
Abstract
To investigate whether rat hippocampal neurogenesis varies with strain and gender, the authors examined proliferating progenitor cells and their progeny in young male and female Sprague-Dawley (SD) and spontaneously hypertensive rats (SHR) using the thymidine analog bromodeoxyuridine (BrdU) combined with immunohistochemistry for the neuronal marker Calbindin D28k and glial fibrillary acidic protein. Rats were given 7 consecutive daily BrdU injections and were killed 1 day or 4 weeks later to allow for discrimination between proliferation and cell survival. Stereologic analysis of the numbers of BrdU-immunoreactive cells in the dentate gyrus revealed both a strain difference with significantly higher cell proliferation and net neurogenesis in SHR than in SD and a gender difference with males from both strains producing significantly more cells than their female counterparts. Whereas the number of progenitors four weeks after BrdU injections was still significantly greater in male than in female SHRs, resulting in a greater net neurogenesis in the male, the number of BrdU-immunoreactive cells did not differ between male and female SD rats, suggesting a greater survival of newly generated cells in the dentate gyrus in female than in male SD rats. No sex or strain difference was observed in the relative ratio of neurogenesis and gliogenesis.
Postnatal neurogenesis in the granule cell layer of the hippocampal formation in the rat, first described by Altman and Das (1965), has been shown to persist well into adulthood in the rodent (Kaplan and Hinds, 1977; Bayer et al., 1982; Kaplan and Bell, 1984; Kempermann et al., 1998b). Furthermore, neurogenesis has been demonstrated to occur in vivo in the adult human dentate gyrus (Eriksson et al., 1998). Neural stem-like cells reside in the subgranular zone at the border of the granule cell layer and the hilus of the dentate gyrus where they continuously proliferate, migrate into the granule cell layer, and differentiate into granule cells (Cameron et al., 1993). Newly generated granule cells express markers of differentiated neurons and have phenotypic characteristics corresponding to differentiated granule cells including axonal processes into the mossy fiber pathway with synaptic connections to their targets in hippocampus CA3 (Stanfield and Trice, 1988; Cameron et al., 1993).
Differentiation of multipotent neural stem cells is determined by a complex interaction between intrinsic programs and external cues such as neurotrophins, hormones, stress, and brain injury (Snyder, 1998, Gould and Tanapat, 1999; Shihabuddin et al., 1999). Marked strain differences in neural stem cell proliferation, differentiation, and survival have been observed in female mice (Kemperman et al., 1997a).
As a background to ongoing studies on lesion-induced neurogenesis, the aim of the current study was to analyze whether proliferation of progenitor cells and the subsequent generation of new neurons within the dentate gyrus differ between female and male rats in spontaneously hypertensive rats (SHR) and Sprague-Dawley (SD) rats, two strains commonly used in experimental stroke research.
MATERIALS AND METHODS
Animals
Four-week-old male and female SHR and SD were obtained from Möllegaard Breeding Center (Ejby, Denmark). Animals were transported under similar conditions and were housed either 3 or 4 rats per cage and maintained on a 12/12 hour light/dark cycle (lights on at 7:00 am) and provided ad libitum access to food and water. Body weight was recorded daily during the first two weeks, then weekly. Animals were allowed to settle for one week in a limited access animal facility. Animals were housed for either one or five weeks in standard housing.
Progenitor proliferation was assessed by labeling dividing neurons using systemic administration of the thymidine analog bromodeoxyuridine (BrdU), which is incorporated into the DNA of dividing cells during the S phase of the mitotic cycle. Survival of cells was estimated by comparing the number of BrdU immunoreactive cells 1 day and 4 weeks after the last injection of BrdU. All cell counts were performed using unbiased stereologic counting techniques (Coggeshal and Lekan, 1996; Gundersen et al., 1988). To determine the phenotypic cell fate of the cells that underwent mitosis during BrdU administration, the authors performed immunohistochemical characterization one month after BrdU labeling using cell-specific markers, including glial fibrillary acidic protein (GFAP, a marker for astroglia) and Calbindin D28k (Sloviter, 1989) (a marker for granule cells in the dentate gyrus).
Experiment 1
To examine whether a sex or strain difference, or both, exist in the number of proliferating hippocampal cells, 5-week-old male and female rats of both strains received daily injections intraperitoneally of BrdU (50 mg/kg body weight) during 7 consecutive days. BrdU was dissolved in 0.9% NaCl and filtered at 22 μm. The thymidine analog BrdU is incorporated into the genetic material upon mitotic division, after which it can be detected immunohistochemically in the daughter cells (del Rio and Soriano, 1989; Nowakowski et al., 1989; Kuhn et al., 1996).
One day after the last BrdU injection, groups of female and male rats from both strains (n = 8 in each group) were killed and perfused. Brains were removed, postfixed, and processed for BrdU immunolabeling.
Experiment 2
To determine whether a sex or strain difference, or both, exist concerning the survival of progenitor cell progeny in the dentate gyrus, groups of female and male rats (n = 5 in each group) from both strains were killed and perfused 4 weeks after the last BrdU injection.
Perfusion
All animals were deeply anesthetized with an overdose of sodium pentobarbital and then perfused transcardially with saline solution followed by 4.0% paraformaldehyde in 0.1 mol/L PBS. After perfusion, the brains were removed, postfixed, and processed for BrdU immunolabeling or for combined immunofluorescence staining for BrdU, GFAP, and Calbindin D28k.
Histologic procedures
After perfusion and removal, the brains were postfixed overnight in 4% paraformaldehyde in 0.1 mol/L phosphate buffer, and thereafter transferred into 30% sucrose. Each brain was sectioned coronary (40 μm) trough the entire hippocampus on a sliding microtome. Sections were stored individually at −20°C in a cryoprotecting buffer containing 25% ethylene glycol, 25% glycerin, and 0.05 mol/L phosphate buffer until they were processed for immunohistochemistry or immunofluorescence.
Tissue preparation for BrdU immunohistochemistry
To visualize BrdU immunoreactivity, DNA has to be denatured. Sections were incubated in 50% formamide/50% 2 × SSC buffer (0.3 mol/L NaCl/0.03 mol/L sodium citrate) at 65°C for 2 hours, rinsed in 2 × SSC, incubated in 2 N HCl for 30 minutes at 37 °C, and rinsed in 0.1 mol/L borate buffer (pH 8.5) for 15 minutes.
Immunohistochemistry
For peroxidase BrdU immunolabeling, free-floating sections were treated with 0.6% H2O2 in tris-buffered saline (TBS) (0.15 mol/L NaCl, 0.1 mol/L Tris-HCl, pH 7.5) for 30 minutes to block endogenous peroxidase. After pretreatment for BrdU immunohistochemistry (see above), sections were rinsed several times in TBS followed by incubation in TBS/0.1% Triton X-100/3% horse serum (TBS-TS) for 30 minutes and incubation with primary anti-BrdU antibody (monoclonal mouse; Boeringer Mannheim, 1:400) in TBS-TS overnight at 4°C. After rinsing in TBS-TS, the sections were incubated for 3 hours with secondary biotinylated horse anti-mouse IgG antibodies at a concentration of 6 μL/mL (Vector Laboratories, Kemila, Stockholm). After rinsing the sections in TBS, ABC reagent was applied for 1 hour, followed by peroxidase detection for approximately 5 minutes using diaminobenzedine as chromogen (0.25 mg/mL diaminobenzidine, 0.01% H2O2, 0.04% NiCl).
Differential interference contrast photomicrographs were obtained using a Nikon Diaphot microscope equipped with a cooled CCD camera (Hamamatsu, Hamamatsu, Japan) coupled to a computerized digital imaging system (Nikon, Gothenburg, Sweden).
Immunofluorescence
For combined fluorescence BrdU immunolabeling, free-floating sections were treated for DNA denaturation as described above, followed by incubation in TBS-TS for 30 minutes. Thereafter, sections were incubated with rabbit anti-Calbindin D28k antiserum 1:1000 (Swant, Bellizona, Switzerland), GFAp antiserum 1:250 (Advanced Immunochemicals, Long Beach, CA, U.S.A.), and rat anti-BrdU antiserum 1:400 (Harlan, Loughborough, England) for 16 hours at 4°C. Calbindin D28k was detected with Texas red-conjugated donkey anti-rabbit IgG, BrdU was detected with a FITC-conjugated donkey anti-rat IgG, and GFAp was detected with Cy5-conjugated donkey anti-guinea pig IgG (all 1:300; Jacksson ImmunoResearch, West Grove, PA, U.S.A.). After rinsing, all of the sections were mounted and then coverslipped under 30% glycerol in TBS. Fluorescence immunolabeling was examined and processed first using a conventional fluorescence microscope (Nikon Diaphot) coupled to a cooled CCD camera (Hamamatsu). Double labeling for BrdU and either GFAp or CalbindinD28k within single cells was verified using confocal laser scanning microscopy (Bio-Rad MRC1024; BioRad, Richmond, CA, U.S.A.). Z sectioning at approximately 1-μm intervals was used for analysis and verification of double labeling. Pinhole settings were held equal between the laser lines.
Data analysis
All slides were coded before analysis. The code was not broken until the analysis was completed. The total number of BrdU-positive cells in the granule cell layer and the corresponding sample volumes were determined in 7 to 9 coronal sections (240 μm apart), disregarding cells that were in sharp focus in the uppermost focal plane (optical disector principle). The section thickness (40 μm) (microtome setting) was used in the dissector because it was assumed that the net error introduced by using the whole section thickness for the volume was considered to be less than the error introduced by measuring the postprocessing section thickness on each slide and counting in a fixed fraction of it. For sections processed for combined immunohistochemistry, 5 to 6 anatomically matched sections from each animal were analyzed. At least 40 BrdU immunoreactive cells per animal were analyzed concerning phenotype. The number of double-labeled cells were expressed as percentages of the total number of BrdU-labeled cells analyzed.
Statistical analysis
All statistical analyzes were performed with Statwiew 4.01 for Macintosh. For all comparisons of morphologic data, factorial analyzes of variance were performed followed by a Fisher's post hoc test, when appropriate. The level of significance was assumed to be 5%. To assess the percentage of surviving BrdU-positive cells, the individual values of the numbers of BrdU-positive cells at 4 weeks after the last of 7 consecutive injections were divided by the mean number of BrdU-positive cells at 1 day after the injections.
RESULTS
Body weights were significantly less in SHRs than in SD rats. Weight curves for 5- to 10-week-old male and female rats (Fig. 1) closely followed data provided by the breeders for the two strains (Møllegaard).
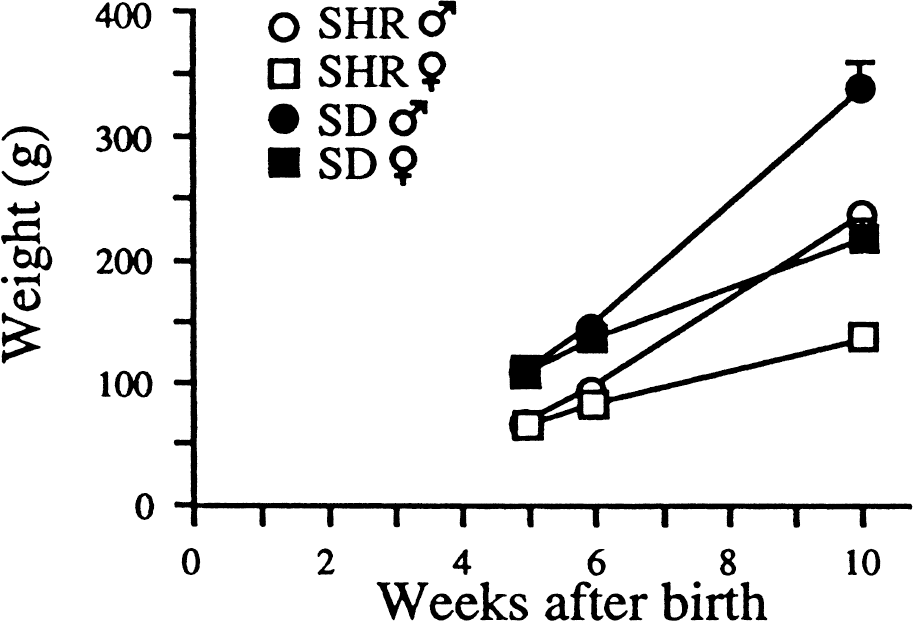
Weight gain of female and male spontaneously hypertensive rats (SHR) and Sprague-Dawley (SD) rats during the experimental period.
Detection of BrdU immunolabeling
Numerous BrdU immunoreactive cells were detected in the dentate gyrus in all groups of animals using either immunohistochemical or immunofluorescent detection (Fig. 2). One day after discontinuation of BrdU injections, proliferating cells could be detected in the subgranular zone and the granule cell layer of the dentate gyrus in the hippocampal region in all groups of the rats (Fig. 2E). The BrdU-labeled cells were most frequently located in the subgranular proliferative zone at the border between the granule cell layer and the hilus with the morphologic appearance of medium-sized elongated to oval-shaped nuclei in both female and male rats (Fig. 2E). The BrdU-immunolabeled cells frequently appeared to be clustered suggesting that the cells were derived from a single ancestral stem-like cell. Some BrdU-immunolabeled cells, often located outside the granule cell layer, had the morphologic appearance of glial precursors or glial cells, that is, small-sized, irregular-shaped nuclei.
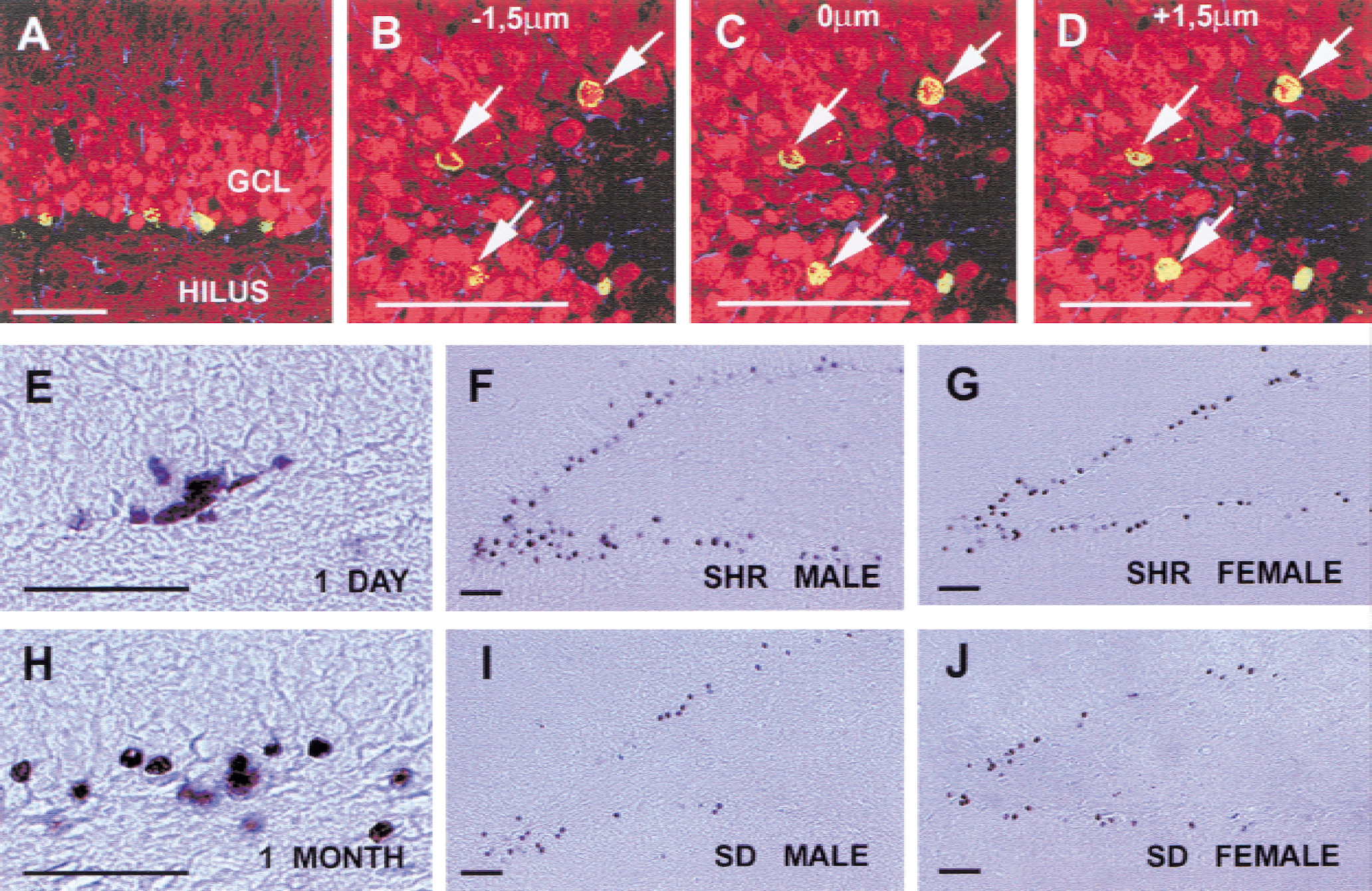
BrdU-labeled cells in the dentate gyrus. Confocal images of sections triple immunolabeled with antibodies against BrdU (green), Calbindin D28k (red), GFAp (blue)
Thirty days after discontinuation of BrdU injections, nuclei were less numerous and appeared to have more round nuclei, frequently with the typical morphology of granule cell chromatin (Fig. 2F to 2J). Cells were most frequently found within the granule cell layer. Changes in morphology and localization of the BrdU-labeled cells suggested that the cells had migrated and differentiated (as described before). However, no obvious morphologic differences could be detected among the BrdU-labeled cells comparing the different groups 30 days after injections. Granule-like cells immunopositive for BrdU and the granule cell marker Calbindin D28k were detected in both male and female SHRs and SD rats 30 days after BrdU injections (Fig. 2A to 2D). In addition, BrdU-positive cells could be detected in the hilar area of the dentate gyrus (CA4).
Sex and strain differences in dentate gyrus proliferation
Male SHRs and SD rats exhibited a significantly greater number of BrdU-immunoreactive cells in the dentate gyrus than their female counterparts one day after the last BrdU injection (F(3,15) = 19.65; P = 0.0001) (Fig. 3). Furthermore, a marked strain difference was observed with significantly more BrdU-labeled cells in SHR compared with SD both in male and female rats (Fig. 3). The cell proliferation in male SD did not differ from that in female SHR.
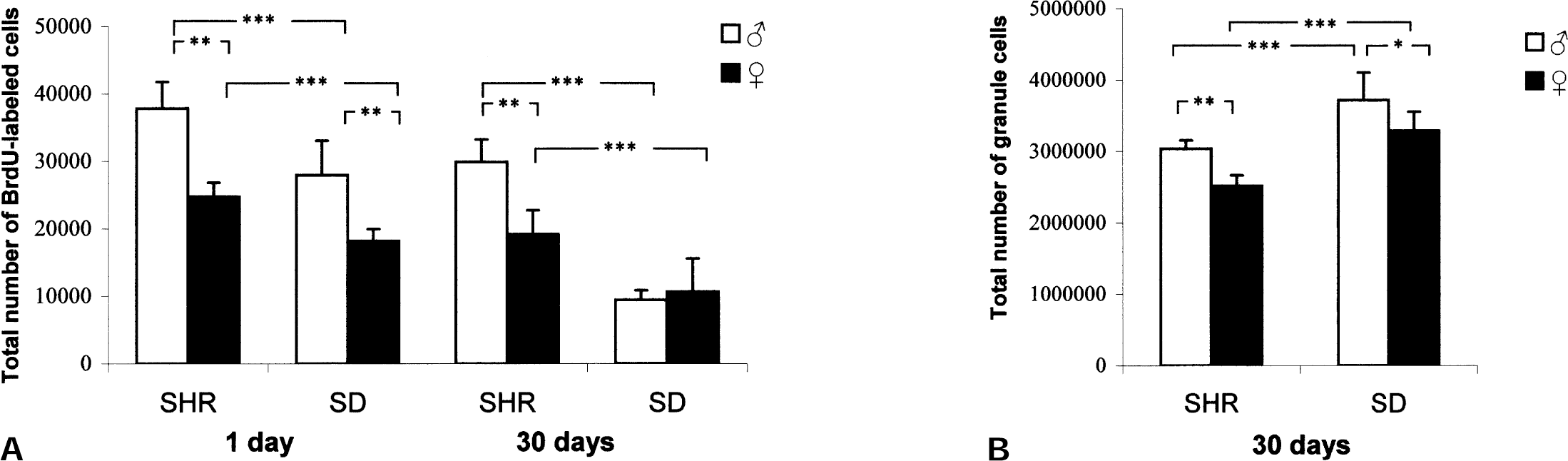
Sex difference in progenitor survival in the dentate gyrus
Four weeks after the last injection of BrdU, survival of newborn cells were estimated (Fig. 3). There were fewer surviving cells after 30 days in all groups. There was a strain difference with a significantly higher number of surviving BrdU-immunolabeled cells 30 days after the last BrdU injection comparing male versus female SHR (F3,16)= 19.65; P = 0.0001). On the other hand there was no detectable difference in the number of surviving cells in male vs. female rats in the SD strain 30 days after the last BrdU injection. Furthermore, there was a marked difference in survival of BrdU immunoreactive cells comparing SHR versus SD both in male and female rats with a more than a 55% increase in survival in SHR compared with SD. The difference was particularly striking between male SD and female SHR which had similar cell proliferation but cell survival rates of 22% and 62%, respectively.
Sex differences in differentiation of BrdU-immunoreactive cells
Co-localization of BrdU immunoreactivity with immunoreactivity for granule cell marker Calbindin and the astroglial marker GFAP was investigated to determine the phenotype of newborn cells 30 days after the last injection of BrdU. Confocal microscopy was used to determine the phenotype of the BrdU-positive cells. Figure 2A to 2D show confocal microscopic images of double-labeled newborn neurons. Neuronal differentiation could be detected in all groups (Fig. 2A to 2D). By four weeks after the last BrdU injection, the majority of these cells were located deep in the gcl and had assumed a morphology indistinguishable from neighboring granule cells. We investigated the phenotype using triple-labeling with antiserum against BrdU, Calbindin D28k and GFAP at 30 days after the last injection of BrdU. We were unable to detect any differences in the fractions of cells positive for both BrdU and Calbindin among the four groups (Table 1). The fraction of cells positive for both BrdU and GFAP was also equal among all groups. The glial cells were found mainly within the subgranular zone and in the hilus.
Phenotype Analysis

Quantification of differentiation of BrdU immunoreactive (progenitor progeny) at 30 days after BrdU injections. Colocalization of BrdU immunoreactivity with the granule cell marker Calbindin D28k and the astrocyte marker GFAp was used to determine the phenotype of progenitor progeny. At least 40 individual BrdU immunoreactive cells per animal were analyzed. The relative rate of the respective phenotype is given as a percentage. No statistically significant gender or strain differences were found in the relative ratio of generated neurons or glial cells. SHR, spontaneously hypertensive rat; SD, Sprague-Dawley, GFAP, glial fibrillary acidic protein.
DISCUSSION
Our data demonstrate a marked gender and strain difference in proliferation and survival of neuronal stem-like cells in SHR and SD rats. The difference in growth and adult weight between these two strains is well known, and does not correspond to any difference in sexual maturation, which occur at 8 to 10 weeks in females and 10 to 12 weeks in male rats in both strains (information provided by the breeder). BrdU was given at the age of 5 weeks to avoid the confounding effect of fluctuation in cell proliferation with the estrous cycle shown to occur in female adult rats (Tanapat et al., 1999).
We have chosen to focus on the dentate gyrus in the hippocampal region because of its well-documented potential for neurogenesis, its well-known cytoarchitectonic structure, its circuitry and its important role in learning and memory.
The cell cycle of granule neurons has been shown to be 16 hours (Nowakowski et al., 1989). Furthermore, BrdU labels DNA only during the S phase of the mitotic cycle. The bioavailability of BrdU for uptake into cells synthesizing DNA has been estimated to last approximately 2 hours (Packard et al., 1973); thus, only a fraction of the total number of dividing cells will be labeled during a 24-hour period after one daily injection of BrdU. The rationale for injecting the animals during seven consecutive days was to be able to measure the mean proliferation. Furthermore, the quantitative impact of continued divisions in the absence of BrdU is decreased by spreading the injections over 7 days.
The lower number of surviving cells 30 days after the end of BrdU injection in all groups corresponds with previous results (Kempermann et al., 1997a, b, 1998a, b; Nilsson et al., 1999). The number of surviving cells may be the net result of proliferation and apoptosis, although apoptosis has so far not been described in BrdU-labeled cells. Recent studies indirectly support this notion by showing that both neurogenesis and apoptosis occur as a result of seizure activity in the dentate gyrus in rat models of limbic epilepsy (Bengzon et al., 1997; Parent et al., 1997).
Sex difference in hippocampal-related structure, function, and neurogenesis
Recent studies have shown a correlation between neurogenesis in the dentate gyrus and spatial learning and memory in female mice (Kempermann et al., 1997b) and male rats (Nilsson et al., 1999). Sex differences in spatial learning favoring males have been documented both in rodents (Beatty, 1984; Williams and Meck, 1991) and humans (Linn and Peterson, 1985; McGee, 1979). In rats, males outperform females on tasks such as the radial arm maze and the Morris water maze test (Roof and Havens, 1992; Williams and Meck, 1991).
Sex difference in progenitor progeny survival
Whereas a comparable fraction of newly formed cells survived in male and female SHR, resulting in a greater net neurogenesis in the males, the survival rate was strikingly reduced in male compared with female SD rats with no difference in net neurogenesis. In a recent study, female SD rats (200 to 280 g; according to the weight, presumably 2- to 3-months-old) were reported to have a greater proliferation and lower survival rate than male rats with no net difference in survival 14 days after the last of 5 BrdU injections.
However, the higher proliferation, studied 2 hours after a single dose BrdU, was shown to occur only during estrous with no difference in proliferation between male and femal rats in proestrus and diestrus (Tanapat et al., 1999). Estradiol previously has been shown to mediate fluctuation in hippocampal synsapse density during the estrous cycle in the female rat (Woolley and McEwen, 1992). Effects of estadiol on proliferation in 2- to 3-month-old female rats may account for the difference in proliferative rate in female versus male rats in the study by Tanapat et al. (1999), compared with the current study in which 5-week-old rats were used.
Increased neurogenesis in SHRs compared with Sprague-Dawley rats
Considering the marked strain differences observed in mice, it is not surprising to find strain differences in rats. Spontaneously hypertensive rat is an inbred strain with many genetic differences from other rat strains. A reasonable assumption would be that the high cell proliferation in the dentate gyrus, at least in part, can be mediated by stimulation of endogenous growth factors, which furthermore might have a stronger effect on male than on female rats. Another evidence for increased cell proliferation in SHR is an exaggerated growth of vascular smooth muscle cells that has been associated with nerve growth factor (NGF) (Spitsbergen et al., 1995; Kapuscinski et al., 1996; Clemow et al., 1998) and some other growth factors (Fukuda, 1997; Fukuda et al., 1997; Nagano et al., 1997). Nerve growth factor, which controls the growth of sympathetic nerves, is increased not only in vascular smooth muscle cells but also in renal tissue in young (2- to 10-weeks old) SHRs (Charchar et al., 1998). It has been proposed that an imbalance between proliferation and apoptosis of smooth muscle cells can account for the abnormality in smooth muscle cell growth in SHR (Diez et al., 1998) and that mutations in or near the NGF gene can, at least in part, explain the high early NGF gene expression in vasculature and kidney in SHR (Sherer et al., 1998, Charchar et al., 1998). A single intraperitoneal injection of an antiserum to NGF into young SHRs (postnatal day 19 to 24) causes a marked reduction in mean arterial blood pressure at the 3 weeks of age (Brock et al., 1996). The blood pressure increases more in male than in female SHRs, which might be another factor contributing to the difference observed in young male and female SHRs in the current study. Whether the content of NGF differs in the brains of SHRs and Sprague-Dawley rats is currently being investigated.
Functional significance of neurogenesis
The functional significance and biologic role of neurogenesis is not well understood. It is, however, tempting to speculate that newborn granule cells in the dentate gyrus forming the three-neuron pathway involved in the establishment of long-term potentiation (Nicoll et al., 1988; Knowles, 1992) might contribute to improved memory function. Enhanced neurogenesis induced by exposure to environmental stimulation has been suggested to contribute to the improved performance in the spatial learning test observed in several recent papers (Kempermann et al., 1997b; Nilsson et al., 1999).
The current results show that male SHRs have a higher net progenitor progeny survival than male SD rats resulting in a higher rate of neurogenesis in male SHRs compared with male SD rats. Functional integration of these neurons seems plausible because newborn neurons in the granule cell layer establish connections to the normal target area in CA3 in the hippocampus (Stanfield and Trice, 1988; Markakis and Gage, 1999). Interestingly, a previous study has demonstrated that young male SHRs outperform young male SD rats in an eight-arm radial maze task (Wyss et al., 1992). This may suggest that species differences in hippocampal neurogenesis may contribute to differences in spatial memory function.
Further studies on gender and strain differences may lead to better understanding of the regulation and functional implications of neurogenesis in the dentate gyrus.
Footnotes
Acknowledgments
The authors acknowledge the expert technical assistance of Ann-Marie Alborn and Barbro Eriksson. The authors are grateful to professors Annica Dahlström, Pam Fredman, and Keiko Funa for generously providing access to their confocal microscope.