
Abstract
Structural and functional reorganization in the vicinity of damaged neocortex and other connected brain areas seems to be responsible for postlesional functional recovery. To better understand the molecular mechanisms underlying this type of plasticity, gene expression patterns were analyzed by using DNA macroarrays comprising 1176 genes. Circumscribed unilateral infarcts consistently affecting the forelimb area of the motor cortex were induced photochemically in adult rats. Ten days after lesioning, cortical gene expression fingerprints were evaluated from an area adjacent to the lesion as well as two contralateral areas of motor and somatosensory cortex. Discrete regions showed distinct expression patterns. Upregulation was observed of different members of transcription factors, immediate early genes, neuronal signaling as well as neuronal growth and structure-associated genes, ipsilaterally (six genes) and/or contralaterally (eight genes in the motor and seven in the somatosensory cortex). In contrast, downregulations were restricted to ipsilateral areas and included genes coding for ion channels, transport proteins, mediators of metabolic pathways, and intracellular transducers (14 genes). A subset of these regulations were further confirmed by real-time polymerase chain reaction (TaqMan assay). At least part of the detected regulations, in particular those of the contralateral hemisphere, are likely to underlie plasticity processes.
Ample evidence indicates that after brain damage, a profound reorganization in the spared cerebral cortex occurs that in part contributes to recovery (Stroemer et al., 1995; Nudo and Milliken, 1996; Jones et al., 1996; Rossini et al., 1998; Witte et al., 2000; Cramer et al., 2000). The molecular basis of these plasticity changes that involve dynamic orchestration and crosstalk of a variety of molecules is still poorly understood. With the development of cDNA array technology, a method has become available that allows parallel expression monitoring of a high number of genes: an exploratory method with a high throughput when compared with traditional techniques, such as differential display polymerase chain reaction (PCR) or subtractive hybridization. Here, we report the use of commercial arrays containing 1,176 genes representing a broad range of functional classes. In contrast to microarrays, these macroarrays can be analyzed using common laboratory equipment that simplifies their handling. The model of photothrombotic ischemic lesion was used because of its high reproducibility concerning the size and localization of the lesion and its penumbra. Ten days after the lesion, the pattern of gene expression in discrete cortical brain regions on the ipsilateral and contralateral side was compared with changes in homologous regions of sham-operated littermates. This represents the first report of using gene arrays in expression profiling of both adjacent and contralateral cortex areas to an ischemic brain lesion.
MATERIALS AND METHODS
Animals, tissues
The experiments were performed on a total of 12 adult male Wistar SPF rats (290 to 310 g). Photothrombotic cortical lesions (3.5 ± 0.5 mm in diameter) affecting the forelimb area of the primary motor cortex were induced photochemically in eight animals (Watson et al., 1985). Briefly, using enflurane anesthesia (2% in N2O/O2), the skull of the animals was exposed, and a fiberoptic bundle (aperture, 3.0 mm) was positioned 1 mm anterior to bregma and 3 mm lateral to the mid-sagittal suture (forelimb area of primary motor cortex). The illumination through the skull (20 minutes) was initiated simultaneously with the application of the dye rose bengal through a femoral vein catheter. Rectal temperature was kept at 37° ± 0.5°C throughout the whole procedure. After surgery rats were returned to their cages with food and water available ad libitum. Four sham-operated controls underwent the same procedure except for switching on the cold light. Lesions in this position, as shown in our previous studies (Keyvani et al., 2000), produce a significant reduction in use of contralateral forelimb with subsequent partial recovery within 1 week. Ten days after lesion induction, the animals were decapitated while under deep anesthesia. After removing the brains, tissues were sampled by dissecting out a round cortex area of 4-mm diameter on the posterolateral side of the lesion as well as two contralateral areas of motor and somatosensory cortex (Fig. 1). For dissecting the ipsilesional area, we always kept a distance of 1.0 to 1.5 mm from the lesion (which is at this time point completely necrotic and clearly demarcated) to avoid contamination with the damaged region. Homologous cortex areas of sham-operated rats were extracted as control probes. The tissues were flash frozen in liquid nitrogen, and stored at −80°C until use. Comparisons were performed between the cortex areas of lesioned rats versus the homologous areas of the control animals. All experimental procedures were in accordance with the guidelines of the local animal care commission.
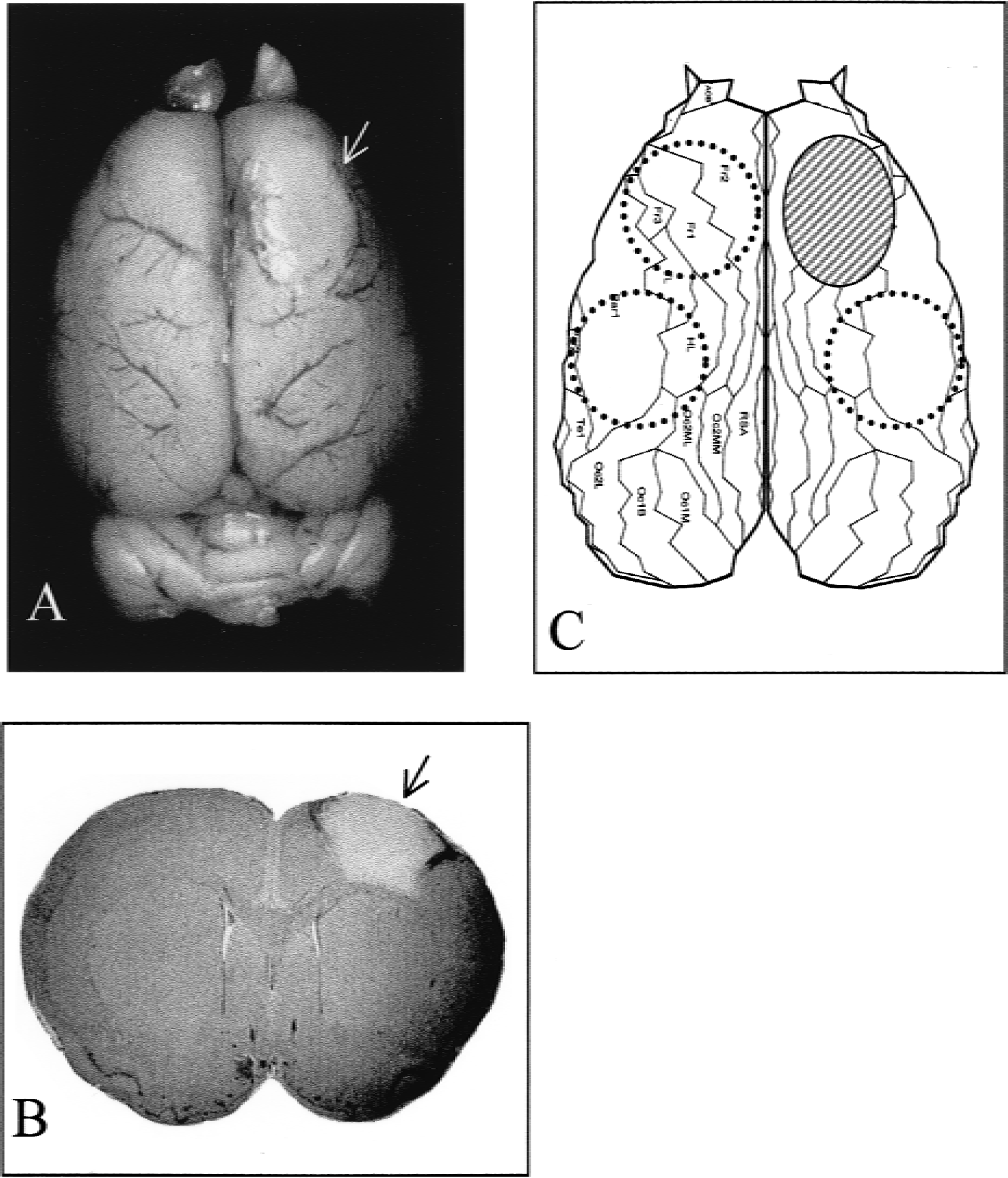
Lesion in rat motor cortex. (
RNA extraction, labeling, and hybridization
Total RNA was isolated using Trizol reagent (Gibco BRL, Eggenstein, Germany) according to the instruction of the manufacturer with the exception that the aqueous phase of the first extraction was reextracted with Trizol reagent. Before reverse transcription, DNAse treatment was performed using DNAse I (Boehringer Mannheim, Penzberg, Germany). DNAse was removed by an additional round of Trizol extraction. DNA contamination was excluded by performing PCR for glyceraldehyde 3-phosphate dehydrogenase (GAPDH) in RNA sample versus cDNA samples as positive control (for primers see Table 1). The RNAs isolated from identical cortical regions of the eight lesioned rats were pooled. The same was done for the control group. RNA quality was assessed by gel electrophoresis, as well as by OD260/280 nm ratios. Atlas Rat 1.2 arrays (Clontech Laboratories, Palo Alto, CA, U.S.A.) containing cDNA fragments of 1,176 genes were used. cDNA synthesis, labeling, hybridization, and washing of the membranes were carried out according to manufacturer's instructions. Briefly, 4-μg total RNA was used as template for cDNA synthesis, which was done in the presence of dATP. The labeled probes were purified by spin-column centrifugation (Atlas Nucleo-Spin, Clontech Laboratories, Palo Alto, CA, U.S.A.), and hybridization was carried out overnight at 68°C in a rotation hybridization oven using probes meeting the manufacturer's recommendation for specific activity (4–10 × 106 cpm). The membranes were then washed (four times with 2x standard sodium citrate, 1% sodium dodecyl sulfate [SDS], and once with 0.1x standard sodium citrate, 0.5% sodium dodecyl sulfate, both at 68°C, followed by 2x standard sodium citrate at room temperature) and exposed for 1 to 3 days to an Imaging Plate (BAS-MS 2040, Fuji, Kanagawa, Japan). Membranes were stripped according to the manufacturer's instructions and reused twice. We compared Atlas Arrays coming from the same lot, and only after parallel hybridization and exposure.
Primers and TaqMan probes

PCR, polymerase chain reaction; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; bp, base pair; NGFI-B, nerve growth factor–induced protein B.
Signal analysis
Spot densitometries were performed by a phosphorimaging system (Fujix Macbas 1000 Bioimaging Analyzer, Kanagawa, Japan) and related software (TINA 2.1 Raytest, Straubenhardt, Germany). Grids were applied to the image of the blot to quantify hybridization intensity of each spot. Background signal was defined as the average value of the signal intensities of representative membrane regions outside of the spot areas. A gene was defined as being expressed if the hybridization signal exceeded the background signal by at least two times. Before comparison analysis, all hybridization signals were normalized to the average intensities of GAPDH and beta-actin housekeeping genes. A gene was regarded as upregulated if the intensity of an expressed gene in the lesion membrane was ⩾ 1.7 times higher than that of the corresponding spot in the sham membrane. A gene was regarded as downregulated if the intensity of an expressed gene in the lesion membrane was ⩽0.6 times smaller than that of the corresponding spot in the sham membrane. All hybridization experiments were performed three times using independent cDNA-synthesis products. The regulations reported in this study could be corroborated consistently in the replicates.
Quantitative real-time PCR (TaqMan)
To validate these results, the expression of a subset of these regulations was examined by performing real-time quantitative PCR. We applied TaqMan assay and an ABI PRISM 5700 Sequence Detection System (Applied Biosystems, Foster City, CA, U.S.A.) as described previously (Heid et al., 1996). The same RNA pools as described herein were used after DNAse I treatment (Boehringer Mannheim) for cDNA synthesis using Superscript II reverse transcriptase system (Gibco BRL) according to the manufacturer's protocol. The PCR primers and TaqMan probes for nerve growth factor–induced protein B (NGFI-B) and beta-actin (Table 1) were designed using Primer Express software version 2.0 (Applied Biosystems). Reporter dyes and quencher dyes for all probes were 6-carboxyfluorescein (FAM) and 6-carboxytetramethylrhodamine (TAMRA), respectively. The constituents of each PCR (50 μL) were 1 μL cDNA (or dH2O), 2x (forward and reverse) 1.5 μL primer (300 nmol/L each), 2.5 μL TaqMan probe (250 nmol/L), 18.5 μL H2O, and 25 μL TaqMan Universal PCR Mastermix (Applied Biosystems). Product amplification was done for 40 PCR cycles, after uracil removal (2 minutes at 50°C) and polymerase activation (10 minutes at 95°C). Each two-step PCR cycle comprised denaturation (15 seconds at 95°C), annealing, and extending (1 minute at 60°C). To exclude variability of RNA concentration, beta-actin was used as an internal control. For relative quantification of the amount of specific mRNA, a standard curve was generated for each run using a twofold-dilution series. NGFI-B and beta-actin mRNA in each sample were determined by the Ct values (PCR cycle number at which the reporter dye fluorescence is detectable above an arbitrary threshold) of the TaqMan PCR according to its standard curve. All assays were run in triplicate.
RESULTS
On average, 30% (± 4%) of genes represented on the array were expressed. The expression levels of the two housekeeping genes, GAPDH and beta-actin, which were used as internal controls did not change significantly from region to region (range of changes, 0.9- to 1.2-fold). Typical images of scanned arrays are shown in Figure 2.
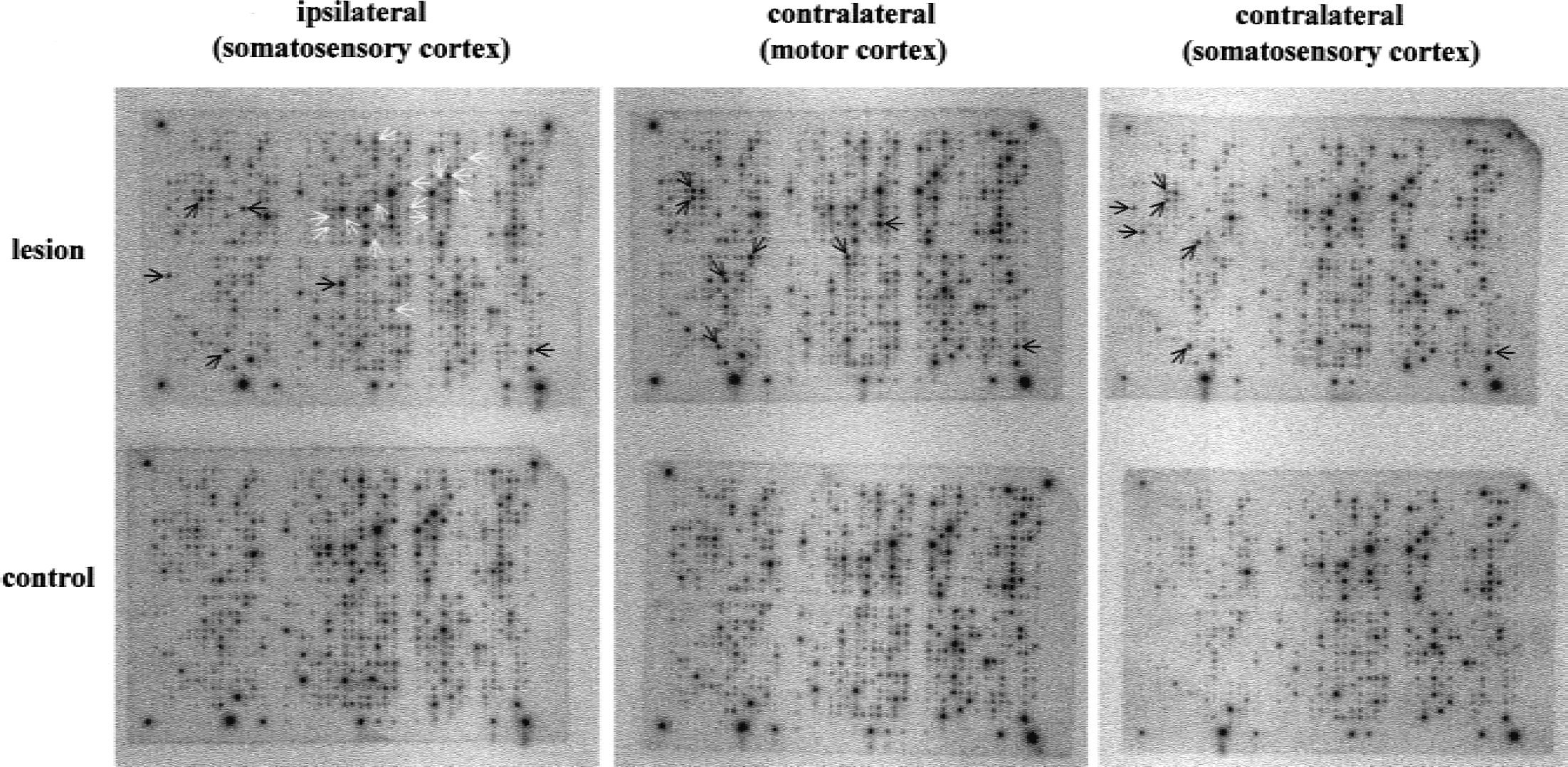
Gene expression patterns of three different cortical areas in ischemic and control brain. Representative images of scanned arrays hybridized with cortical samples of lesioned
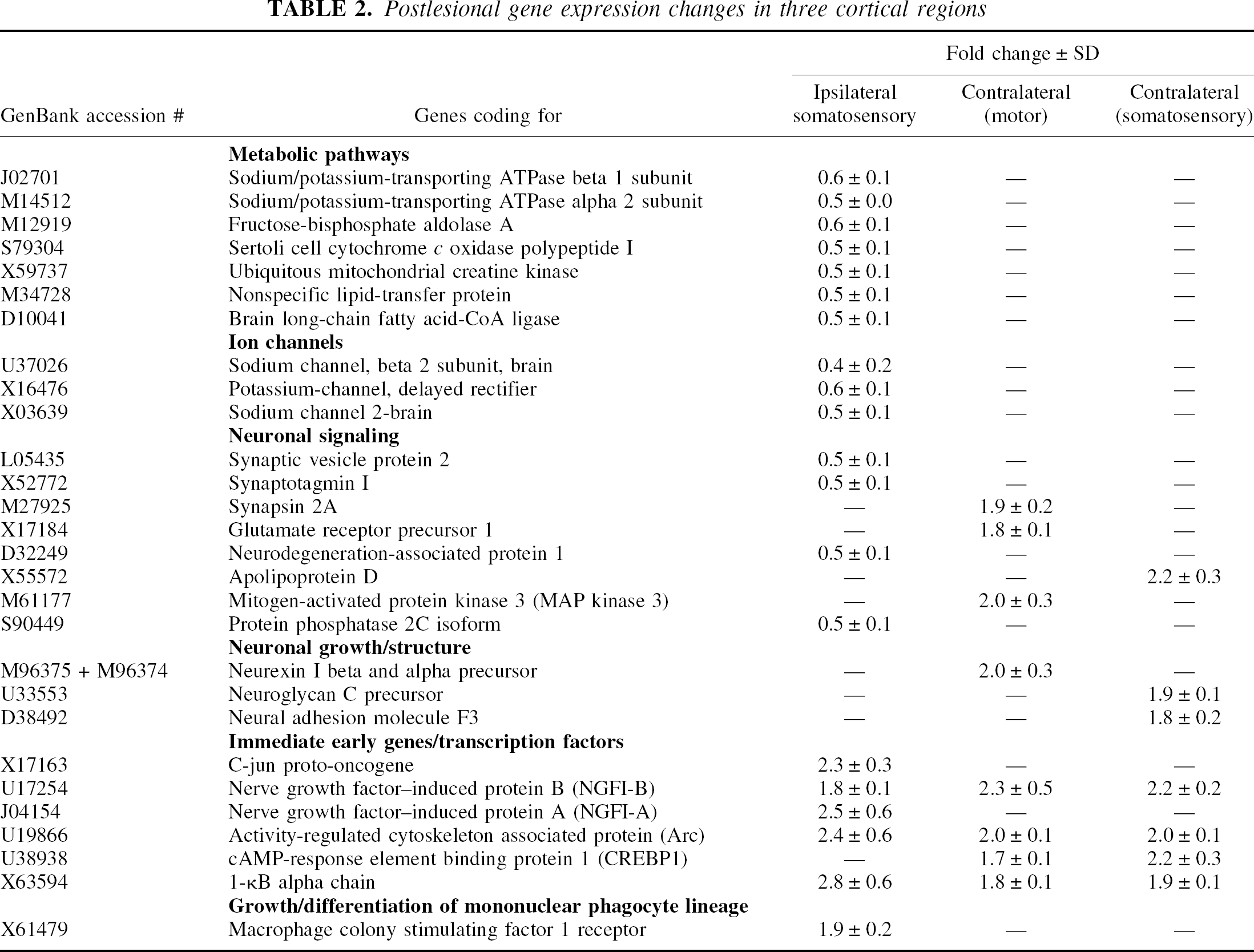
Discrete regions responded with distinct gene expression profiles to the ischemic event. Table 2 summarizes all transcripts with altered expression in the three examined cortex areas. In the ipsilateral perilesional cortex, 20 genes showed an expression alteration. Among 14 genes that were downregulated by 1.7- to 2.5-fold, 7 could be identified as enzymes of metabolic pathways, 3 as ion channels, and 2 as synapse-related genes. Neurodegeneration-associated protein 1 and protein phosphatase 2C, both involved in neuronal signaling processes, were also suppressed. Five of the six genes whose expression was increased in the perilesional cortex (1.8- to 2.8-fold) were immediate early genes and/or transcription factors. Additionally, macrophage colony-stimulating factor 1 receptor was found to be induced in this region. The contralesional cortex showed exclusively upregulations (1.7- to 2.3-fold), including eight genes in the homotopic motor cortex and seven in the adjacent somatosensory cortex. These transcripts could be categorized as genes involved in neuronal signaling, growth, or structure, as well as transcription factors/immediate early genes. I-kB alpha, NGFI-B, and activity-regulated cytoskeleton associated protein (Arc) were found to be upregulated in all three regions. Furthermore, CREBP1 was induced in both contralateral cortices, but not ipsilaterally.
Postlesional gene expression changes in three cortical regions
A subset of the regulations already mentioned was confirmed by using real-time PCR (TaqMan). Analysis of mRNA levels for NGFI-B in three different cortical areas showed a clear increase of its expression level when compared with analogous cortical areas of sham-operated animals (Fig. 3). The fold changes ± SD of NGFI-B were 2.6 ± 0.1 in the ipsilesional somatosensory cortex, 3.9 ± 0.2 in the contralesional motor cortex, and 2.8 ± 0.2 in the contralesional somatosensory cortex. In contrast, the level of beta-actin in experimental versus control samples remained unchanged (fold change range: 0.9–1.1).
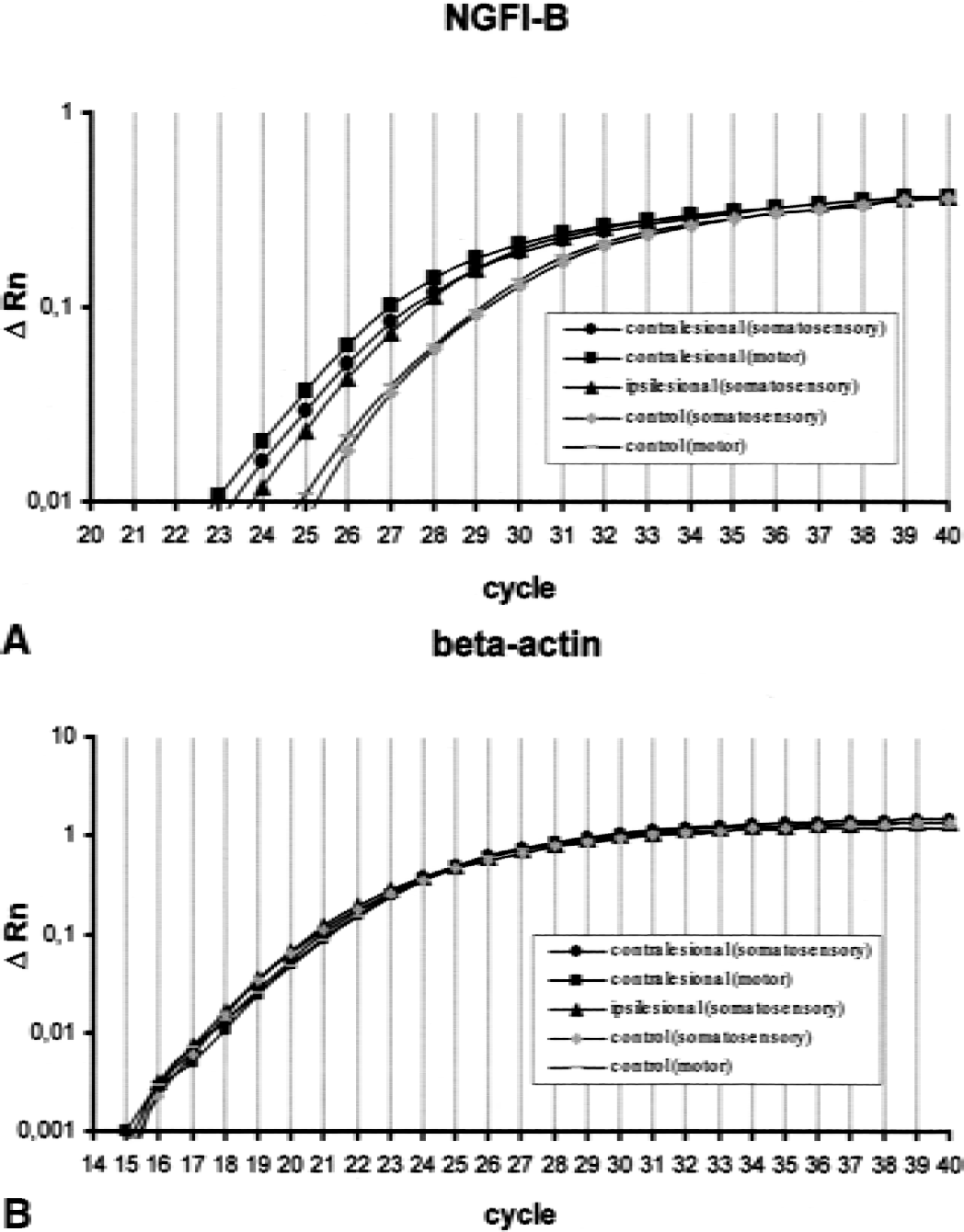
Amplification plots showing the increase in reporter fluorescence over 40 cycles.
The standard curves of both genes showed a linear relationship between the log of the initial template amount and the threshold cycle (data not shown).
DISCUSSION
The present study provides an evaluation of changes in gene expression in three discrete cortical areas 10 days after a circumscribed ischemic lesion in the motor cortex.
To perform an independent validation of the macroarray data, we used a quantitative PCR method (TaqMan) to analyze a subset of these regulations (Fig. 3). Overall, changes in mRNA abundance seem to be underestimated by macroarray analysis compared with the TaqMan assay, although general agreement between the two methods was observed.
One conspicuous change in the ipsilateral cortex was downregulation of genes coding for enzymes involved in cell metabolism, including sodium/potassium-transporting ATPase (alpha and beta subunits), fructose-bisphosphate aldolase, ubiquitous mitochondrial creatine kinase, and brain long-chain fatty acid–coenzyme A ligase. The phenomenon of postlesional cerebral hypometabolism has been reported previously (Baron et al., 1986; Kiyosawa et al., 1987; Iglesias et al., 2000), and is thought to be caused by nonspecific degenerating processes (e.g., as a result of deafferentiation). In principle, it is difficult to dissociate molecular events reflecting impaired metabolism from adaptive processes counteracting the deleterious effects of ischemia, particularly if perilesional brain areas are addressed. Therefore, we think that the functional interpretation of dysfunction is more likely, whereas the alternate interpretation as compensatory mechanisms cannot be excluded.
Sodium and potassium channels, synaptic vesicle protein 2, synaptotagmin I, and protein phosphatase 2C were also downregulated at the transcriptional level in the ipsilateral hemisphere. All these genes are involved in neuronal signaling processes. To particularize, sodium or potassium channels are fundamental to electrical signaling in the nervous system. The properties of dendritic sodium channels are important for action potential–mediated effects on synaptic integration and plasticity in neurons (Jung et al., 1997). Protein phosphatase 2C is one of four major serine-threonine specific phosphoprotein phosphatases that modulate intraneuronal signal transduction cascades (Abe et al., 1992), and synaptic vesicle protein 2 (Janz et al., 1999) as well as synaptotagmin I (Littleton et al., 1993) play an integral role in neurotransmission. Furthermore, neurodegeneration-associated protein 1 (Neurodap1) was also found to be ipsilaterally downregulated. The product of this axotomy-suppressed gene is bound to the postsynaptic density region of axosomatic synapses, and is supposed to play a role in synaptic communication and plasticity through the control of the formation of postsynaptic density (Nakayama et al., 1995). Given that plasticity includes not only a dynamic process of sprouting, facilitation, and potentiation but also pruning and depression, the question seems reasonable if at least part of these downregulations may be a component of molecular changes underlying postlesional plasticity, and not solely a nonspecific appearance of the local pathophysiology.
Macrophage colony-stimulating factor 1 receptor, which was upregulated in the perilesional cortex, seems to be essential for proliferation and activation of microglia. The upregulation of this gene may reflect microglia activation and could play a central role in preparing macrophage-related cells to take part in the cellular response to the necrotic infarct area (Raivich et al., 1998).
Several members of the immediate early gene and/or transcription factor family were bilaterally upregulated. These are genes that stand at the beginning of a cascade of molecular changes with variable target pathways. Their multifaceted cause and effect of expression is probably the reason for their overrepresentation among the detected upregulations of genes in this study. Transcript inductions were observed for NGFI-B, activity-regulated cytoskeleton associated protein (Arc), and I-κB alpha chain, bilaterally. Furthermore, NGFI-A and c-jun were ipsilaterally upregulated, and cAMP-response element binding protein 1 (CREBP1) was contralaterally upregulated. The expression of NGFI-A, NGFI-B (Honkaniemi et al., 1997; Soriano et al., 2000), Arc (Kunizuka et al., 1999; Soriano et al., 2000), c-jun (Gubits et al., 1993), and CREB (Tanaka et al., 1999) has been already reported in association with ischemia within the first 24 postlesional hours on the side of lesion. In addition, all of these genes are associated with changes related to neuronal plasticity. NGFI-A and NGFI-B are thought to be involved in the sleep-waking cycle (Cirelli and Tononi, 2000) or cortical changes as a consequence of environmental enrichment (Mohammed et al., 1993; Wallace et al., 1995). Arc seems to play a role in activity-dependent plasticity of dendrites (Lyford et al., 1995) and long-term potentiation (Guzowski et al., 2000). CREB is an evolutionarily conserved transcription regulator essential for long-term memory formation (Bartsch et al., 1998; Barth et al., 2000). I-κB is a regulator of NF-κB, which is a potent transcriptional activator involved in synaptic activity (Meberg et al., 1996). Jun transcription factors are thought to play an essential role not only in fundamental processes such as cell proliferation, differentiation, and programmed neuronal cell death, but also in complex processes such as plastic adaptations in the mature brain (Schlingensiepen et al., 1994).
The expression of three genes associated with neuronal structure changed in the contralateral hemisphere. Neurexin I precursor was upregulated in the contralateral motor cortex. Neuroligin and neurexin form an intracellular adhesion complex. They are supposed to be part of the machinery used during formation and remodeling of CNS synapses (Scheiffele et al., 2000). Neuroglycan C precursor and neuronal adhesion molecule F3 were both induced in the contralateral somatosensory cortex.
Neuroglycan C is a transmembrane chondroitin sulfate proteoglycan present exclusively in CNS and involved in synaptoneogenesis and formation of neuronal networks in developing cerebellum and retina (Aono et al., 2000; Inatani et al., 2000). F3, on the other hand, is an axon-associated glycoprotein that seems to intervene in activity-dependent structural axonal plasticity (Pierre et al., 1998).
Finally, a number of genes associated with neuronal signaling were also upregulated in the contralateral hemisphere. Synapsin 2A, a synaptic vesicle protein, essential for regulating neurotransmitter release (Hosaka et al., 1999), MAP kinase 3, a critical regulator of cell division and differentiation with functions in learning and memory processes (Selcher et al., 1999; Bolshakov et al., 2000), as well as GluR1 precursor, an AMPA receptor gene (Hayashi et al., 2000), were induced in the contralateral motor cortex. In the somatosensory cortex of the contralesional hemisphere, apolipoprotein D was found to be upregulated. The product of this gene, a protein of the lipocalin family, is likely to be a multiligand, multifunctional transporter. While apolipoprotein E is thought to be involved in maintaining the integrity of the synaptodendritic complex (Masliah et al., 1996), the physiological function of apolipoprotein D is unknown (Terrisse et al., 1999).
Based on these data, we conclude that most observed gene upregulations, particularly on the contralateral side, may reflect molecular processes rearranging the cerebral cortex. It has been previously demonstrated that lesions in the forelimb area of the motor cortex (as in this report) produce significant behavioral asymmetry, i.e., a neglect of contralateral forelimb or a preferential use of the ipsilateral limb (Jones and Schallert, 1994; Keyvani et al., 2000). Thus, besides the lesion as a stimulus, a use dependency should be also taken into account as an eliciting factor for the molecular rearrangements, apparently occurring in the connected motor and somatosensory cortices.
Taken together, 10 days after an ischemic lesion affecting the rat motor cortex, region-specific changes of gene expression appear to occur. The expression pattern in the perilesional cortex differed substantially from that of the contralateral side. Furthermore, also in the contralesional hemisphere, the motor and somatosensory cortices showed different expression patterns. At least part of the detected regulations, in particular those of the contralateral hemisphere, are likely to be related to plasticity processes. The challenges that lie ahead are to characterize the function of these genes and to identify those that can serve as potential therapeutic targets for improving postlesional brain recovery.
Footnotes
Acknowledgments:
We thank Daniela Steinhof and Ulrike Botschen for skillful technical assistance.