
Abstract
The corpus callosum is the largest connection between the functionally asymmetric cerebral hemispheres. The objective of this study was to measure functional activity of callosal fiber tracts during speech processing. We analyzed the regional glucose metabolism of the corpus callosum and of speech-relevant cortical areas in 10 normal individuals at rest and during word repetition. We used three-dimensionally registered magnetic resonance imaging to visualize the individual brain morphology and high-resolution positron emission tomography for metabolic measurements. The task-induced metabolic changes of the callosal midbody and isthmus had a significant negative correlation with key regions of language processing in the left inferior frontal cortex (Brodmann's area 44) and in the right superior temporal cortex (Brodmann's area 22) (e.g., correlation of metabolic changes in the surface aspects of the right Brodmann's area 22 and the callosal midbody/isthmus: r = −0.91, P < 0.001). The study indicates that language processing in asymmetrically organized cortical areas inhibits the reciprocal transcallosal information exchange in favor of the lateralized mental operation. Our data agree with anatomic, electrophysiologic, and pharmacologic experiments that point to the important role of collateral inhibition for the transcallosal information exchange.
Keywords
The two hemispheres … speak as a unit in that the right hemisphere during speech is not idling or diverted but is actively focused to aid and sustain the cerebral processing involved in speech, to add tone and expression and to inhibit unrelated activity (Sperry, 1986).
The language dominance of the left cerebral hemisphere requires the reciprocal adjustment of neuronal activity in both cerebral cortices. The corpus callosum is the largest interhemispheric fiber tract that connects the left and right cortical networks. Studies of split-brain patients demonstrate that lateralized cognitive operations, such as language processing, involve interhemispheric information transfer through the corpus callosum (Gazzaniga, 1970). Morphometric studies of human postmortem brains, however, indicate that the interhemispheric information exchange is not equally organized in all humans, but that it varies considerably with the degree of the individual brain functional asymmetry. Taking handedness as a measure of functional lateralization, these studies have found that the midsagittal size of the human corpus callosum and, consequently, the number of transcallosal fibers are larger in left-handed and ambidextrous people than in those with consistent right-hand preference (Witelson, 1989; Cowell et al., 1993).
Neuroimaging with positron emission tomography (PET) can provide information about the functional interactions within entire neural networks. Two major strategies exist by which such PET data are currently analyzed. One strategy compares the pattern of activity between two or more tasks, looking for those areas that show significant changes. The second determines the functional relationships between regional activities in an attempt to find the neural network that mediates the task (McIntosh et al., 1994; Horwitz et al., 1995). Using this second, correlational approach, previous studies of regional brain metabolism revealed that a widespread network of significant connections orchestrates the speech-relevant areas of both cerebral hemispheres according to the actual needs of speech processing (Karbe et al., 1998). Within this large bilateral network of functionally intercorrelated cortex areas, specially close interhemispheric correlations connect relatively small portions of the left and right auditory association cortex that are situated in that part of the left Brodmann's area (BA) 22 that is buried in the superior temporal sulcus and in the surface aspects of the right BA 22 (Karbe et al., 1995). The present study explores whether the energy consumption of the corpus callosum reflects the dynamics of the presumed interhemispheric transmission of language information.
PATIENTS AND METHODS
We studied 10 normal individuals (9 men, 1 woman, mean age 39 years, SD 14 years) with three-dimensional-registered magnetic resonance imaging (MRI) and with high-resolution PET using the 18F-2-fluoro-2-deoxy-D-glucose method. A laterality questionnaire and standardized tests of skilled motor performance classified all subjects as consistent right-handers (Oldfield, 1971). All subjects gave their informed consent. Magnetic resonance imaging was performed in all individuals with a superconducting 1-T instrument (Magnetom, Siemens, Erlangen, Germany). The MRI scanner produced 64 adjacent transaxial T1-weighted tomograms with a slice thickness of 2.5 mm and a pixel size of 0.98 × 0.98 mm. Positron emission tomography was performed with a high-resolution scanner (ECAT EXACT HR, Siemens-CTI, Knoxville, TN, U.S.A.), which provided 47 contiguous transaxial slices through the brain with a center-to-center distance of 3.125 mm and a spatial resolution (full width-half maximum of point-spread function) of 3.6 mm transaxially and 4 mm axially (Wienhard et al., 1994). After bolus injection of 185 MBq 18F-2-fluoro-2-deoxy-D-glucose, six frames of 10 minutes each were recorded. Images were reconstructed on a 128 × 128 matrix with 2.17-mm pixel size by applying a Hanning filter with a cutoff frequency of 0.4 cycles per pixel. This procedure included corrections for random coincidences, scatter, and attenuation; with regard to attenuation, a threshold algorithm was used to define the contour of the head. Images of the regional cerebral metabolic rate of glucose were calculated based on frames 3 to 6 and multiple arterialized blood samples using the autoradiographic model with adjustment of rate constants K1 and k3 to measured activity, as previously described (Wienhard et al., 1985).
Two PET studies were performed in each individual within 1 week. In the first PET study, the individuals were examined in a resting condition, with eyes closed and ears unplugged, in a room with low ambient noise (Heiss et al., 1984). The second PET study was performed under the same condition but during verbal stimulation. The subject had to repeat German nouns that one of the investigators randomly presented aloud. The total amount of presented nouns, which was adapted to the individual repetition pace, was 1,224 (SD, 242) words in this study. The period of stimulus presentation and the number of presented and repeated words clearly exceeded the range in which the number of presented stimuli correlates with the degree of regional cortical activation (Price et al., 1992). Thus, the number of presented words did not correlate significantly with the regional metabolic activation. The 30-minute period of stimulation started immediately before 18F-2-fluoro-2-deoxy-D-glucose injection. The MRI and PET images were coregistered in parallel to the intercommissural line as defined on the three-dimensional MRI display by using the Multi-Purpose Matching Program. The uncertainties from the coregistration were found to be less than 1.5 mm in any direction (Pietrzyk et al., 1994). The original three-dimensional-registered MRI and PET images were interpolated to 1-mm3 voxels. Sagittal, coronal, and transaxial 1-mm slices were created from the 1-mm3 voxels. A software program written in-house allowed a simultaneous, three-dimensional display of MRI and PET images. Regions of interest were placed subsequently on several adjacent MRI slices in one plane. The software program automatically superimposed the regions on the corresponding PET images and simultaneously displayed them on the two other planes so that the three-dimensional extent of a region could immediately be seen (Herholz et al., 1996). Thus metabolic changes were measured in regions based on the individual brain morphology. We analyzed cortical activity in the main regions of language processing, i.e., in the left BA 44, which is the core of Broca's area, in the left BA 22, which is the core of Wernicke's area, in their right-hemispheric mates, and in the entire corpus callosum. The regions of interest were outlined as follows:
The sulcal area of BA 22, i.e., that part of BA 22 that is buried in the superior temporal sulcus. The anterior and posterior borders of BA 22 (sulcus) coincided with the anterior border of Heschl's gyrus and with the posterior border of the planum temporale, respectively. The medial border was defined by the medial fold of the superior temporal sulcus; the lateral border coincided with the lateral border of the planum temporale (Karbe et al., 1995). The part of BA 22 that is located on the surface of the superior temporal gyrus. Brodmann's area 22 (surface) medially started at the lateral border of the Sylvian fissure and stretched to the lateral surface of the superior temporal gyrus (Karbe et al., 1995). Brodmann's area 44, which was defined as the convolution posterior to the ascending ramus of the Sylvian fissure (Foundas et al., 1995). The corpus callosum. The contour of the corpus callosum was delineated on seven sagittal 1-mm slices starting midsagittally on MRI scans. The outer contour of 1-mm3 voxels was excluded from metabolic measurements to reduce partial volume effects. We divided the corpus callosum into three parts on the midsagittal plane: the anterior third (rostrum, genu, rostral body), the middle half (midbody and isthmus), and the posterior sixth (splenium).
The metabolic changes of the anatomically defined regions (Cregional) were calculated from:
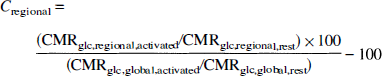
where CMRglc,regional,activated is the CMRglc during activation, CMRglc,regional,rest is the CMRglc at rest, and CMRglc,global,activated and CMRglc,global,rest are the CMRglc of the entire brain during activation and at rest, respectively. The global CMRglc value of the entire brain included all brain structures except for the cerebrospinal fluid space, i.e., ventricles and cisterns were excluded from the calculation. SPSS for Windows, Version 6.0 was used for all statistical calculations.
RESULTS
Table 1 lists the mean metabolic changes of the callosal and cortical regions of interest. Table 2 gives the Pearson correlation coefficients between the three callosal and the six cortical regions. The 18 correlations clearly show a tendency to be different from zero. The Bonferroni step-down adjustment of Holm (Holm, 1979) was used to provide overall protection against reporting significant results that are false. Using this adjustment, two correlations are statistically significant, i.e., the correlations of the callosal midbody/isthmus with the left BA 44 and right BA 22 (surface). Interestingly, both correlations are negative. That means that the callosal activity decreases as the cortical activity increases. Figure 1 shows both significant inverse corticocallosal correlations in a schematic lateral view; Fig. 2 shows the relationship between the right BA 22 (surface part) and the callosal midbody/isthmus in a correlation plot.
Mean metabolic changes with standard deviation during single word repetition
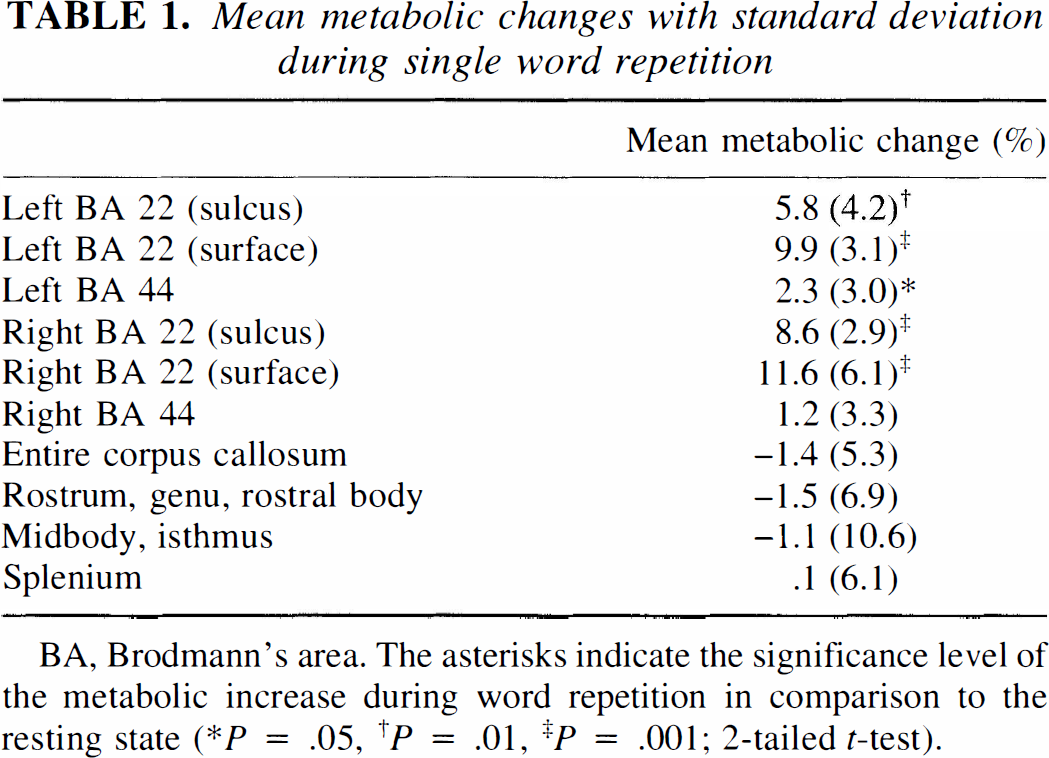
BA, Brodmann's area. The asterisks indicate the significance level of the metabolic increase during word repetition in comparison to the resting state (∗P = .05, †P = .01, ‡P = .001; 2-tailed t-test).
Pearson correlation coefficients (with P value) of the cortical and callosal regions of interest
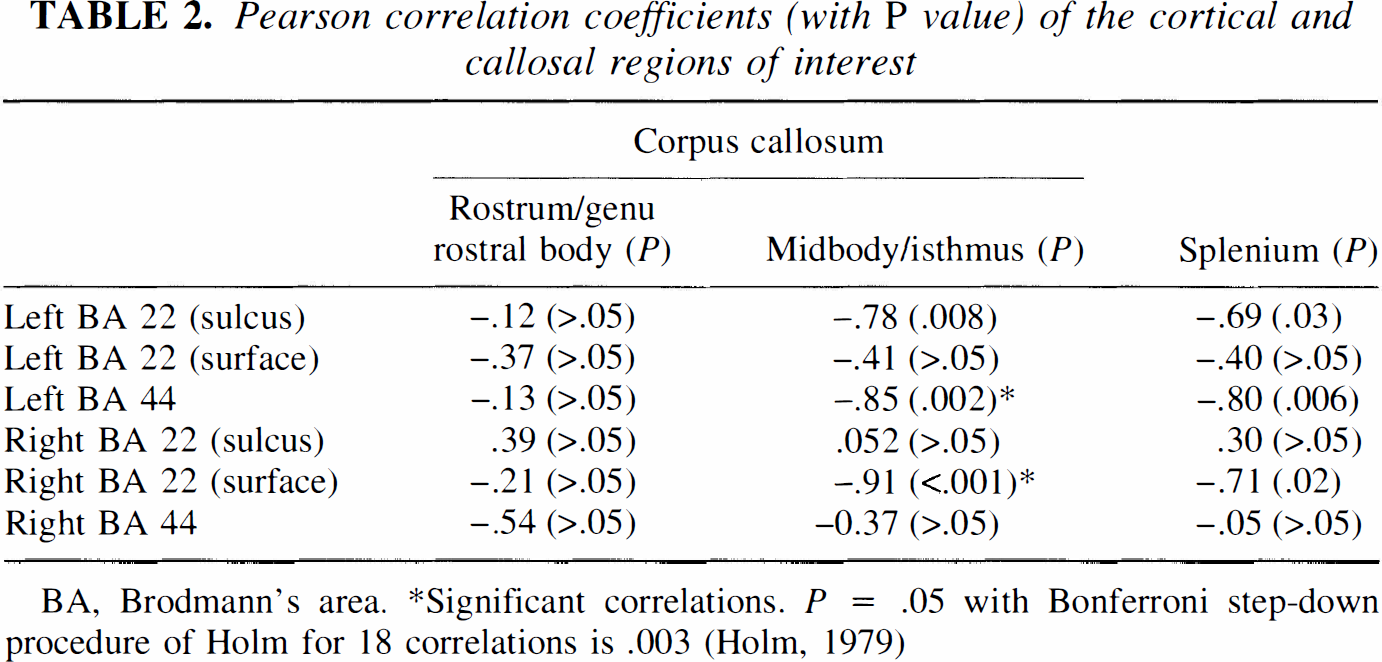
BA, Brodmann's area.
Significant correlations. P = .05 with Bonferroni step-down procedure of Holm for 18 correlations is .003 (Holm, 1979)
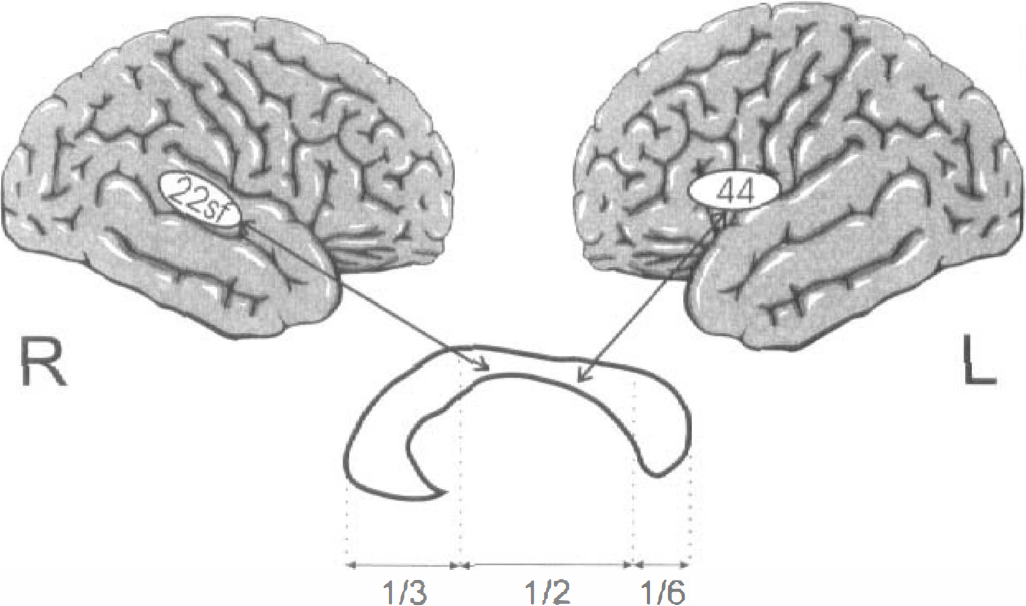
Right Brodmann's area (BA) 22 (surface part) (22sf) and left BA 44 on schematic lateral views of the brain hemispheres. Below is a midsagittal cut through the corpus callosum with the subunits rostrum/genu/rostral body, midbody/isthmus, splenium. The speech-induced activity of both cortical areas has a significant inverse correlation with the activity of the callosal midbody/isthmus.
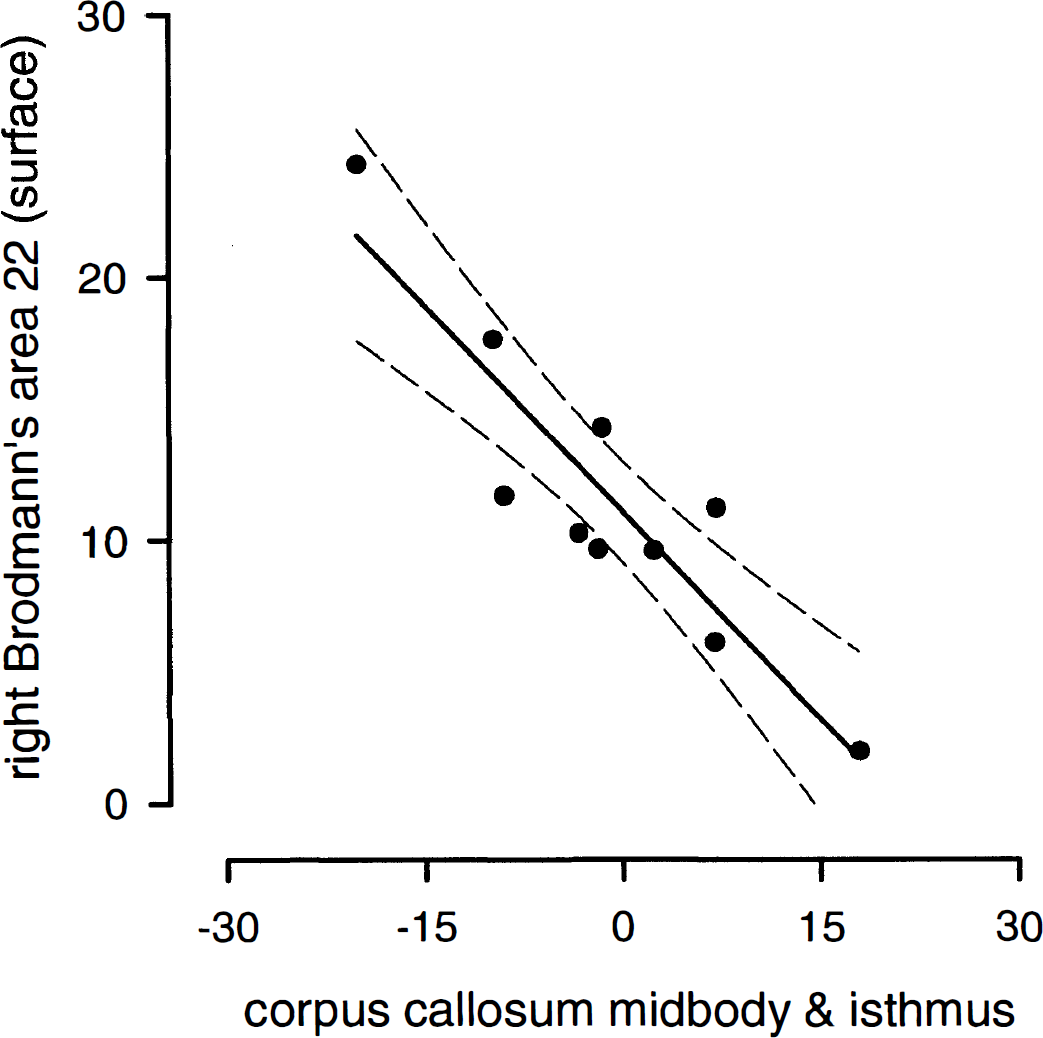
Correlation plot of metabolic changes (%) of the callosal midbody/isthmus and the right Brodmann's area 22 (surface) during word repetition with 95% confidence interval (dotted lines) (r = −0.91, P < 0.001).
DISCUSSION
White matter metabolism has rarely been analyzed with PET in humans, although animal studies indicate that white matter glucose consumption depends on the level of brain functional activity. In their basic description of the deoxyglucose method, Sokoloff et al. (1977) established that the corpus callosum metabolism is 29% higher in conscious than in anesthetized rats. Furthermore, animal studies of neuropeptide-induced hypermetabolism clearly demonstrate that neuroexcitatory effects also involve connecting white matter tracts, especially the corpus callosum (Gross et al., 1992). Using radiolabeled deoxyglucose and PET, our study demonstrates for the first time in humans a significant inverse relationship between cortical gray matter activation and white matter metabolic changes during a mental operation. We identified the callosal midbody and isthmus as a white matter region that mediates functional brain asymmetry between key regions of auditory and language processing. We have to emphasize, however, that the resolution even of the new generation of PET scanners did not allow us to identify single fiber tracts within the callosal midbody. The metabolic changes that we found in this study therefore point to a modulation of intercerebral activity that predominantly, but not exclusively, involves key regions of functional brain asymmetry.
Studies of dichotic listening indicate substantial transfer of lateralized information between the human auditory cortices, on the basic level of word perception (Kimura, 1967). Neuroanatomic studies of animals (Cipolloni and Pandya, 1985) and neuropsychological studies of patients with partial versus complete callosotomy (Musiek and Reeves, 1986) consistently indicate that this transfer between the auditory cortices uses specific fiber tracts of the callosal midbody and isthmus, i.e., that part of the corpus callosum that we identified as speech-relevant in this study. More interesting than the localization of the callosal metabolic changes, however, are the organization and direction of callosal information transfer that our data imply. If the transcallosal auditory communication was a direct projection and simply excitatory, the metabolic increase of callosal fibers would parallel the activation of cortical neurons. We found, however, a negative correlation between callosal and cortical energy consumption during word repetition, i.e., the cortical metabolism of functionally asymmetric areas increased as the callosal metabolism decreased. It has to be emphasized in this context that our sample of individuals only included consistent right-handers, predominantly men, i.e., individuals with presumably left-dominant, asymmetrically organized brains (Shaywitz et al., 1995; Geschwind and Galaburda, 1984). In these brains, we found significant negative corticocallosal correlations. Because the deoxyglucose method directly reflects the local energy utilization accompanying membrane depolarization (Mata et al., 1980), our findings suggest that the activation of specially asymmetric cortex areas reduces the overall number of activated callosal fibers. Finding a modification of activity in such a large part of the corpus callosum, furthermore, indicates that key regions of brain functional asymmetry may be able basically to adjust the interhemispheric information exchange beyond a simple one-to-one talk with their contralateral mate. That means that lateralized mental operations can only be executed when the transcallosal activity is reduced to the actual operational needs, or in other words, that the orchestra has to decrease in volume when the soloist is playing. Figure 3 schematically illustrates this concept of transcallosal mediation of functional brain asymmetry.
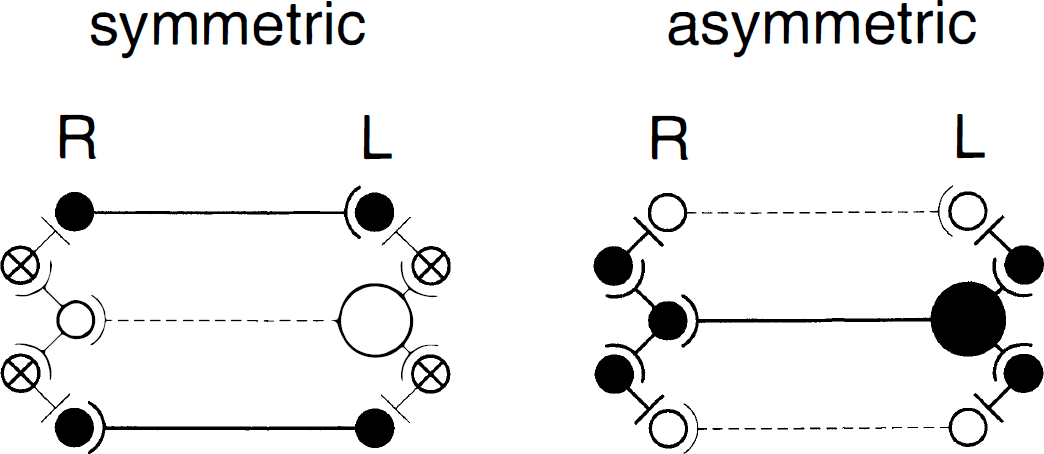
Schematic diagrams of symmetric and asymmetric interhemispheric information processing. The left diagram shows the symmetric transmission of nonlateralized information. Neurons of both hemispheres (filled circles) reciprocally excite neurons in their contralateral projection field. Inhibitory neurons (crossed circles) and specific neurons with functional dominance (large circle) are not involved. The right diagram shows the excitation of a functionally dominant neuron of the left hemisphere (large circle) during specific information processing. The excitation of the dominant neuron is transmitted to local inhibitory interneurons, ipsilaterally and contralaterally. The information transfer to the contralateral projection field uses a disynaptic feed-forward circuit of excitatory/inhibitory transcallosal transmission. Thus inhibitory neurons bilaterally suppress the background activity of symmetric interhemispheric information exchange. Because the excitation of inhibitory neurons is also energy consuming, the activation of the dominant neuron (large filled circle) induces a significant bilateral metabolic increase that is closely correlated. Callosal metabolism decreases at the same time because the overall number of excited transcallosal fibers decreases in favor of the transmission of excitatory/inhibitory information through the specific transcallosal axon.
Interestingly, callosal metabolism increased to some extent in 4 of 10 individuals during word repetition in comparison with the resting state (Fig. 2). Moreover, this finding smoothly fits our model of collateral inhibition proposed in Fig. 3, because the corpus callosum presumably includes fibers that transmit specific, task-related information and others that transmit unrelated activity. In asymmetrically organized brains, a few callosal fibers that transmit specific information may suppress unrelated interhemispheric activity through the corpus callosum to such an extent that the overall callosal activity is reduced in comparison with the resting state. In more symmetrically organized brains, however, both hemispheres actively contribute to the mental processing. Thus, the bilateral execution of mental operations needs less energy-consuming activation of inhibitory neurons in the cortex but more specific information exchange through the corpus callosum. The callosal energy consumption because of the task-related information transmission may therefore exceed the reduction of unrelated transcallosal activity in individuals with minor functional lateralization.
Our results bear direct relevance to the debate on the ontogenic development and function of callosal fibers with respect to cerebral hemisphere dominance (Witelson, 1985; Kertesz et al., 1987). Studies of human and mammalian brains suggest that the more asymmetric a region is, the smaller the number of neurons and callosal axons (Galaburda et al., 1990; Aboitiz, 1992). This decrease of neuronal density is probably caused by specific neuronal degeneration, which adjusts the magnitude of each neuronal population to the size of functional needs of its projection field during brain development (Cowan et al., 1984). The concept shown in Fig. 3 entails the decrease of transcallosally projecting neurons in asymmetric cortical areas. The symmetric information transfer involves two pairs of neurons including two callosal axons, whereas the asymmetric information processing needs only one pair of excitatory neurons with one callosal axon. After the basic adjustment of neuronal populations during brain development, the mature brain apparently has the faculty to dynamically regulate neuronal activity according to the actual demands. This regulation principally has the same goal that the developmental changes had, the inhibition of inappropriate or excessive neuronal activity. Anatomic, electrophysiologic, and pharmacologic experiments, however, indicate that callosal fibers usually are glutaminergic and excitatory so that transcallosal inhibition most frequently is mediated by disynaptic transmission involving local inhibitory interneurons (Innocenti, 1994; Conti and Manzoni, 1994). Our findings suggest that this disynaptic collateral inhibition of interfering neuronal information is particularly important for the orchestration of functionally asymmetric regions in the speech-relevant cortex. Thus this study confirms Sperry's statement quoted at the beginning of this article that the two hemispheres can only speak as a unit when they inhibit unrelated activity.