furthermore, impaired angiogenesis in mice lacking the eNOS gene has not been improved by administration of VEGF (Murohara et al., 1998). In the present investigation, mRNA and protein expression of eNOS in cerebral vessels was markedly downregulated in middleaged female rats after ovariectomy and returned to intact levels with ERT. Such changes in eNOS expression, in addition to the altered VEGF expression levels, may critically affect VEGF-induced cerebral capillary angiogenesis in middle-aged females after estrogen deprivation or treatment. Several studies have shown an increase in eNOS protein in endothelial cell cultures after incubation with physiologic concentrations of estrogen (Hishikawa et al., 1995; Hayashi et al., 1995). However, the effects of estrogen deprivation and replacement on eNOS expression in cerebral vessels of middle-aged females cannot be solely attributed to the direct regulation by the sex hormone because middle-aged females are severely hypoestrogenic even when they are intact. Vascular endothelial growth factor—induced activation of its receptor KDR appears to lead to upregulation of eNOS protein (Kroll and Waltenberger, 1998), and VEGF has been reported to increase both eNOS mRNA and protein in endothelial cells in culture (Papapetropoulos et al., 1997). Therefore, the regulation of eNOS expression may be intimately linked with the VEGF expression levels in cerebral vessels of middle-aged females after ovariectomy or ERT.
Bcl-2, a cell survival factor, acts upstream to suppress caspase activation, inhibits free radical formation, regulates calcium sequestration, and prevents the proapoptotic actions of Bax or Bad (MacManus and Linnik, 1997; Merry and Korsmeyer, 1997). Although estrogen may directly upregulate this survival factor through receptor-mediated interactions with regions of the Bcl-2 promoter, which contains several putative estrogen-responsive sites (Teixeira et al., 1995), the effects of estrogen deprivation and replacement on Bcl-2 expression in cerebral vessels were strikingly parallel to their effects on VEGF expression. Indeed, activation of KDR with VEGF mediates upregulation of Bcl-2 (Dimmeler and Zeiher, 2000). The occurrence of endothelial cell apoptosis would have deleterious effects on the formation of new capillaries. Alternatively, the angiogenic activity attributed to VEGF may be mediated in part by its ability to enhance endothelial cell survival by inducing expression of Bcl-2 (Nör et al., 1999). We thus assume that regulation of Bcl-2 by VEGF, which depends on estrogen status, may play a critical role in the development of cerebral capillaries in middle-aged females by affecting the balance between endothelial cell viability and apoptosis. In our studies, the effects of estrogen deprivation and replacement were specific to Bcl-2, because expression of other members of the Bcl-2 family such as Bax and Bad were unaffected by differing hormonal status. Thus, the expression levels of proapoptotic proteins in cerebral vessels seems to remain constant regardless of whether middle-aged females were subjected to ovariectomy with or without ERT.
Previous studies suggest that physiologic estrogen levels may cause neuroprotection through receptor-dependent mechanisms (Singer et al., 1996; Dubal et al., 1998). Recent work using ERα- and ERβ-knockout mice in an animal model of stroke has discovered that ERα plays a pivotal role in the ability of physiologic estrogen levels to protect the cerebral cortex against ischemia (Dubal et al., 2001). It appears that the ERa-mediated protection against brain injury is through blood flow-independent neuroprotective mechanisms (Dubal et al., 2001). The present study showed that ERα and ERβ were significantly diminished in cerebral vessels at protein and mRNA levels when middle-aged female rats were subjected to ovariectomy. We found, however, that ovariectomy significantly decreased the ratio of ERβ/ERα in cerebral vessels. The ratio of receptor subtype expression may be crucial to understand how estrogen acts because each receptor can differentially activate certain response elements (Peach et al., 1997). The ovariectomy-induced decrease in the ERβ/ERα ratio indicates a potentially intriguing role for ERβ in the ability of estrogen to promote the development of cerebral capillaries. Thus, it is interesting to speculate that estrogen may act through ERβ to regulate expressions of VEGF and its angiogenic receptor KDR in cerebral vessels, thereby contributing to flow-preserving neuroprotection. Further studies, however, are necessary to definitively link VEGF and KDR with ERβ-targeted gene expression.
In summary, our results hint that VEGF may be a critical regulatory molecule for physiologic cerebral angiogenesis in hypoestrogenic elderly women. We suggest that VEGF, at least in part, plays a key role in the flow-preserving neuroprotective action of estrogen through its angiogenic and antiapoptotic properties. Despite the fact that circulating estrogen levels were essentially low in middle-aged female rats, estrogen deprivation and replacement dramatically changed vascular VEGF expression and consequently capillary morphology. The data imply that elderly postmenopausal women may benefit from ERT that uses relatively low doses of hormone, although many other factors involved in the estrogen-mediated neuroprotective mechanisms must clearly be taken into consideration.
Footnotes
Acknowledgments
The authors thank Prof. Toshihiko Iwanaga for his kind advice and suggestions.