
Abstract
The integrity of cerebral microvessels requires the close apposition of the endothelium to the astrocyte endfeet. Integrins α1β1 and α6β4 are cellular matrix receptors that may contribute to cerebral microvascular integrity. It has been hypothesized that focal ischemia alters integrin expression in a characteristic time-dependent manner consistent with neuron injury. The effects of middle cerebral artery occlusion (MCAO) and various periods of reperfusion on microvasclar integrin α1β1 and α6β4 expression were examined in the basal ganglia of 17 primates. Integrin subunits α1 and β1 colocalized with the endothelial cell antigen CD31 in nonischemic microvessels and with glial fibrillary acidic protein on astrocyte fibers. Rapid, simultaneous, and significant disappearance of both integrin α1 and β1 subunits and integrin α6β4 occurred by 2 hours MCAO, which was greatest in the region of neuron injury (ischemic core, Ic), and progressively less in the peripheral (Ip) and nonischemic regions (N). Transcription of subunit β1 mRNA on microvessels increased significantly in the Ic/Ip border and in multiple circular subregions within Ic. Microvascular integrin α1β1 and integrin α6β4 expression are rapidly and coordinately lost in Ic after MCAO. With loss of integrin α1β1, multiple regions of microvascular β1 mRNA up-regulation within Ic suggest that microvessel responses to focal ischemia are dynamic, and that multiple cores, not a single core, are generated. These changes imply that microvascular integrity is modified in a heterogeneous, but ordered pattern.
Exclusively in the brain, microvascular endothelial cells and astrocytes contribute to the permeability barrier and basal lamina (extracellular matrix [ECM]), which protect the central nervous system from edema and hemorrhage (Risau and Wolburg, 1990). The adhesion of astrocyte endfeet to the ECM ligands within the basal lamina is presumed to occur in part through integrin α6β4(Wagner et al., 1997). Endothelial cell adhesion to the ECM also is mediated by integrin–matrix ligand binding (Chintala et al., 1996; Helmer et al., 1987; Sonnenberg, 1993; Tuckwell et al., 1994; Webersinke et al., 1992). Although the endothelial cell integrins α1β1, α2β1, α3β1, and α6β1 associate with matrix within most vessels (Albelda and Buck, 1990; Faull and Ginsberg, 1995; Hynes, 1987; Ruoslahti, 1991), integrin α1β1 (VLA-1) is predominantly expressed on noncerebral microvascular endothelial cells in vivo (Defilippi et al., 1991). Integrin subunits α1 and β1 also are expressed in nearly equivalent distribution in all normal cerebral microvessels in the nonhuman primate (Haring et al., 1996a). These subunits are found on microvascular endothelial cells of nonischemic brain in association with the 130 kDa adhesion and signaling molecule CD31 (Haring et al., 1996a; Sun et al., 2000; Yang et al., 1999). The laminins and type IV collagen, ligands for integrin α1β1, and laminin-5, the ligand for α6β4, are abundant constituents of the cerebral microvascular basal lamina (Gehlsen et al., 1989; Hamann et al., 1995; Vandenberg et al., 1995).
The cerebral microvasculature is generally regarded to be less sensitive than neurons to focal ischemia. Although the basal lamina antigens laminin-1, laminin-5, collagen IV, and cellular fibronectin are lost from cerebral microvessels after middle cerebral artery occlusion (MCAO) (Hamann et al., 1995; Wagner et al., 1997), several measures of microvascular reactivity suggest a more rapid response in keeping with neuron injury. Using in situ methods to detect DNA scission, evidence of neuron injury appears within 2 hours in the nonhuman primate basal ganglia after MCAO, much earlier than previously shown in other models (Tagaya et al., 1997). Integrin αVβ3 and vascular endothelial growth factor are rapidly expressed on smooth muscle cells in select activated precapillary arterioles in close spatial and temporal relation to neuron injury detected by DNA scission (Abumiya et al., 1999; Tagaya et al., 1997). In addition, the expressions of latent metalloproteinase-2 and urokinase, which may play roles in basal lamina dissolution, increase 1 to 2 hours after MCAO (Heo et al., 1999). These early responses suggest that microvascular endothelial cell and astrocyte integrin receptor expression may be equally sensitive to focal ischemia and also related to neuron injury.
Those observations in conjunction with the known association of integrins α1β1 and α6β4 with cerebral microvessels prompted the hypotheses that (1) microvascular responses, evidenced by integrin down-regulation and detachment of endothelial cells and astrocytes from ECM, are rapid, simultaneous, and temporally linked to neuron injury, and (2) altered regulation of integrin expression occurs in a heterogenous, but organized fashion. In support of these hypotheses, the authors demonstrate that integrin α1β1 expressed by cerebral microvascular endothelial cells and α6β4 are rapidly down-regulated in response to MCAO in relation to neuron injury, and that the down-regulation of subunit β1 is partially because of shut down of its transcription. In the ischemic basal ganglia, these latter changes are reflected topographically as multiple subregions of microvascular response accompanying ischemic neurons. The current findings have important implications concerning the progressive nature of microvascular injury during experimental stroke.
MATERIALS AND METHODS
All procedures in this study were approved by the Institutional Animal Research Committee and were performed in accordance with the standards published by the National Research Council (The Guide for Care and Use of Laboratory Animals), the National Institutes of Health Policy on Human Care and Use of Laboratory Animals, and the U.S. Department of Agriculture and Animal Welfare Act. In compliance with these standards, every effort was made to ensure that the subjects were free of pain or discomfort. All subjects undergoing experimental procedures were clinically free of apparent infection or inflammation and showed no neurologic deficits. The principal investigator, the primary veterinarian, and primate handling staff were present for all procedures.
Materials from 17 adolescent male baboons (Papio anubis/cynocephalus) were used in this study. The nonhuman primate model of right middle cerebral artery occlusion (MCAO) and reperfusion, as well as the surgical techniques and anesthesia for transorbital implantation of the MCAO device (PS Medical, Goleta, CA, U.S.A.), have been described in detail elsewhere (Abumiya et al., 1999; del Zoppo et al., 1986; Haring et al., 1996b; Okada et al., 1996; Wagner et al., 1997). After implantation of the device, each animal was allowed a 7-day intervention-free period.
Subjects belonged to the following 6 experimental groups: (1) a control group of subjects that did not undergo device implantation surgery (n = 3); (2) subjects undergoing 2-hour MCAO (n = 3); (3) subjects undergoing 3-hour MCAO with 1-hour reperfusion (n = 3), 4-hour reperfusion (n = 3), or 24-hour reperfusion (n = 3); and (4) a separate group of subjects undergoing 3 hours MCAO and 1-hour reperfusion (n = 2). Experiments 1 through 3 were terminated by left ventricular perfusion with an isosmotic nonfixative containing solution (50 g/L bovine serum albumin, 2,000 IU/L heparin, 6.7 μmol/L sodium nitroprusside in Plasmalyte™ chilled to 4°C). In preparation for perfusion, all animals underwent craniotomy under anesthesia with pentothal sodium (15 mg/kg infusion) assisted by mechanical ventilation. Immediately after the perfusion procedure, the brain was quickly excised en bloc from the cranium, immersed in ice, and subdivided by coronal slicing to 1-cm thickness. Tissue blocks (1.0 × 1.0 × 0.5 cm) from stereotactically identical sites from both basal ganglia, parietal and temporal cortices, and cerebellum were embedded in Tissue-Tek OCT compound (Miles, Elkhart, IN, U.S.A.), frozen in 2-methylbutane and dry ice, and stored at −80°C until sectioning. Alternate coronal 1.0 × 1.0 × 0.5 cm tissue blocks were immersion-fixed in 2% paraformaldehyde for 24 hours and then embedded in paraffin for sectioning. The two subjects in the separate group undergoing 3-hour MCAO and 1-hour reperfusion were perfusion-fixed with paraformaldehyde, as described by del Zoppo et al. (1991), to provide tissues for electron-microscopic studies.
Immunohistochemistry
Consecutive 10-μm cryosections from both ischemic and nonischemic basal ganglia, parietal and temporal cortices, and cerebellum were fixed with cold acetone for 10 minutes, then washed. Solubilized milk solids were applied to reduce nonspecific binding of the primary antibody (Johnson et al., 1984). Sections then were incubated with the primary antibody for 2 hours at 37°C, followed by incubation with biotinylated secondary antibody (Vector Laboratories, Burlingame, CA, U.S.A.) for 30 minutes as previously described (Abumiya et al., 1999; Wagner et al., 1997). The avidin-biotin-peroxidase complex technique was used to visualize immunoreactive signals, and 3-amino-9-ethyl carbazole (AEC; Biomeda, Foster, CA, U.S.A.) served as the chromogen. Sections then were counterstained with Mayer's hematoxylin (Biomeda). For dual-label fluorescence immunohistochemistry (IF), 10-μm tissue sections were fixed and incubated with a mixture of a monoclonal antibody and a polyclonal antibody against the desired antigens, followed by incubation with a mixture of fluorescein isothiocyanate (FITC)-conjugated anti-mouse IgG and tetramethyl rhodamine isothiocyanate (TRITC)-conjugated anti-rabbit IgG. Control studies included deletion of the primary antibody, use of an irrelevant primary antibody, and for IF preparations, exchange of the secondary antibody conjugated fluorochromes.
Antibodies
All antibodies used in this study were well characterized (Table 1). Each antibody exhibited immunoreactivity to nonischemic baboon brain microvasculature or to astrocytes, with both peroxidase and fluorescence staining methods (Haring et al., 1996a; Wagner et al., 1997). Optimal working dilutions were established for each antibody preparation against nonhuman primate brain tissues.
Characterization of antibodies
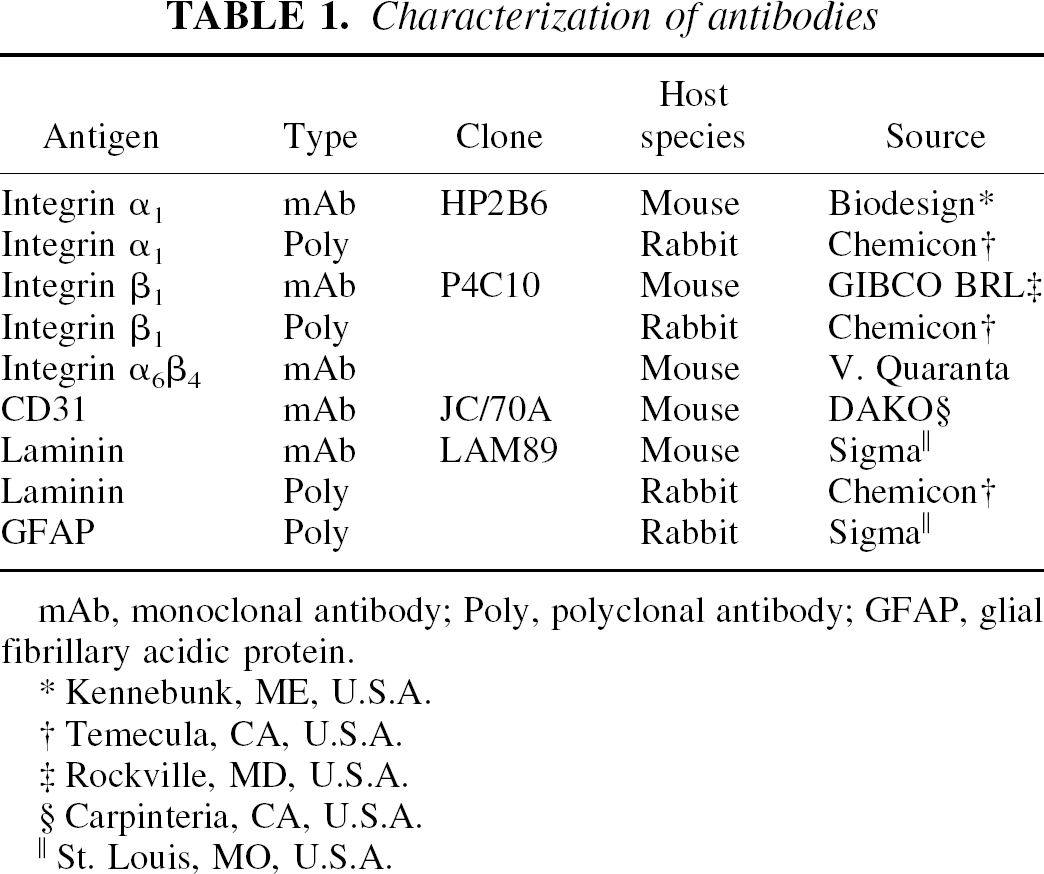
mAb, monoclonal antibody; Poly, polyclonal antibody; GFAP, glial fibrillary acidic protein.
Kennebunk, ME, U.S.A.
Temecula, CA, U.S.A.
Rockville, MD, U.S.A.
Carpinteria, CA, U.S.A.
St. Louis, MO, U.S.A.
Cellular injury
Cells with evidence of DNA damage and repair were detected by in situ incorporation of digoxigenin-dUTP with terminal deoxynucleotidyl transferase (TdT; Boehringer Mannheim, Indianapolis, IN, U.S.A.) or DNA polymerase I (pol I; Promega, Madison, WI, U.S.A.) (Gavrieli et al., 1992; Gold et al., 1993; Schmitz et al., 1991; Tagaya et al., 1997). Both approaches gave nearly identical results (Tagaya et al., 1997). Tissue sections were incubated with TdT (0.3 U/μL) and digoxigenin-11-dUTP (0.5 nmol/L DIG-dUTP; Boehringer Mannheim; Roche, Indianapolis, IN, U.S.A.) in 0.1 mol/L sodium cacodylate (Sigma, St. Louis, MO, U.S.A.), 1 mmol/L CoCl2 (Sigma), 0.1 mmol/L dithiothreitol (Sigma), and 50 μg/mL bovine serum albumin (Sigma) for 60 minutes at 37°C. For experiments with pol I, sections were incubated with a mixture of digoxigenin-dUTP and unlabeled deoxynucleotides, buffer containing 50 mmol/L Tris-HCl (pH 7.5), 10 mmol/L MgSO4, 0.1 mmol/L dithiothreitol, 50 μg/mL bovine serum albumin, and the polymerase for 1 hour at 37°C. Sections then were incubated for 60 minutes with horseradish peroxidase-conjugated antidigoxigenin antibody (Boehringer Mannheim) or rhodamine-conjugated antidigoxigenin antibody (Boehringer Mannheim) to detect incorporated digoxigenin-dUTP. For the immunoperoxidase method, the chromogen AEC was used for color development.
In situ hybridization
35S-UTP cRNA probes were transcribed from linearized templates according to an established protocol (Promega). Antisense RNA probes for integrin β1 were prepared from two cDNA templates (GenBank nr. X07979). The 375 base pair (bp) sequence spanning nucleotides 2390 through 2496 of the translated transmembrane domain, and bp2497 through 2764 including the cytoplasmic tail and a portion of the untranslated region [(5′)GCTAAATTTG…AATCAAGCTT(3′)] was inserted into a pGEM-3Z vector (Promega). A second sequence encompassing 324 bp from bp2440 through 2763 [(5′)AAATCCTAATT…CTTCCTGGAC(3′)] was cloned into a pCR-Script vector (Stratagene, La Jolla, CA, U.S.A.). Antisense RNA probes for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) encompassed 606 bp from bp228 to 833 of the translated region [(5′)AAGCTTGTGA…CGGAGGGCCC(3′)] from the cDNA were cloned into a pGEM-7Zf(+) plasmid (Promega). cDNA sequences for GAPDH and integrin β1 (375 bp) inserted into their respective plasmids were transcribed by SP6 and T7 RNA polymerases, and the integrin β1 (324 bp) cDNA was transcribed by T3 and T7 RNA polymerases to provide sense and antisense RNA probes, respectively. After transcription, the DNA templates were digested with RNase-free DNase, and the RNA probes were extracted, precipitated at −80°C, and resuspended in 20 μL Tris-ethylene diamine tetraacetic acid (EDTA) buffer according to protocol. RNase protection assays were routinely performed using the riboprobes of interest.
In situ hybridization was performed as described in the protocol of Wilcox et al. (1993) with modifications by Seiffert et al. (Levin and del Zoppo, 1994; Seiffert et al., 1991; Seiffert et al., 1994). Sections were deparaffinized in xylene and washed in ethanol. Then the sections were washed in 0.5 × SCC, digested 10 minutes in a 1 μg/mL proteinase K solution, and washed in 0.5 × SCC. Each section was incubated at 42°C for 3 hours with 75 μL prehybridization buffer (50% formamide (v/v), 0.3 mol/L NaCl, 20 mmol/L Tris (pH 8), 5 mmol/L EDTA, 1% Denhardt's solution, 10% dextran sulfate (w/v), 10 mmol/L dithiothreitol) to which 5 × 105 cpm of 35S-labeled probe were added. The sections were incubated overnight at 55°C. Each section then was washed in 2 × sodium chloride/sodium citrate (SSC; 0.15 mol/L NaC1, 0.015 mol/L trisodium citrate, pH 7.0)/2 mmol/L EDTA, digested with 20 μg/mL RNase solution (30 minutes), and washed in 2 × SSC/EDTA. A high stringency wash of 0.1 × SSC/β-mercaptoethanol/EDTA was used for 2 hours at 55°C followed by a 0.5 × SSC wash (4 × 10 minutes). The slides then were dried and exposed on Hyperfilm™ (Amersham, Arlington, IL, U.S.A.) for 3 days at −80°C. To visualize hybridization signals microscopically, the slides were dipped in NTB2 nuclear emulsion (Kodak, New Haven, CT, U.S.A.) then exposed at 4°C for 2 to 4 weeks. After development, slides were counterstained with Richardson or hematoxylin stain. Probes for GAPDH were applied to sections of hippocampus in parallel positive-control experiments.
Quantitative analysis
The number of immunoreactive microvessels and their minimum transverse diameters were determined with the aid of a computerized video-imaging system (VIDAS; Carl Zeiss, Oberkochen, Germany). Analysis was performed within 1.5 mm2 regions of interest defined by 25 nonoverlapping microscopic fields at 400×. The absolute numbers and density (number per unit area) of immunoreactive vessels were further analyzed on the basis of microvascular subclasses previously defined (Haring et al., 1996a; Okada et al., 1994). Immunoreactivities of integrin subunits α1 and β1 also were described in relation to the number of microvessels expressing laminin-1 antigen (α1/laminin or β1/laminin) detectable by the monoclonal antibody LAM-89 (Hamann et al., 1995). No significant differences in laminin expression were observed between basal ganglia of control subjects and the nonischemic basal ganglia of animals subjected to ischemia (data not shown).
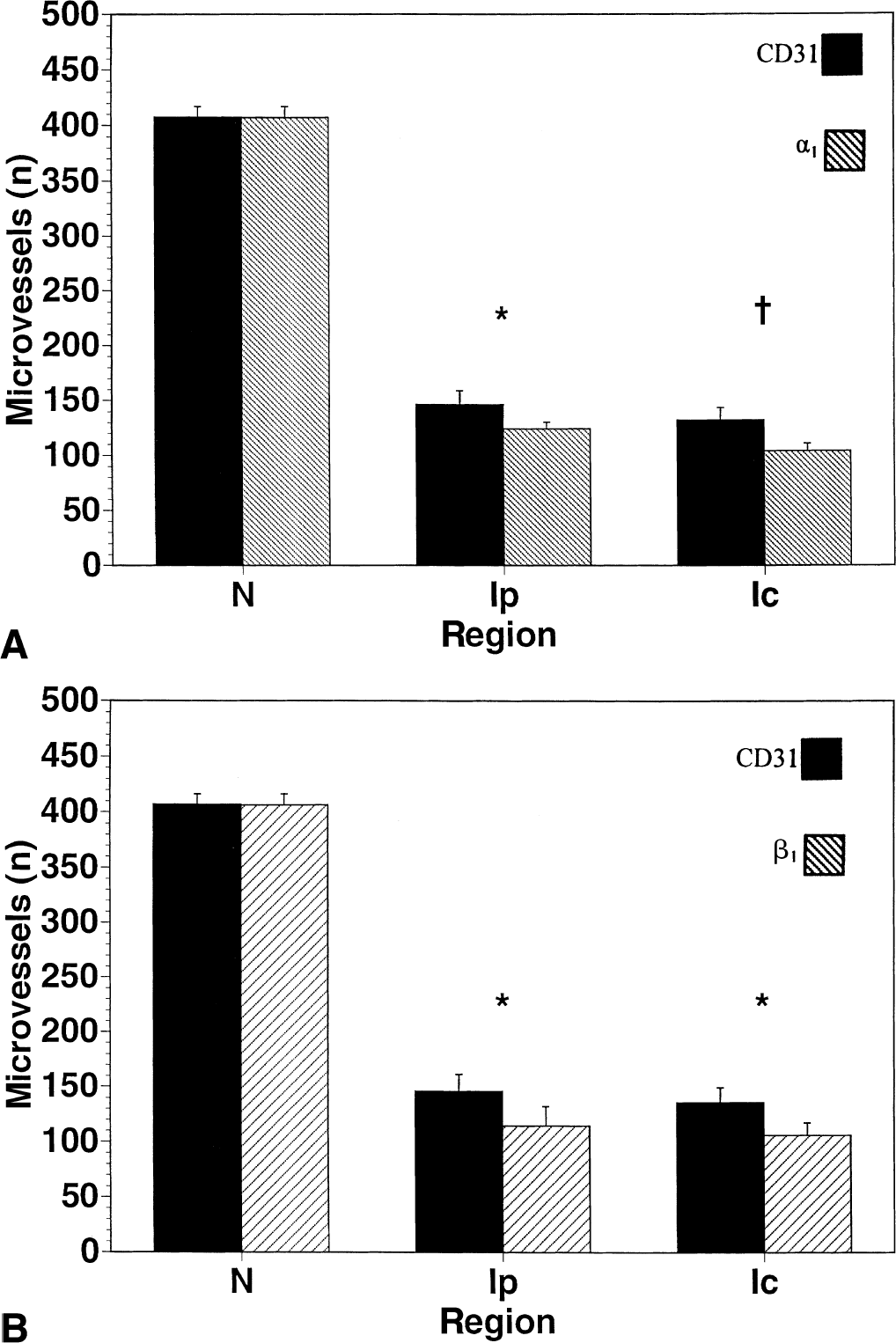
The number of microvessels displaying integrin antigens were correlated with regions displaying cells with dUTP incorporation (dUTP+ cells). At 2 hours of MCAO, 80.0% ± 6.6% of dUTP+ cells are neurons in this model (Tagaya et al., 1997). The number of subunit α1- and β1-immunoreactive and integrin α6β4-immunoreactive microvessels was determined in regions of interest within the dUTP+ region (Ic), in the adjacent dUTP− region (Ip), and in the contralateral nonischemic basal ganglia (N) (Fig. 1).
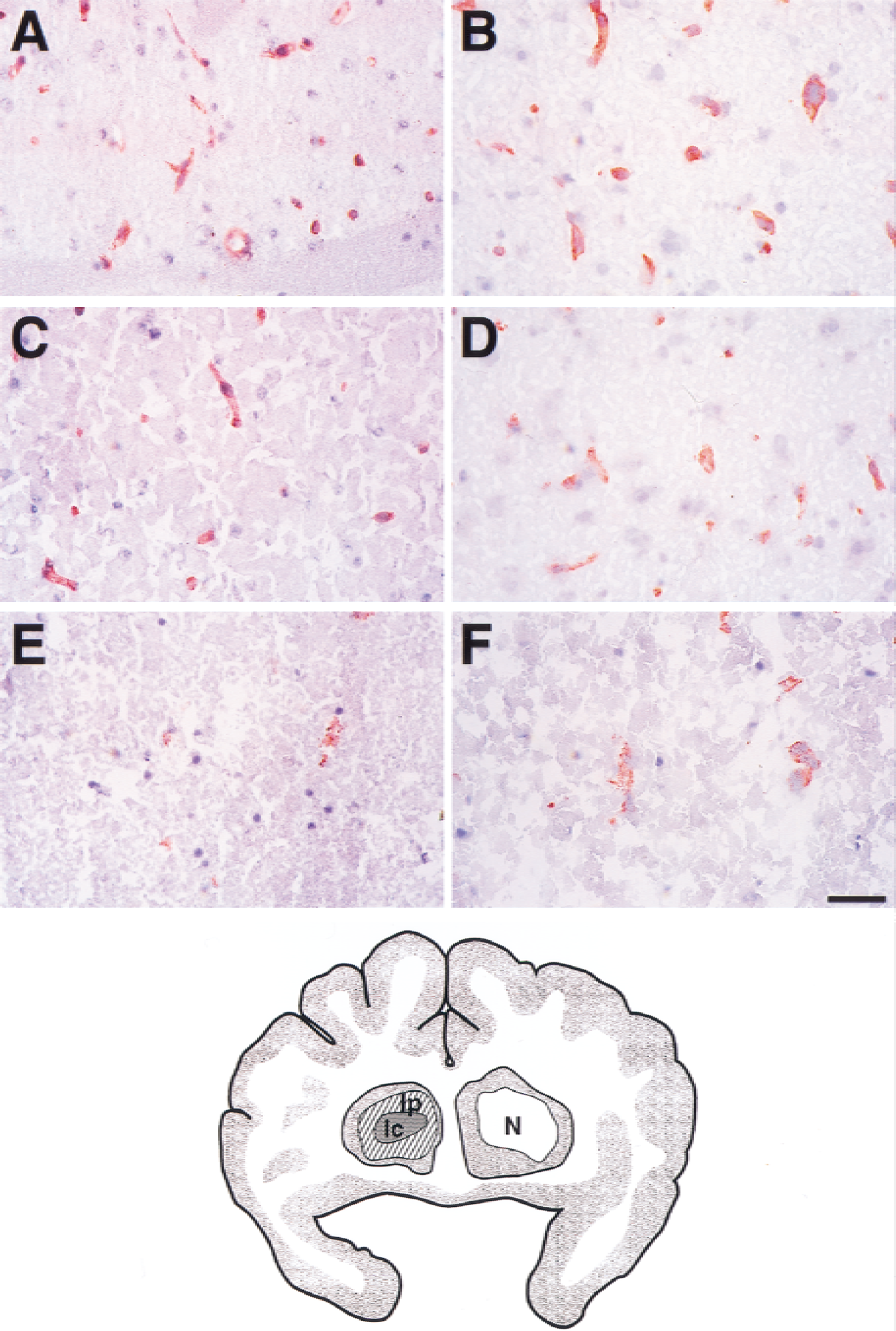
Effect of focal ischemia and reperfusion on integrin α1 and β1 subunit expression in the basal ganglia. α1
Microvessels with 35S granules occupying ≥75% of their circumference were scored as expressing the transcript of interest if the density of 35S signals was above background. The number of microvessels with increased expression was related visually to the regions of dUTP+ incorporation.
Laser-confocal microscopy
Colocalization of vascular antigens was detected by laser-confocal microscopy (LCM) (LSM Invert 410; Carl Zeiss). The relation of integrin subunits α1 or β1 antigens to the endothelial cell marker CD31 was examined using TRITC-and FITC-conjugated secondary antibodies, respectively. Each image was collected at a contrast level of 394 and variable brightness levels using an oil-immersion 100× objective and an internal zoom of 2. For gauging relative integrin expression over astrocyte fibers, immunofluorescence intensities of FITC-labeled anti–glial fibrillary acidic protein antibody were analyzed. Before each measurement, the LCM system was calibrated using slides with defined fluorescence microspheres (Molecular Probes), exposed at 488 nm, with the 100× objective. After calibration, all LCM settings remained unchanged throughout the entire data acquisition process. A region of interest that projected on each glial fibrillary acidic protein–immunoreactive fiber was chosen by an investigator who did not perform the data evaluation. After the optimal confocal plane was determined, the LCM image was captured and translated into its corresponding pixel distribution histogram within a defined range of gray values. On the basis of pixel distribution, the background threshold was determined and subtracted from the overall signals. Subsequently, only pixels representing intensities above that threshold underwent further analysis. The total gray value and the total area covered by three pixels were determined as the integral of those pixels above the background threshold. Finally, the total gray value was normalized to the total area covered by the selected pixels. Intensity data are expressed in arbitrary units.
Electron microscopy
A limited study of microvascular endothelial integrity was performed on specimens prepared as described previously (del Zoppo et al., 1991). Ultrathin sections were cut using a diamond knife on a Reichert Ultracut E, mounted on formvar coated copper grids (150 mesh), and stained with uranyl acetate and lead citrate. The sections were examined on either a Hitachi 12a or a Philips EM208S electron microscope and specific images were documented photographically for further analysis.
Statistical analysis
Data are presented as literal values or as the mean ± SD of multiple experiments. Statistical analyses were performed using two-way analysis of variance or Student's t-test where appropriate. Significance was set at 2 P < 0.05.
RESULTS
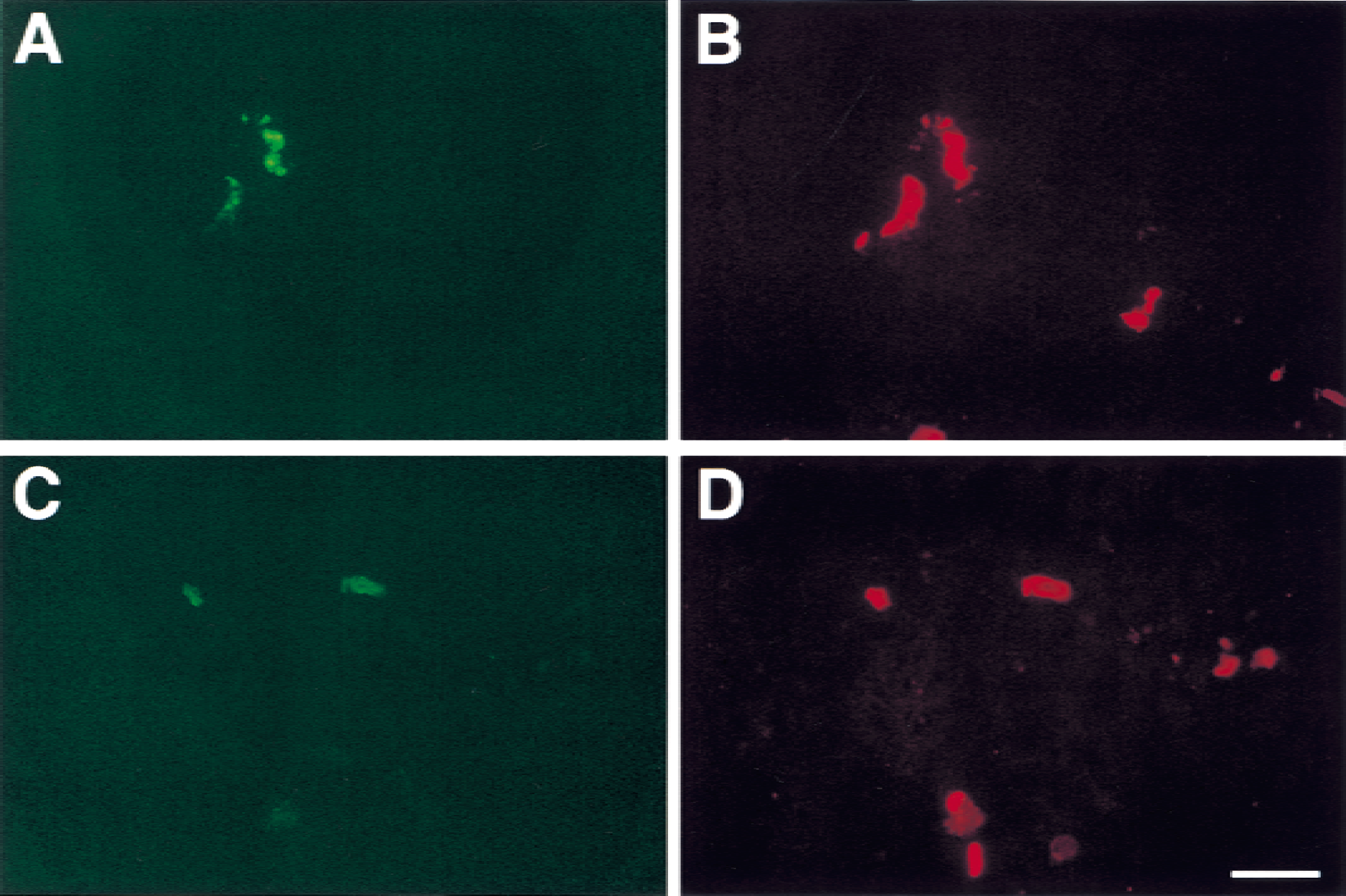
In nonischemic basal ganglia, nearly identical numbers of microvessels displayed integrin subunits α1 and β1, which slightly exceeded the number displaying integrin α6β4 (Fig. 1) (Haring et al., 1996a). Subunit α1 and β1 antigens colocalized with CD31 antigen on microvascular endothelial cells (Fig. 2), but not with vascular smooth muscle cell α-actin (data not shown).
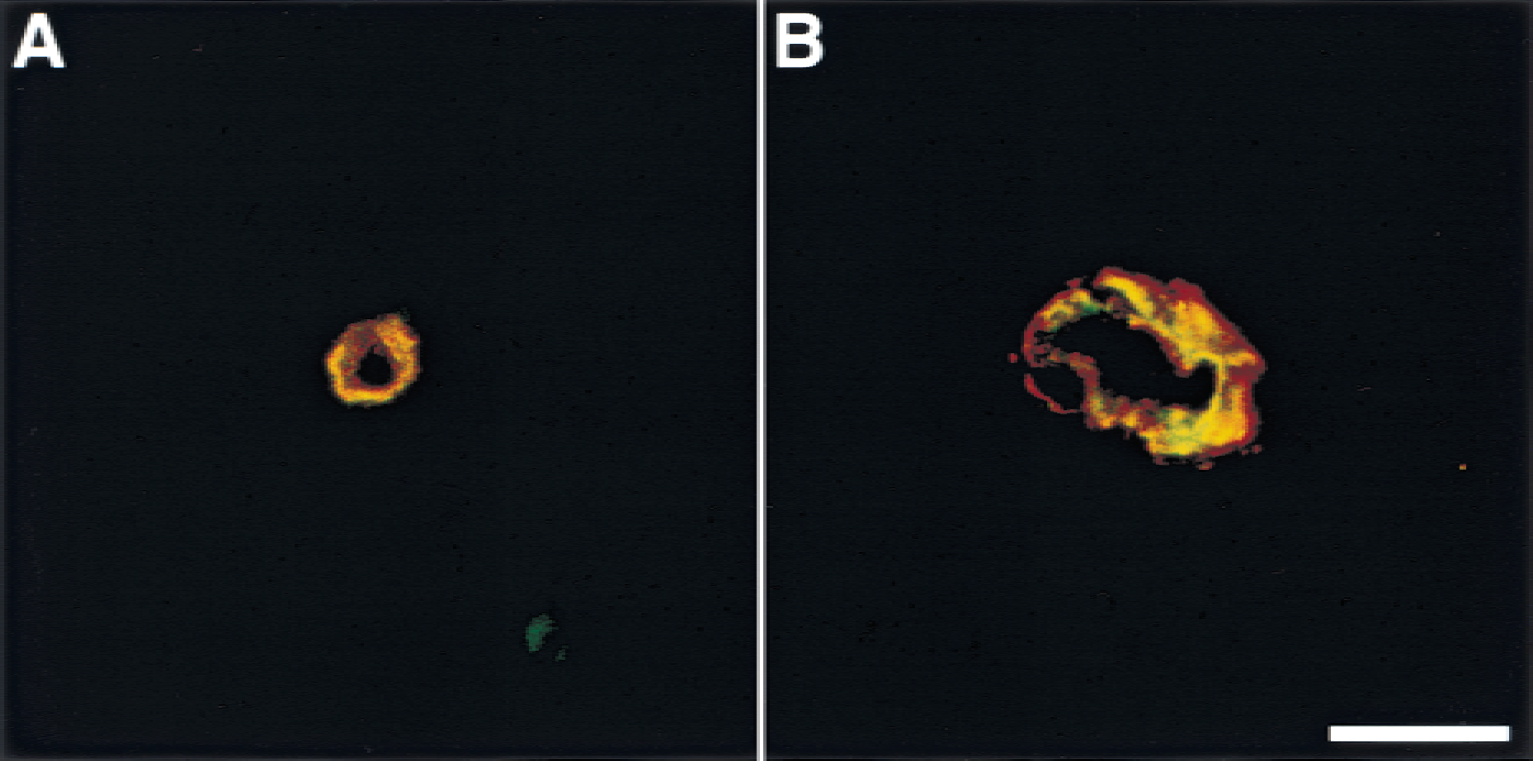
Comparative localization of integrin α1 and β1 antigens and endothelial CD31 by dual immunofluorescence laser confocal microscopy. FITC is green, TRITC is red, and complete colocalization is yellow in the images.
Middle cerebral artery occlusion initiated a significant rapid reduction in the number of subunit α1 and β1-immunoreactive microvessels by 2 hours after MCAO, and this effect was most prominent within the central ischemic region (Ic) (2 P < 0.001 overall) (Figs. 1 and 3). In the neighboring Ip regions, the loss of integrin α1 and β1 antigen expression was less than that seen in the Ic region (Figs. 1 and 3). The mean densities of α1-immunoreactive microvessels decreased from 89.7 ± 1.7/1.5 mm2 (N) to 69.3 ± 3.4/1.5 mm2 in the Ip region and 33.3 ± 5.9/1.5 mm2 in the Ic region (2 P < 0.001 between regions), whereas the mean density of β1-immunoreactive microvessels decreased from 110.7 ± 3.9/1.5 mm2 (N) to 84.0 ± 13.4/1.5 mm2 in the Ip region and 34.0 ± 7.9/1.5 mm2 in the Ic region (2 P < 0.001 between regions) (Fig. 4). In the Ic region, by 24 hours after MCAO, only 22% ± 5% and 21% ± 8% of microvessels expressed subunit α1 and β1 antigens, respectively.
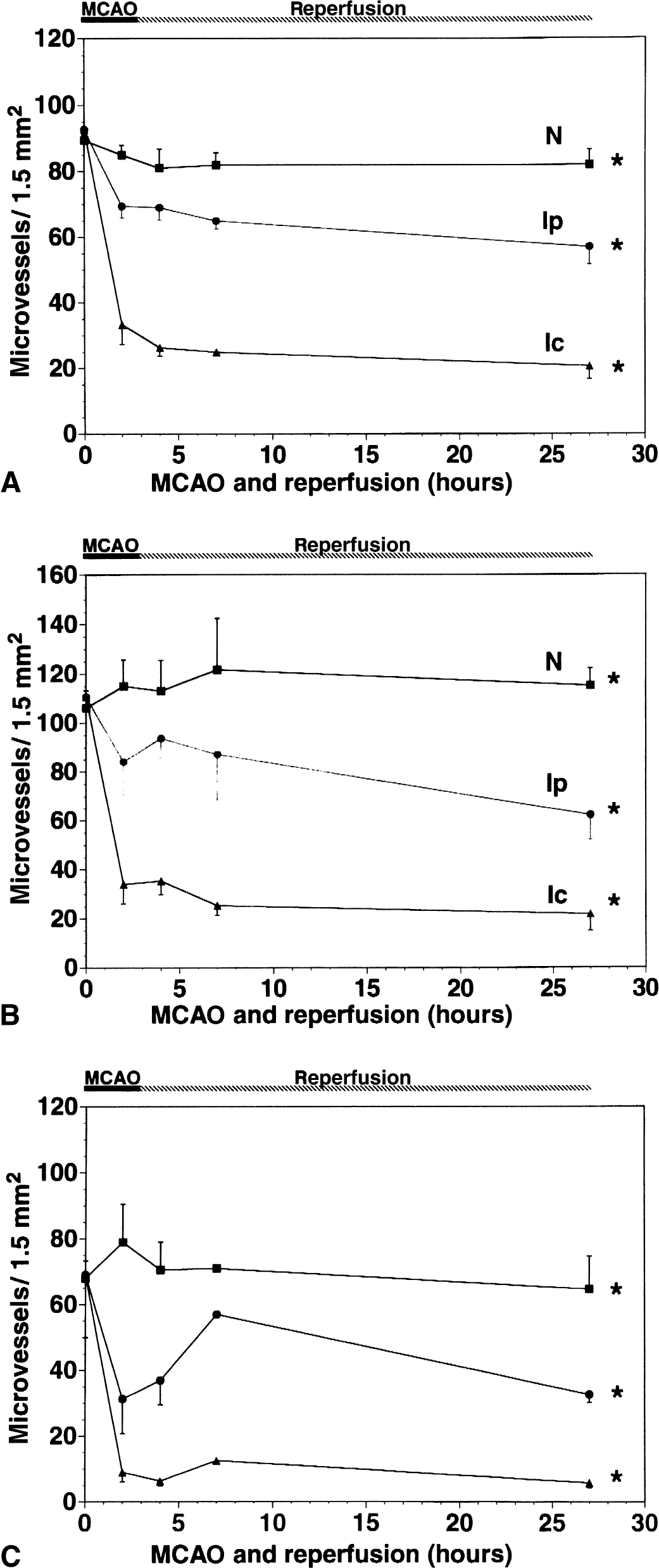
Temporal profile of changes in the number of microvessels displaying integrin subunit α1
Responses of microvessels displaying CD31 and integrin α1-immunoreactivity
To determine whether the loss of integrin subunits α1 and β1 might be related to other alterations exhibited by endothelial cells, dual immunofluorescence histochemistry experiments compared their expression to that of endothelial cell CD31 and laminin-1 in the basal lamina. The number of microvessels displaying endothelial cell CD31 antigen decreased significantly in the Ip and the Ic regions by 2 hours MCAO (2 P < 0.05, each comparison), which was marked by a significant further relative reduction in the number of α1- and β1-immunoreactive microvessels (Fig. 4). However, all microvessels that retained expression of α1 and β1 antigens also expressed CD31. Detectable expression of integrin α1β1 was found only on CD31+ endothelial cells in microvessels with intact basal lamina. A similar correlation could be made for laminin-1 expression as those microvessels devoid of α1 and β1 expression also exhibited a loss of laminin-1 expression. The decrease in subunit α1 and β1 antigen expression in both the Ic and Ip regions exceeded the simultaneous loss in laminin-1 immunoreactive microvessels (2 P < 0.001, data not shown). Figure 5 demonstrates the loss of α1- and β1-immunoreactive microvessels relative to the expression of laminin in microvessels within the Ic region. A separate electron microscopy survey demonstrated that all microvessels in the ischemic zone by 4 hours after MCAO (n = 2) had visibly intact endothelial cells, equivalent in number and distribution to the contralateral basal ganglia and temporal cortex (data not shown).
Extended loss of integrin subunit α1 and β1 antigens compared with that of laminin expression in the ischemic basal ganglia.
Astrocyte fibers around select microvessels displayed both subunit α1 and β1 antigens in the nonischemic cerebral tissues (Haring et al., 1996a). However, in the Ic region, relative FITC-intensity measurements of astrocyte fibers revealed a significant loss in fiber-associated β1 immunoreactivity, from 23.8 ± 8.9 units to 5.8 ± 1.3 units as early as at 2 hours of MCAO (2 P < 0.001) (Fig. 6), which coincided with the loss of microvascular subunit α1 and β1 antigens. At 24 hours of reperfusion after 3-hour MCAO, the FITC signal intensity had decreased further to 3.3 ± 0.76 (2 P < 0.005).
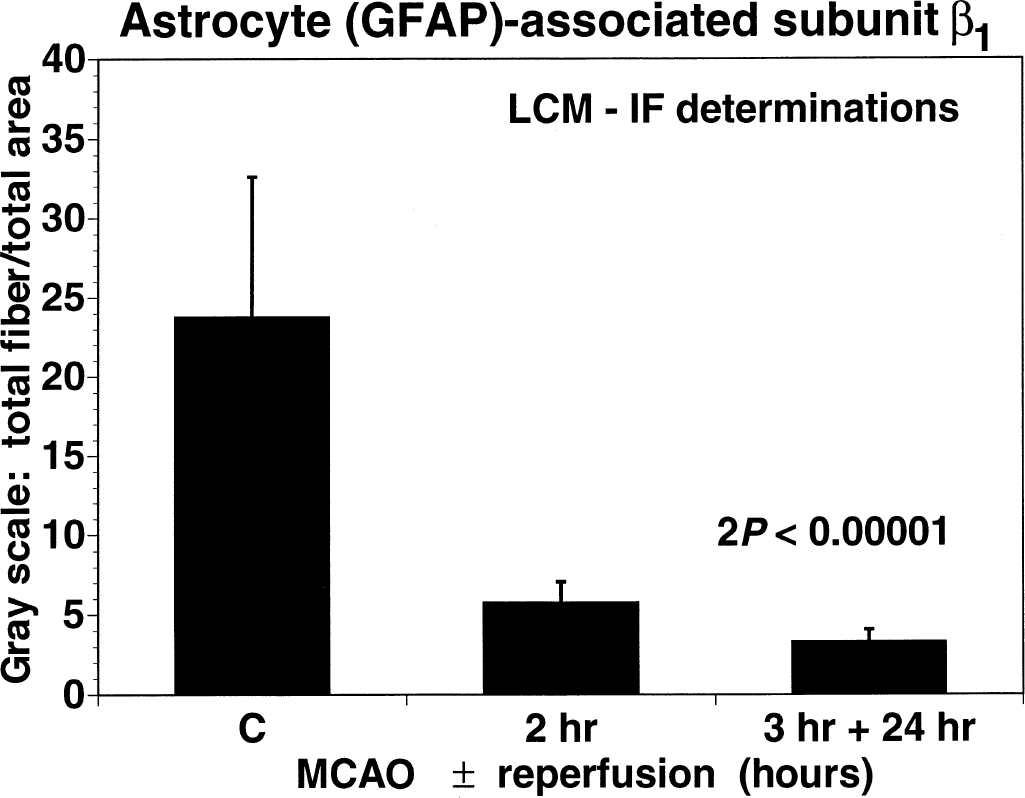
Changes in fluorescence intensity of integrin β1 immunoreactive astrocyte fibers (FITC) around selected microvessels in response to middle cerebral artery occlusion (MCAO) at 2 hours (2 P < 0.0001) and 24 hours of reperfusion after 3-hour MCAO (2 P < 0.005). Values are expressed in arbitrary units (see text). GFAP, glial fibrillary acidic protein; LCM, laser-confocal microscopy; IF, fluorescence immunohistochemistry.
Integrin α6β4 expression has been shown to colocalize with astrocyte endfeet in uninjured brain tissue (Wagner et al., 1997) and to decrease after MCAO. Coinciding with the loss of integrin α1β1 antigens, the mean density of α6β4-immunoreactive microvessels decreased from 79.0 ± 11.5/1.5 mm2 (N) to 31.3 ± 10.5/1.5mm2 in the Ip region and 9.0 ± 3.0/1.5mm2 in the Ic region by 2 hours of MCAO (2 P < 0.001) (Fig. 3). This coincided with astrocyte endfoot swelling observed by 4 hours of MCAO (data not shown). By 24 hours, residual integrin α6β4 expression was confined to 8.8% of microvessels.
To determine whether the ischemia-related loss in α1 and β1 antigen expression might reflect a change in transcription, in situ hybridization experiments with two β1 antisense RNA probes were performed in both nonischemic control and 2-hour MCAO subjects. Both β1 probes gave identical results. Sense probes displayed no obvious cell or microvessel-associated signals in matched tissue sections. At 2-hour MCAO within the Ic region, subregions containing no subunit β1 mRNA expression on any microvessel or cell were surrounded by groups of microvessels with β1 mRNA hybridization signals (Figs. 7 and 8). Subunit β1 mRNA signals were found on fewer microvessels in the contralateral nonischemic basal ganglia of ischemic subjects and the basal ganglia of control subjects (Fig. 7).
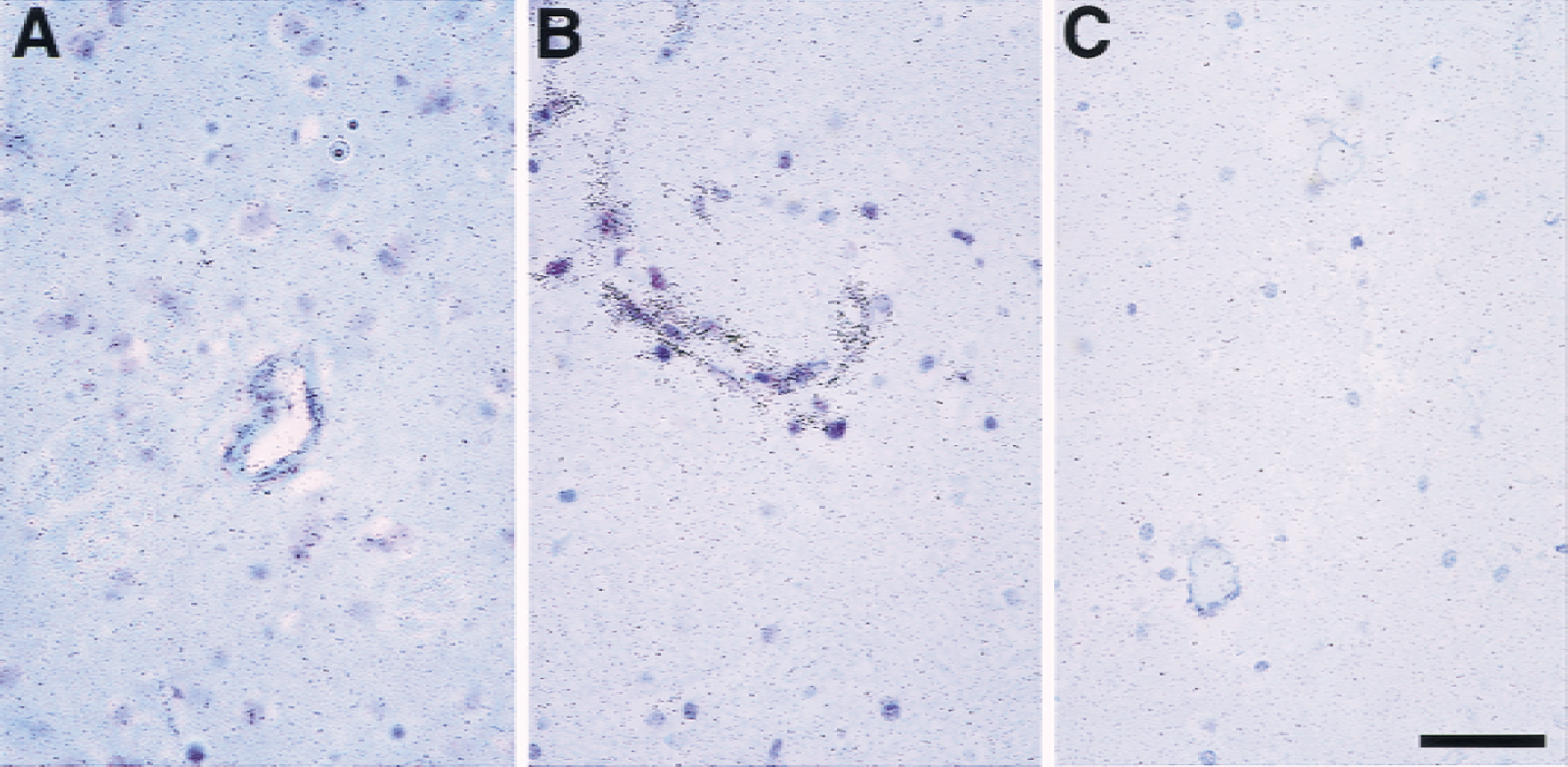
Representative photomicrographs of in situ hybridization for integrin β1 mRNA.
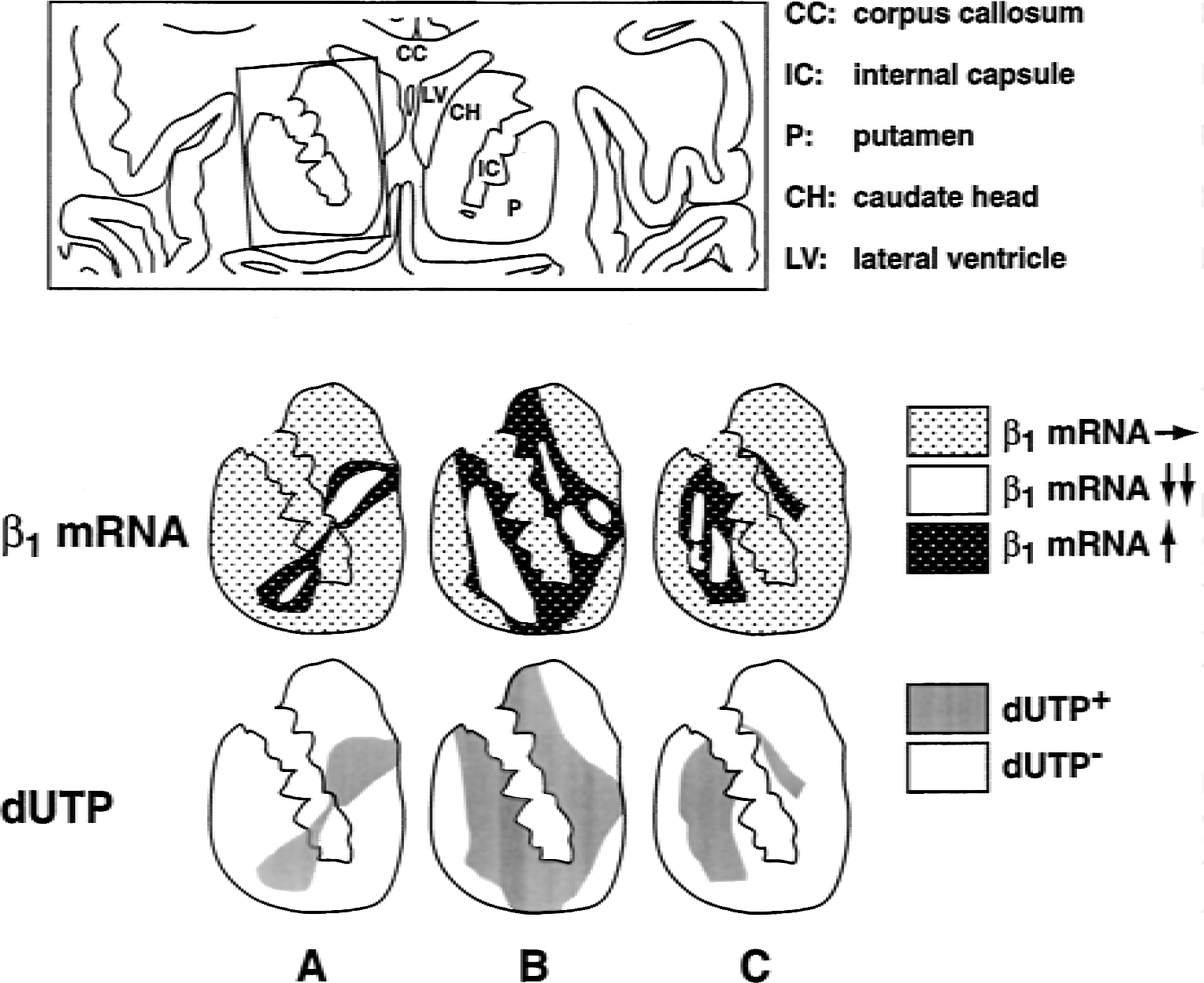
Patterns of β1 mRNA expression compared with the distribution of neuronal cell injury detected as cellular dUTP incorporation in the basal ganglia at 2-hour MCAO in each subject (n = 3) (A to C). Areas of reduced β1 expression, enhanced expression of β1, and normal β1 expression are depicted. Nonischemic contralateral basal ganglia and both basal ganglia of control subjects (n = 3) displayed scattered microvessels with β1 mRNA expression and ≥1 cells with dUTP incorporation (not shown). Regions of cell dUTP incorporation are displayed in relation to β1 mRNA expression
In the Ic region, at the dUTP+/dUTP− boundary, a significant increase in the number of microvessels displaying β1 transcripts and in the microvessel-associated β1 mRNA signal intensity was also seen (Fig. 7). At 2 hours of MCAO, 39.3% ± 3.0% microvessels displaying collagen IV antigen at the Ic/Ip boundary also displayed increased β1 mRNA (n = 3 subjects). The microvessels that exhibited increased β1 mRNA levels were 9.8 ± 0.8 μm in diameter, as compared with a mean diameter of 6.2 ± 0.7 μm for all microvessels in the ischemic basal ganglia. This reflects the observation that only cells in noncapillary microvessels showed increased β1 mRNA in the Ic/Ip boundary. The β1 35S signals were prominent over the vascular media, although the exact cell associations could not be identified. Despite low frequency nonvascular 35S signals in the nonischemic or ischemic regions, localization of increased β1 mRNA within astrocytes at any time point could not be ruled out.
At 2 hours of MCAO, the patterns of microvascular β1 mRNA expression were closely associated with the region of neuronal dUTP incorporation in the ischemic basal ganglia in all subjects (Fig. 8). Up-regulation of microvascular β1 mRNA was seen predominantly in the dUTP+ gray matter of the basal ganglia. In all subjects, despite a general reduction in the number of microvessels displaying subunit β1 antigen, multiple islands absent of vascular β1 mRNA signal were surrounded by borders exhibiting increased β1 transcription, all continuous and completely confined within the dUTP+ region (Fig. 8).
DISCUSSION
Cerebral microvascular endothelial cells are the first to witness alterations in blood flow and changes in oxygen and nutrient supply to the brain. The intact endothelium, through the blood–brain barrier and the subjacent basal lamina and ECM, prevents contact between plasma constituents and the neuropil. Unlike other β1 integrins, integrin α1β1 has been found on all microvessels (Gehlsen et al., 1989; Hall et al., 1990; Korhonen et al., 1990; Kramer et al., 1990) and now on cerebral microvessels including capillaries (Haring et al., 1996a; Paulus et al., 1993). Both integrins α6β4 and α1β1 are associated with astrocyte endfeet and fibers, respectively (Wagner et al., 1997). Although ischemia is known to alter microvascular blood–brain barrier and basal lamina integrity, its effects on cerebral microvascular cell-matrix adhesion are not known. This is the first article to demonstrate the coordinated and altered appearance of integrins expressed by cerebral endothelial cells and astrocytes during focal cerebral ischemia. In the current experiments, MCAO produced a rapid simultaneous reduction in the number of microvessels displaying integrins α1β1 and α6β4 in the central ischemic region of neuron injury (dUTP+, Ic) within the basal ganglia, which exceeded that of the peripheral dUTP− region (Ip), and non-ischemic zones (N). These findings indicate that ischemia affects events on both sides of the microvessel basal lamina matrix simultaneously and with equal speed.
Furthermore, the presence of a central core of injury manifested by the near absence of endothelial cell integrin α1β1, and astrocyte integrin α1β1 and α6β4 antigens surrounded by a peripheral region of reduced injury is topographically consistent with current notions of regional cerebral blood flow reduction and associated metabolic alterations after MCAO (Astrup et al., 1981; Furlan et al., 1996; Jones et al., 1981). The persistent decrease in integrin expression reflects injury to the endothelial cells and astrocytes, alterations of the matrix by processes initiated by ischemia within microvascular endothelium or astrocytes, or both. The presence of β1 integrin up-regulation confirms the presence of viable microvascular cells within the Ic. However, increased microvascular β1 mRNA was presented in multiple circular arrays around subregions devoid of β1 antigen or transcript appearance. These changes indicate that, despite widespread loss of integrin antigens, injury was not complete. This contrasts with the commonly held view that striatal injury after MCAO is complete and irreversible.
Under the circumstances of these experiments, loss of integrin expression could reflect the contributions of several processes. Conformational changes in either α or β integrin subunits or their proteolysis could alter their antigenicity and disengage endothelial cells or astrocytes from matrix ligands (Luque et al., 1994). Although shedding of the extracellular domains of certain endothelial cell adhesion receptors is known to generate soluble fragments (Fassbender et al., 1995), to the authors' knowledge, no evidence currently exists that proteolysis alone causes loss of α1 and β1 in this manner.
Loss of integrins α1β1 and α6β4 may reflect changes in the major basal lamina and ECM ligands after MCAO (Hamann et al., 1995; Wagner et al., 1997). Plasmin and metalloproteinase-2 cleave laminin-1 and collagen IV, leading to degradation of the vascular matrix (Krane, 1994). The loss of vascular integrin expression was greater than the loss of their respective matrix ligands (Hamann et al., 1995) and may reflect greater sensitivity of integrins α1β1 and α6β4 to ischemia. Alternatively, given the multiplicity of ligand antigenic sites, a change in laminin-1 structure undetectable by immunohistochemical methods could contribute to the changes in α1β1 expression.
Hossmann et al. and others have suggested that a general regional shutdown in protein synthesis occurs during focal cerebral ischemia (Bodsch et al., 1986; Erdogdu et al., 1993; Kleihues and Hossmann, 1971) and experimental global cerebral ischemia (Kranse and Tiffany, 1993; Widmann et al., 1991). The latter is partly explained by selective impairment of nuclear-cytoplasmic transport of mRNA, so that while total RNA synthesis is normal during reperfusion, individual mRNA species may be differentially lost (Matsumoto et al., 1990; Sakaguchi et al., 1988). Although unproven, the presence of residual flow, as during focal cerebral ischemia, may modulate these changes. The early appearance of the endothelial cell-leukocyte adhesion receptors ICAM-1 and E-selectin (Haring et al., 1996b; Okada et al., 1994), integrin αvβ3 on microvascular smooth muscle cells (Abumiya et al., 1999; Okada et al., 1995), and latent metalloproteinase-2 (Heo et al., 1999) after MCAO indicate that local shutdown of microvascular cell protein synthesis is selective. Moreover, within the Ic region, microvascular α1β1 expression was not completely lost (Fig. 3). This residual expression is consistent with continued synthesis of integrin α1β1 antigen on selected microvessels within both the Ic and Ip regions (Ip > Ic). The less prominent down-regulation of CD31 antigen also supports a selective modulation of endothelial cell protein synthesis. Finally, preservation of integrin α1β1 expression coincided with microvessels expressing laminin-1 antigen. Previously, it had been shown that up-regulation of E-selectin occurs only in those vessels with intact basal lamina (Haring et al., 1996b). This implies that matrix integrity is required for cerebral endothelial cell integrin expression and viability.
Dynamic changes in endothelial cell-matrix interactions are known to result from down-regulation of integrin receptors of the β1 subfamily by the cytokines TNF-α and IL-1β (Defilippi et al., 1992). In one study, stimulation of endothelial cells by TNF-α for 24 hours resulted in a 60% loss of integrin α6β1 expression from baseline and significantly decreased endothelial cell adhesion to laminin matrix (Defilippi et al., 1992). Integrin α6β1 recognizes a different binding site on laminin-1 than integrin α1β1(Hall et al., 1990; Kramer et al., 1990; Sonnenberg et al., 1990). This mechanism is relevant as TNF-α and IL-1β are generated in ischemic cortex by microglia and astrocytes during MCAO (Buttini et al., 1996; Liu et al., 1993; Loddick and Rothwell, 1996). In addition, IL-1β may participate in early postischemic brain edema formation (Yamasaki et al., 1992). In rat skin, functional blockade of the integrin β1 subunit causes edema formation by inhibiting cell-matrix adhesion and decreasing interstitial fluid pressure (Reed et al., 1992; Rodt et al., 1994). Hence, β1 integrins, through their participation in endothelial cell and astrocyte-matrix interactions, may play a role in preventing edema formation together with the intact blood–brain barrier. In addition, growth and differentiation factors can regulate integrin expression in selected cells (Smyth et al., 1993). For instance, TGF-β enhances expression of subunits β1, β2, and β3in vitro (Heino et al., 1989; Ignotz and Massague, 1987). Cytokine generation during ischemia may promote the disintegration of endothelial cell-matrix interactions, whereas a gradient of cytokine generation that decreases from the Ic to the Ip region could explain the greater subunit β1 expression in the Ip region. These considerations suggest that cytokine or growth factor production and residual flow during focal ischemia could contribute to the concentric up-regulation of microvascular β1 mRNA within the Ic region and at the Ic/Ip boundary.
The complex but ordered pattern of microvascular transcriptional response in β1, multiple contiguous concentric borders around subregions of absent β1 mRNA, suggests that focal ischemia alters endothelial cell integrin expression and integrity in groups or arrays of microvessels that are related anatomically. The location and pattern of increased β1 transcription may also bear on the nature of the ‘ischemic penumbra’ in the striatum. The concept of a peripheral ‘penumbra’ region of ischemic injury, characterized by critically reduced CBF, preserved neuronal structure, and potentially reversible electrical failure without membrane or metabolic failure, has assumed a single core of irreversible damage (Furlan et al., 1996; Heiss, 1992; Jones et al., 1981). In contrast, these experiments suggest that multiple cores of irreversible microvascular cell injury, rather than a single core, develop early (by 2 hours) within the ischemic basal ganglia after MCAO. The ischemic ‘penumbra’ has also been defined by the duration of CBF reduction in the ischemic zone (Astrup et al., 1981). The multiple subregions of β1 mRNA down-regulation surrounded by borders of microvascular β1 mRNA up-regulation within the region of severe neuron injury may reflect reduced CBF or varied critical thresholds of different cell populations or microvessels to local ischemia. Hence, cells or microvessels surrounding the ischemic core may be incorporated into the tissues of irreversible injury in a time-dependent manner (Furlan et al., 1996). In this concept, microvessels displaying increased β1 mRNA within the Ic/Ip boundary would be at risk for further injury, so that the subregions of β1 mRNA loss (Fig. 8) ultimately merge together into larger regions during the developing infarct. Within the basal ganglia, the overall size of injured tissue is already fixed by 2-hour MCAO (Tagaya et al., 1997). This heterogeneity of microvascular response is consistent with acute observations made by positron emission tomography in select ischemic stroke patients (Young et al., 1997).
β1 integrins play important roles in maintaining cell viability. Disruption of interactions between subunit β1 and ECM proteins are reported to induce apoptosis in CID-9 epithelial cells (Boudreau et al., 1995), lung endothelial cells (Hoyt et al., 1996), and colonic crypt cells (Strater et al., 1996). In epithelial cells, apoptotic cell death appears controlled through an integrin-dependent negative regulation of interleukin-1β converting enzyme expression (Boudreau et al., 1995). Colocalization experiments performed here also suggest that microvascular endothelial cell viability is compromised early after MCAO. By 2-hour MCAO, a 67% reduction in microvascular CD31 antigen expression and a significant 74% reduction of α1 and β1 immunoreactivity occurred in the Ic region (Fig. 5). Despite the loss of CD31, α1, and β1 antigen expression could not be attributed to obvious endothelial cell detachment, confirming an earlier study (del Zoppo et al., 1986). Furthermore, it does not appear that apoptosis is a substantial contributor to loss of endothelial cell viability, despite the loss of integrin α1β1 expression, because dUTP incorporation was observed in less than 2.1% of microvessels by 24 hours after MCAO (Tagaya et al., 1997).
Products of ischemic injury also may alter endothelial cell integrin β1 expression. Bradley et al. (1995) have shown that exposure of endothelial cells in vitro to a source of free radicals disrupts endothelial cell-matrix interactions by reversibly altering the abluminal distribution of subunit β1. In contrast, the reduction in microvascular endothelial cell integrin α1/β1 expression in the current study was not increased in either the Ic or Ip regions by reperfusion of the MCA, which was consistent with irreversible shutdown of subunit α1 and β1 antigen expression by processes initiated before 2 hours of MCAO. It is of interest whether reperfusion after shorter periods of MCAO (for example, 1 hour) may recover endothelial cell integrin α1 or β1 or astrocyte α6β4 antigen expression in these regions.
In conclusion, because the interaction of endothelial integrin α1β1 and astrocyte integrin α6β4 with their basal lamina ligands contributes to maintaining microvascular integrity under normal conditions, the simultaneous loss of integrins α1β1 or α6β4 in concert with loss of matrix components may reflect decreased endothelial cell viability. The loss may also lead to the development of vasogenic edema, or hemorrhagic transformation in the central ischemic zone, or both (Hamann et al., 1996). A provocative notion is that neuron viability may be affected by the loss of endothelial cell α1β1, or astrocyte α6β4, or both—that is, neuron and microvascular injury may be directly linked. Support for this comes from the nearly identical temporal and topographical changes in both integrins in response to MCAO in these experiments. Neuron-associated integrin expression in other species and in cell culture has been reported (Albelda et al., 1989; Einheber et al., 1996; Nishimura et al., 1998; Paulus et al., 1993), but their relation to microvessel viability or matrix-ligand expression has not been described. Within the central ischemic zone where microvascular integrin α1β1 and α6β4 antigens decrease, up-regulation of integrin β1 mRNA expression occurs, indicating selective microvessel viability. An important implication of this observation is that early restoration of blood flow to the ischemic basal ganglia where microvascular injury is heterogeneous may preserve microvascular and neuronal viability within the ischemic core, as well as the peripheral zones. The current studies demonstrate an inextricable link between cerebral microvessel integrin expression and neuron injury.
Footnotes
Acknowledgments:
The authors are grateful to John Hallenbeck, M.D., for his suggestions on the late stages of this manuscript. The authors also thank Malcolm Wood, Ph.D., for performing the EM studies, and Marcia Filbert and Diane Posvar for their expert assistance.