
Abstract
General linear time-invariant compartmental systems were examined to determine which systems meet the conditions necessary for application of the spectral analysis technique to the sum of the concentrations in all compartments. Spectral analysis can be used to characterize the reversible and irreversible components of the system and to estimate the minimum number of compartments, but it applies only to systems in which the measured data can be expressed as a positively weighted sum of convolution integrals of the input function with an exponential function that has real-valued nonpositive decay constants. The conditions are met by compartmental systems that are strongly connected, have exchange of material with the environment confined to a single compartment, and do not contain cycles, i.e., there is no possibility for material to pass from one compartment through two or more compartments back to the initial compartment. Certain noncyclic systems with traps, systems with cycles that obey a specified loop condition, and noninterconnected collections of such systems also meet the conditions. Dynamic positron emission tomographic data obtained after injection of a radiotracer, the kinetics of which can be described by any model in the class of models identified here, can be appropriately analyzed with the spectral analysis technique.
Keywords
Linear compartmental systems have been used extensively to describe the kinetics of radiotracers in tissue after injection of the radiotracer into the blood. The compartments in these systems can be physical spaces, or they can represent quantities, specific activities, or concentrations of different chemical species. In the latter case the intercompartmental transfer coefficients reflect the rates of conversion from one species to another. When external radiation detection methods, such as positron emission tomography (PET), are used to determine tissue activities, it is not possible to measure separately compartments that are smaller than the spatial resolution of the instrument, nor is it possible to distinguish among the various chemical species. It is rather the total of the radioactivities in all the compartments in the field of view that is measured. Nevertheless, most of the literature on compartmental systems has been concerned with measurement of the content of individual compartments, and little attention has been directed to the particular problem of characterizing the sum of the contents in all compartments of the system.
Analyses of the kinetics of tracers in which tissue activities have been measured with PET have usually been based on a specific compartmental model structure postulated from known physical or biochemical properties of the tracer. Use of a predefined model structure, however, may lead to significant errors if the model fails to take into account all the relevant biochemical compartments, or if it fails to account for the heterogeneity of tissues included in the field of view of measurement (Schmidt et al., 1991, 1992). As an alternative to the use of a fixed kinetic model, a spectral technique based on a more general linear compartmental system has been introduced and used for analyzing dynamic PET data (Cunningham and Jones, 1993; Cunningham et al., 1993; Turkheimer et al., 1994, 1998). The spectral analysis technique does not require that the number of compartments be fixed a priori, but rather it provides an estimate of the minimum number of compartments necessary to describe the kinetics of the system. The spectral analysis technique applies to heterogeneous as well as homogeneous tissues; this makes it particularly useful for the analysis of tracer kinetics in brain with PET because the limited spatial resolution of the scanner assures that most, if not all, measurements include activities from a heterogeneous mixture of gray and white matter tissues. Furthermore, unlike another widely used technique that can also be applied to heterogeneous tissues, the multiple-time graphical analysis technique (Patlak et al. 1983; Patlak and Blasberg, 1985), spectral analysis does not require steady-state conditions for the tracer. The spectral analysis technique also provides an estimate of the rate constant of trapping of tracer in the tissue as well as the amplitudes and decay constants of the reversible components (Cunningham and Jones, 1993). This information can be used for subsequent specification of a kinetic model, or it can be used to estimate selected parameters of the system that do not depend on the specific model configuration, such as the total volume of distribution of the tracer (Cunningham and Jones, 1993; Turkheimer et al., 1998). Recently, resampling-based procedures have been developed to examine the statistical properties, including variance and bias, of the estimates produced when the spectral analysis technique is applied to single data sets (Turkheimer et al., 1998).
The spectral analysis technique cannot be used with all linear compartmental systems, however; there is a set of conditions that must be fulfilled for it to be applied. It is the purpose of the present study to identify a class of compartmental systems that meet these conditions. Strongly connected systems that contain no cycles (or contain cycles that obey a specified loop condition) and for which exchange of material with the environment is confined to a single compartment are shown to meet the conditions necessary for application of spectral analysis. Certain weakly connected systems that include traps as well as noninterconnected collections of such systems are also shown to meet the conditions. These compartmental structures may prove to be useful for modeling of biological systems in which unmetabolized tracer delivered by the blood exchanges with a pool of unmetabolized tracer in the tissue before undergoing parallel series of metabolic changes or binding to various receptors in the tissue.
SPECTRAL ANALYSIS OF TOTAL TISSUE RADIOACTIVITY
Most kinetic models used with PET tracers are based on the description of the system as a collection of well-mixed compartments with exchange of tracer among the compartments that is assumed to follow first-order kinetics. Tracer delivered to the tissue by the arterial blood exchanges with one or more compartments in the tissue, and exchange of tracer among the various tissue compartments can occur as well. The spectral analysis technique assumes that the total concentration of tracer in the field of view can be written as the sum of convolution integrals
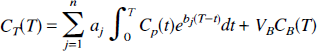
where CT(T) is the total activity in the field of view at the time T of the measurement; n is the number of detectable compartments in the tissue; Cp(t) represents the arterial input function (t minutes after injection of tracer); aj and bj are parameters that depend on exchange of tracer among the compartments; VB is the fraction of the total volume occupied by the blood pool (0 ≤ VB ≤ 1); and CB(T) is the concentration of radioactivity in the blood. In equation 1 Cp(t) may represent arterial blood concentration, arterial plasma concentration, or metabolite-corrected, non-protein-bound plasma concentration of activity depending on which fraction of radioactivity in the blood is exchangeable with the tissue. CB(T), on the other hand, always relates to the total radioactivity in the blood pool. The exponents bj are assumed to be nonpositive real numbers and the coefficients aj to be nonnegative real numbers. The remainder of the paper is concerned with identifying a class of systems in which these conditions, which will be referred to as the spectral analytic conditions, are met, namely, (1) the total tissue activity can be described by an equation of the form of equation 1, (2) the exponents bj are nonpositive real numbers, and (3) the coefficients aj are nonnegative real numbers. We begin with an examination of linear compartmental models.
THE TIME-INVARIANT LINEAR COMPARTMENTAL MODEL
The general n–compartmental system is described by the state equation
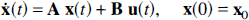
where
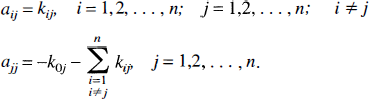
The elements of
Solution of equation 2 yields
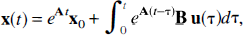
where the term e
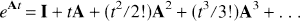
and
The m observed outputs,
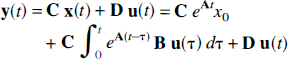
where
CHARACTERISTICS OF PET DATA
In the case of dynamic PET data, there is initially no radioactivity in the system, i.e.,
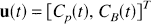
but only the first of these inputs is available for influx into the tissue. The second input does not enter the tissue, but remains in the vascular space and is included in the total activity measurement. If k10, k20, &, kn0 are the influx rate constants into compartments 1, 2, &, n, then
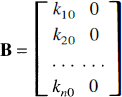
Each measurement consists of one value (m = 1), namely the total activity in the region or pixel, which consists of the sum of the concentrations in each of the tissue compartments plus the radioactivity in the blood pool. Therefore, for this system
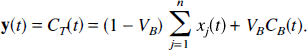
STRONGLY CONNECTED SYSTEMS THAT MEET THE SPECTRAL ANALYTIC CONDITIONS
First, consider strongly connected noncyclic compartmental systems, i.e., systems for which it is possible for material to reach every compartment from every other compartment, but for which it is not possible for material to pass from a given compartment through two or more other compartments back to the starting compartment. Strongly connected noncyclic systems include the catenary, the mammillary, and the catenary-mammillary (CM) systems described by Thron (1982) (Fig. 1). They also include strongly connected noncyclic combinations of any of these systems (Fig. 1). The state transition matrix of a noncyclic strongly connected system is sign-symmetrical, i.e., aijaji ≥ 0, i ≠ j; aij = 0 if and only if aji = 0, and diagonally symmetrizable, i.e., similar to a symmetric matrix via a diagonal similarity transform (Hearon, 1963). It will be shown in the subsequent sections that the key property of diagonal symmetrizability, together with restricting the environmental input and output to the same compartment, is sufficient to assure that the system meets the spectral analytic conditions.
Strongly connected compartmental systems that do not contain cycles: catenary (
Nondefective coefficient matrix
A matrix with distinct eigenvalues is always nondefective (Reid, 1983). Because any coefficient matrix taken at random would be more likely to have distinct eigenvalues than repeated ones, we would expect nondefective coefficient matrices to be the most commonly encountered ones. For this reason
The state transition matrix
Real-valued nonpositive eigenvalues
The eigenvalues of a compartmental system may, in general, be either real or complex numbers, but the real part of every complex eigenvalue is always nonpositive (Hearon, 1963). The similarity of
Nonnegative coefficients
Nonnegativity of coefficients has been shown to occur in the content of a single compartment of a noncyclic, strongly connected system if that single compartment is the only compartment initially loaded (Hearon, 1979). This is equivalent to the assertion that if the system is initially in the zero state and a single compartment receives an input u(t) from the environment, then that compartment will exhibit nonnegative coefficients of its component terms. The theorem is formally restated below; the symbol
Monotonicity Theorem (Equivalent Form) (
Hearon, 1979
). Let the state transition coefficient matrix
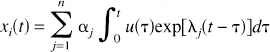
where α j ≥ 0. Note that the nonnegative coefficients apply only to compartment i.
To examine the coefficients of the sum of the concentrations in all compartments, we now look at those strongly connected compartmental systems in which exchange of material with the environment is confined to a single compartment. If compartment i is the only compartment from which material can pass from and to the environment, then the change in concentration in the sum of compartments is the difference between the influx into compartment i and the efflux from compartment i:
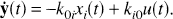
If the system is closed, i.e. k0i = 0, then integration of equation 11 produces
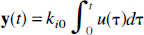
which clearly has only positive coefficients. On the other hand, if the system is open, i.e., k0i > 0, then λ j is real and negative, as shown in the discussion of real-valued nonpositive eigenvalues, and we can apply the Monotonicity Theorem stated above to obtain
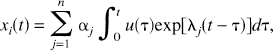
where α j ≥ 0. Substituting equation 13 into equation 11 yields
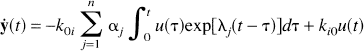
which on integrating becomes
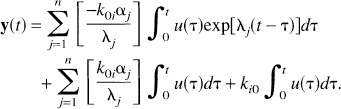
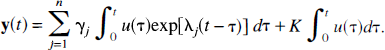
It follows immediately from λ j < 0, –k0i < 0, and α j ≥ 0 that γ j ≥ 0. The coefficient K is the steady-state total concentration in the system after a unit impulse input; because the strongly connected open system contains no traps, K = 0.
SYSTEMS WITH TRAPS
What happens when the requirement for strong connections in the open system is relaxed? Consider the matrix
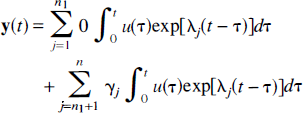
where γ j ≥: 0 and λ j > 0.
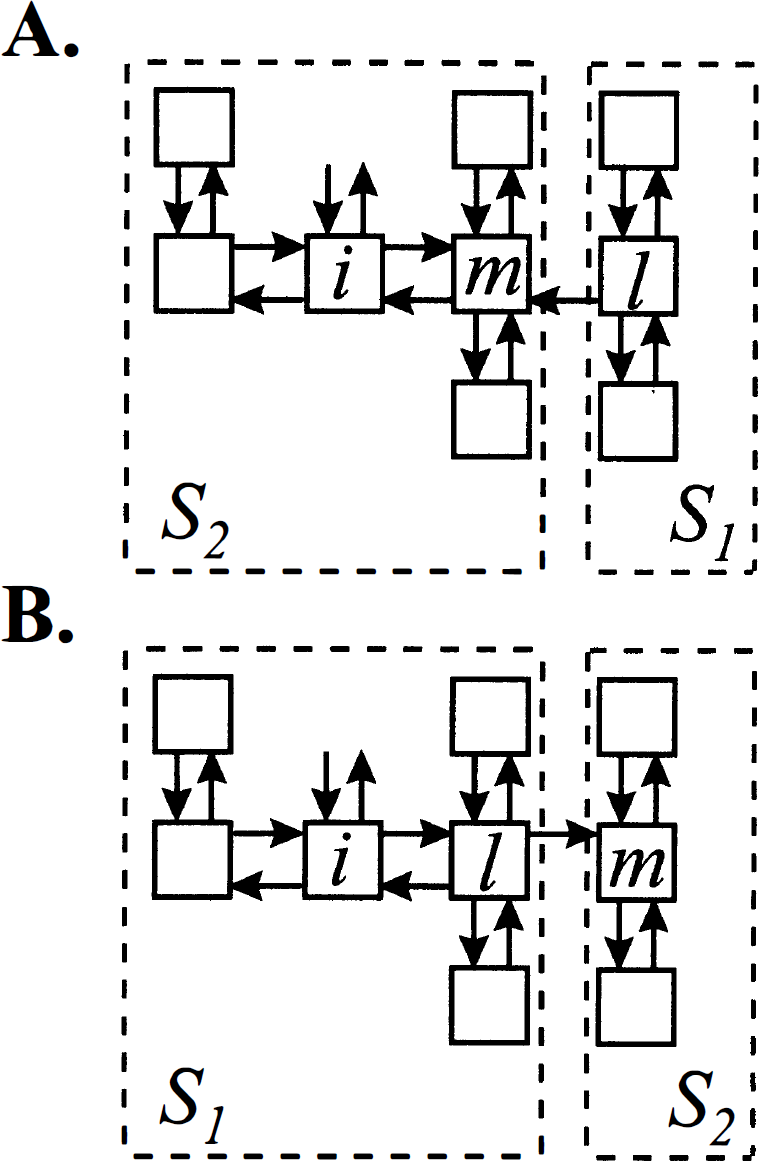
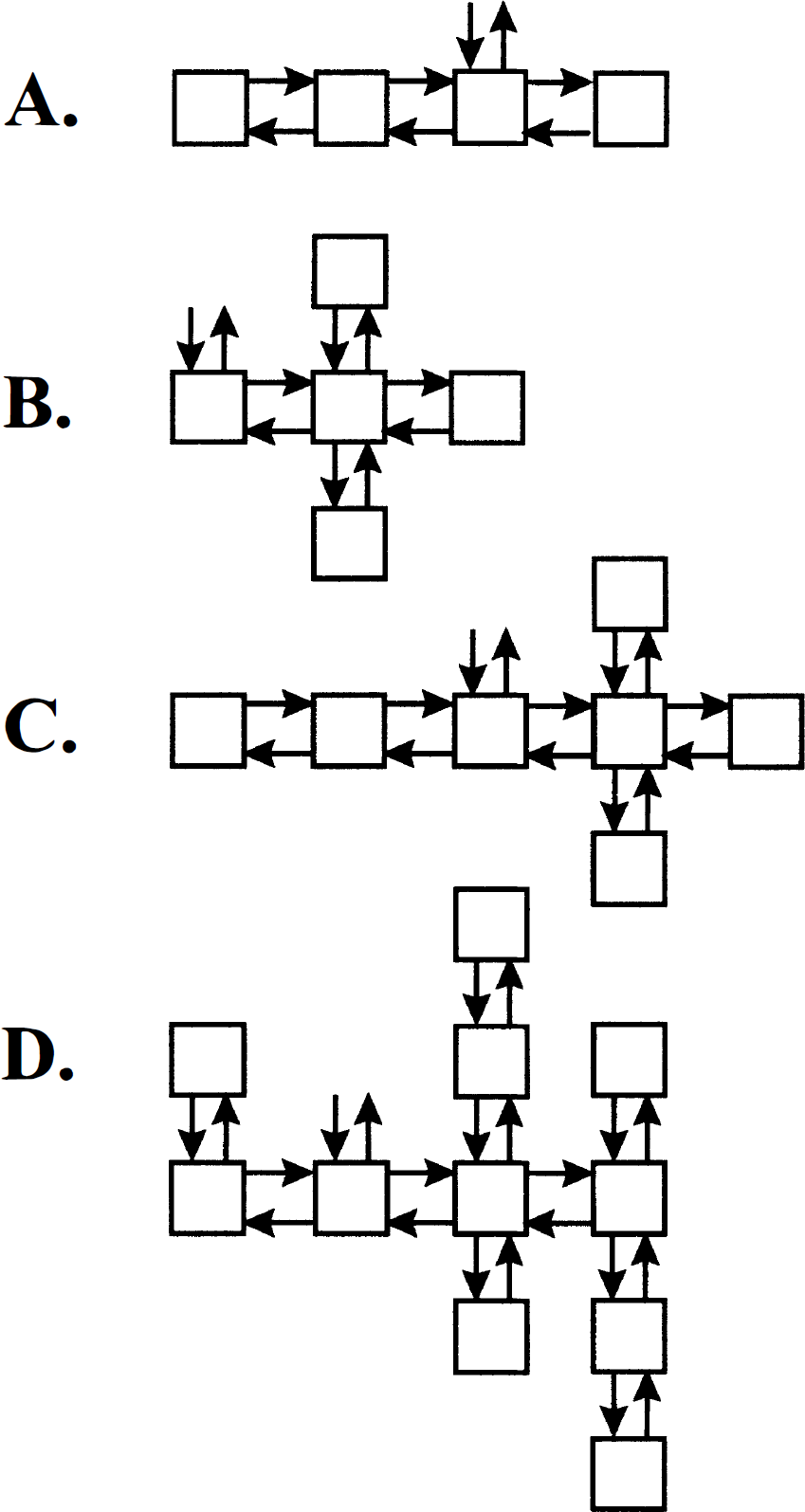
Compartmental systems formed by removing a single link from a strongly connected noncyclic system. Removal of the link from compartment m to compartment l splits the system into two subsystems. Input from the environment and excretion to the environment is confined to compartment i. (
Now consider the case when compartment i is in subsystem S1 (Fig. 2B). S1 is noncyclic, strongly connected, and has no traps. Hence, by the Monotonicity Theorem stated in the previous section, the concentration in the ith compartment is
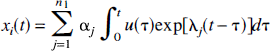
where α j ≥ 0 and λ j < 0. The rate of change in the total concentration in the combined system S1 + S2 is, as it was for the strongly connected system, given by equation 11. Equations 14 and 15 also hold (with n replaced by n1) so that, for this system
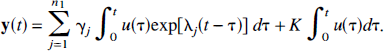
where
It is clear that removal of additional links in the subsystem that is not reachable from the input compartment, such as subsystem S1 in Fig. 2A, or from a subsystem that is already a trap, such as S2 in Fig. 2B, does not alter the total concentration of the system. Removal of an additional link in the subsystem connected to the compartment that exchanges with the environment leads to a partitioning of that subsystem, and creates a new subsystem that either is not input-reachable or is another trap. In either case, the characteristics of the total measured output, namely nonnegativity of coefficients and nonpositivity of eigenvalues, are preserved. Thus any compartmental system that can be constructed by removing links from a noncyclic, strongly connected system that has exchange with the environment confined to a single compartment satisfies the spectral analytic conditions. Examples are illustrated in Fig. 3.
Compartmental system that is not strongly connected (
SYSTEMS THAT CONTAIN CYCLES
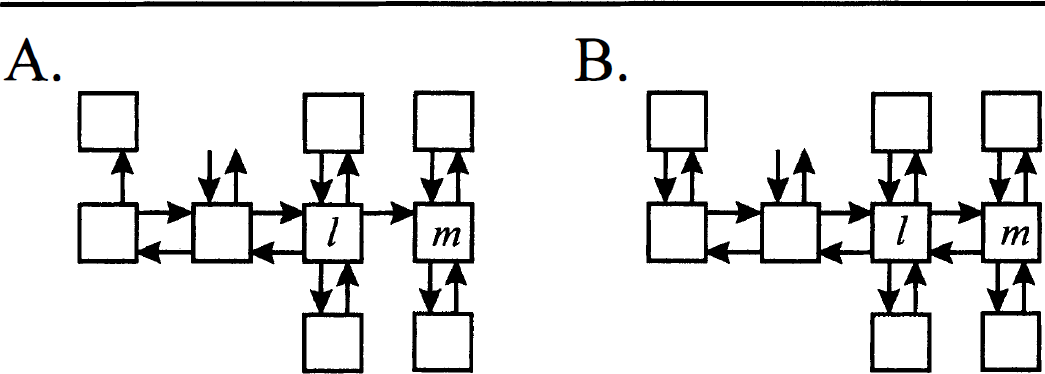
Diagonal symmetrizability, the key property from which the Monotonicity Theorem follows, does not require that the system be strictly without cycles, but rather that all cycles present satisfy a specific loop condition (Hearon, 1963, 1979). Recall that a cycle exists if it is possible for material to pass from one compartment through two or more other compartments back to the initial compartment, i.e., there is a set of three or more indices α, β, γ, & ρ in which none of the transfer coefficients kβα, kγβ, & kαρ is zero. A system will be said to satisfy the loop condition if, for every cycle, the product of rate constants is the same regardless of which direction material is transferred through the loop, i.e.,
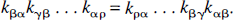
Noncyclic systems trivially satisfy equation 20. If the state transition matrix
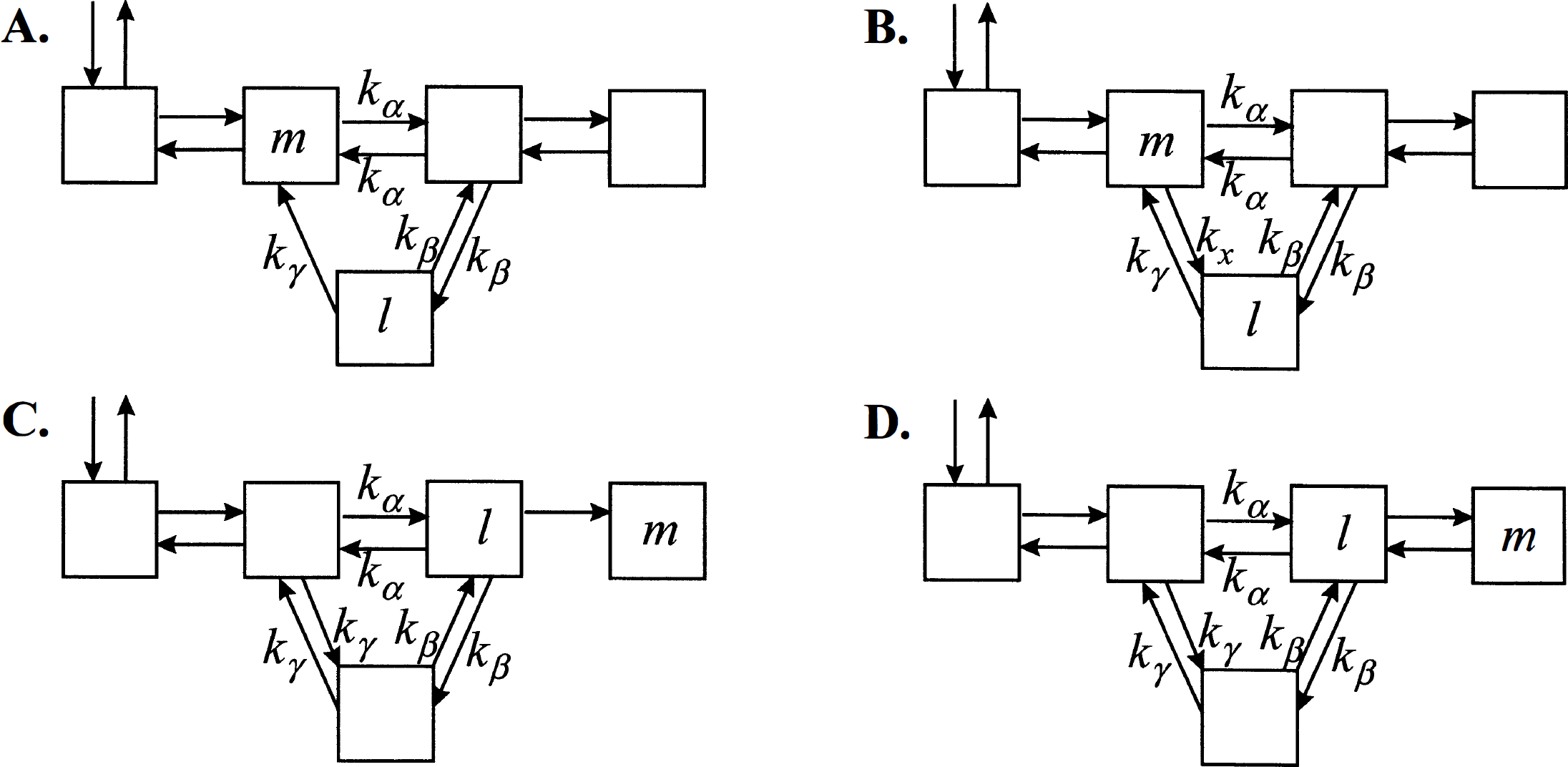
Compartmental systems with cycles. (
RECAPITULATION
After the simple addition of the blood pool contribution to the total measured activity, the identification of a class of kinetic models to which spectral analysis can be applied for the analysis of PET data is summarized in the following theorem.
THEOREM. Let
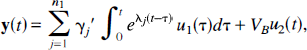
where n1 ≤ n, λ j are real, λ j ≤ 0, and γ j ' = (1 –VB)γ j = (1-VB) (–k0iα j /λ j ) ≥ 0.
DISCUSSION
The above analysis does not address whether the full complement of n eigenvalues λ j will be visible in the output. As has been previously shown (Hearon, 1963; Fagarasan and DiStefano, 1986), the symmetrizability of the system provides that if the system has repeated eigenvalues, then the system has hidden eigenvalues regardless of what compartmental contents are measured. This is evidenced by the absence of terms of the form tk eλjt for k > 0 in equation 1. When the system contains multicompartmental subsystems that function as a trap, there are modes (eigenvalues) of the system that are not visible in the measured sum of all compartments. In the most extreme example, when a system has no efflux to the environment and only the sum of all compartments is measured, then only the zero eigenvalue is visible in the output; all other modes are hidden. The presence of compartments that cannot be reached from the input compartment also leads to hidden modes in the system. For a further discussion on hidden modes, the reader is referred to Fagarasan and DiStefano (1986).
It should be emphasized that the conditions on the compartmental structure discussed here are sufficient for applying the spectral analysis technique, but they are not necessary conditions. A condition on the coefficient matrix
The compartmental structures shown in the present study to satisfy the spectral analytic conditions include the catenary, mammillary, and various combinations of these systems when input from the environment and efflux to the environment are confined to the same single compartment in the system. Any system in which tracer delivered by the blood exchanges with a single compartment in the tissue and then undergoes one or more series of metabolic or binding transformations satisfies these conditions. The one-compartment kinetic model used for measurement of cerebral blood flow with a freely diffusible tracer in a homogeneous tissue, originally designed for use with [14C]iodoantipyrine and quantitative autoradiography in animals (Sakurada et al., 1978) and now used with H215O and PET in humans (Huang et al., 1983; Herscovitch et al., 1983), is a simple model that satisfies the spectral analytic conditions (Fig. 5A). The two-compartmental model for measurement of glucose metabolism in homogeneous tissues with either [14C]deoxyglucose (Sokoloff et al., 1977) or [18F]fluorodeoxyglucose (Reivich et al., 1979) includes the assumption that influx of tracer from the plasma and efflux of tracer to the plasma occurs in only one compartment (Fig. 5B). Thus it clearly satisfies the conditions for application of spectral analysis. The three-compartment model used in some receptor-ligand binding studies, such as those designed to assess muscarinic cholinergic receptor binding by use of [11C]scopolamine (Frey et al., 1985) or to assess benzodiazepine receptor density by use of [11C]flumazenil (Koeppe et al., 1991), also assumes a single tissue pool of free ligand that exchanges with the free ligand in plasma (Fig. 5C). The system does not contain any cycles, and therefore this model, too, satisfies the spectral analytic conditions.
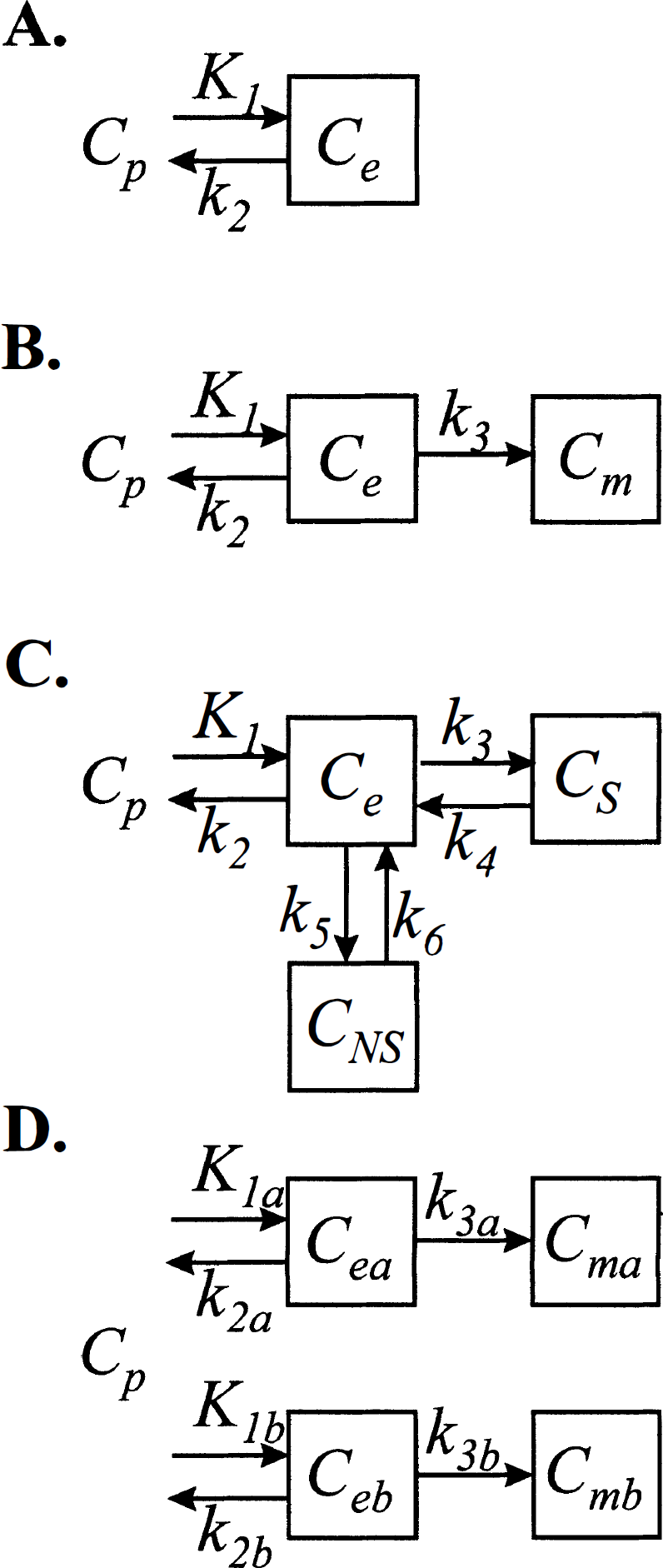
Compartmental models that satisfy the conditions for application of spectral analysis to total tissue activity. (
The preceding models were designed for application in homogeneous tissues. A collection of replicates of the compartmental systems designed for homogeneous tissues, however, can be used to model the activity in a heterogeneous mixture of tissues (Fig. 5D), because activity in a heterogeneous mixture of tissues is the weighted sum of the activity in the constituent tissues constituting the mixture (Schmidt et al., 1991). Because of the additivity of the components in equation 1, a collection of systems that are not interconnected and in which each independent system in the collection satisfies the spectral analytic conditions also satisfies the spectral analytic conditions. This property renders the spectral analysis technique particularly useful for the analysis of tracer kinetics in PET studies in which several kinetically dissimilar tissues may be in the field of view of the PET camera because of its limited spatial resolution.
Spectral analysis is not an appropriate tool for analyzing models that contain feedback loops, that have efflux from more than one compartment, or that consist of connected collections of subsystems, even if the subsystems of the collection meet the conditions for application of spectral analysis (Fig. 6). For example, the three-compartment model for deoxyglucose that accounts for the intracellular compartmentation of glucose phosphatase contains a feedback loop (Schmidt et al., 1989) (Fig. 6A). For some combinations of values of the rate constants, the eigenvalues bj of the system are complex-valued; hence oscillations in the system are possible. Unless the feedback rate constant, k5, is zero, this model is not a candidate for application of spectral analysis. The model of Huang et al. (1991) that describes the kinetics of L-3,4-dihydroxy-6-[18F]fluoro-phenylalanine ([18F] FDOPA) and L-3,4-dihydroxy-6-[18F]fluoro-3-O-methylphenylalanine ([18F]3-OMFD) in the striatum is given in Fig. 6B. It consists of two unconnected catenary subsystems; hence the eigenvalues of the system are real-valued. Spectral analysis cannot be applied to the total tissue activity in this system as configured, however, because the positivity of coefficients is not assured unless the rate constant k4, loss from the fluorodopa metabolite compartment, is zero (Fig. 6B). If k4 were zero, for example, if the experimental period were sufficiently short that loss of metabolites were negligible, spectral analysis could be applied to the total tissue activity with a simple modification to include the two input functions, i.e.,
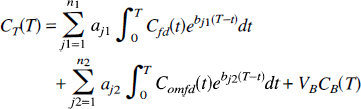
where Cfd(t) and Comfd(t) are the arterial plasma concentrations of [18F]FDOPA and [18F]3-OMFD, respectively. Two subsystems are also used in the model of Gjedde et al. (1991) to describe [18F]FDOPA and [18F]3-OMFD kinetics (Fig. 6C). Gjedde and coworkers have assumed that there is no loss of fluorodopa metabolites during the experimental period, so both subsystems in this model meet the conditions necessary for application of spectral analysis. Because this model includes the possibility of conversion of [18F]FDOPA to [18F]3-OMFD in the tissue, however, the subsystems are connected and the complete system may not satisfy the conditions necessary for application of spectral analysis to total tissue activity.
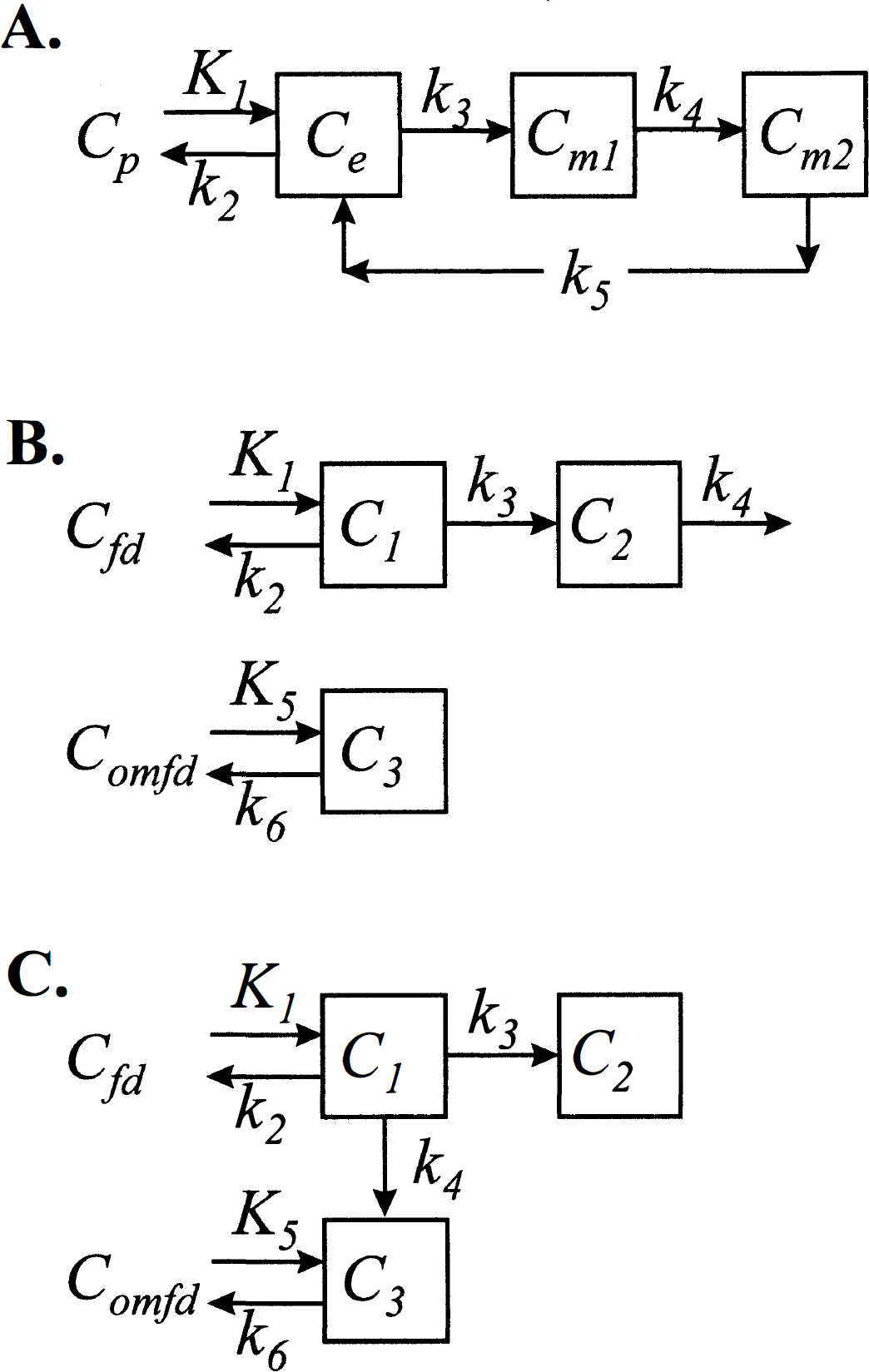
Compartmental models that do not meet the conditions for application of spectral analysis. (
Spectral analysis is potentially a useful tool for the analysis of dynamic PET data that does not require that the kinetic model of the system be completely specified a priori. Instead, it can be used for estimating the minimum number of compartments needed to specify the system and to characterize the components of the system. It provides information that can be used for subsequent specification of a kinetic model, or it can be used to estimate selected parameters of the system that do not depend on the specific model configuration. It applies to heterogeneous as well as homogeneous tissues. Although it is based on a general linear compartmental system, it does not apply to all such systems. The present study has taken a first step in identifying a class of compartmental systems for which the analysis is appropriate.