
Abstract
Reactive astrocytes are thought to protect the penumbra during brain ischemia, but direct evidence has been lacking due to the absence of suitable experimental models. Previously, we generated mice deficient in two intermediate filament (IF) proteins, glial fibrillary acidic protein (GFAP) and vimentin, whose upregulation is the hallmark of reactive astrocytes. GFAP−/−Vim−/− mice exhibit attenuated posttraumatic reactive gliosis, improved integration of neural grafts, and posttraumatic regeneration. Seven days after middle cerebral artery (MCA) transection, infarct volume was 210 to 350% higher in GFAP−/−Vim−/− than in wild-type (WT) mice; GFAP−/−, Vim−/− and WT mice had the same infarct volume. Endothelin B receptor (ETBR) immunoreactivity was strong on cultured astrocytes and reactive astrocytes around infarct in WT mice but undetectable in GFAP−/−Vim−/− astrocytes. In WT astrocytes, ETBR colocalized extensively with bundles of IFs. GFAP−/−Vim−/− astrocytes showed attenuated endothelin-3-induced blockage of gap junctions. Total and glutamate transporter-1 (GLT-1)-mediated glutamate transport was lower in GFAP−/−Vim−/− than in WT mice. DNA array analysis and quantitative real-time PCR showed downregulation of plasminogen activator inhibitor-1 (PAI-1), an inhibitor of tissue plasminogen activator. Thus, reactive astrocytes have a protective role in brain ischemia, and the absence of astrocyte IFs is linked to changes in glutamate transport, ETBR-mediated control of gap junctions, and PAI-1 expression.
Introduction
Astroglial cells are the most abundant cells in the central nervous system (CNS) and are believed to play a major role in the brain and spinal cord pathologies. Although never proved directly, astrocytes are thought to exert a neuroprotective effect in stroke by shielding neurons from oxidative stress (Kraig et al, 1995). This hypothesis is based on the ability of astrocytes to engage in spatial buffering, to transport and metabolize amino acids, glucose, and other key molecules, and to upregulate antioxidants and free radical scavengers in the ischemic region.
Reactive gliosis is a response of astrocytes to CNS injury, including brain ischemia, mechanical trauma, neurodegenerative diseases, and tumors. Reactive astrocytes undergo changes in morphology and in their expression of a wide range of molecules (Eddleston and Mucke, 1993; Ridet et al, 1997). The hallmark of reactive gliosis, regardless of its origin, is increased expression of glial fibrillary acidic protein (GFAP) and vimentin in reactive astrocytes. Glial fibrillary acidic protein and vimentin are building blocks of intermediate filaments (IFs), which, together with microtubules and actin filaments, constitute the cytoskeleton.
We and others have generated GFAP−/− mice (Gomi et al, 1995; Liedtke et al, 1996; McCall et al, 1996; Pekny et al, 1995) and found that their astrocytes are free of IFs in the absence of any challenge, since vimentin requires GFAP to polymerize into filaments (Pekny et al, 1995). However, reactive astrocytes in GFAP−/− mice contain IFs because they also produce nestin, an IF protein that copolymerizes with vimentin in both GFAP−/− and wild-type (WT) reactive astrocytes (Eliasson et al, 1999; Pekny et al, 1998a). GFAP−/− mice exhibit normal healing after brain and spinal cord injury (Pekny et al, 1999b; Pekny et al, 1995). In mice lacking both GFAP and vimentin (GFAP−/−Vim−/−), however, reactive astrocytes are devoid of IFs, reflecting the inability of nestin to self-polymerize (Eliasson et al, 1999). As a result, reactive gliosis and glial scar formation after neurotrauma in the brain or spinal cord are attenuated (Pekny et al, 1999b; Pekny and Pekna, 2004). After neurotrauma, IF-free astrocytes of GFAP−/−Vim−/− mice reach comparable volume of tissue as WT astrocytes; however, GFAP−/−Vim−/−astrocytes show less prominent thickening of the main cellular processes (Wilhelmsson et al, 2004). Posttraumatic regeneration of neuronal synapses (Wilhelmsson et al, 2004) and integration of neural grafts (Kinouchi et al, 2003) were improved in GFAP−/−Vim−/− mice, despite more prominent synaptic loss at the initial stage after neurotrauma (Wilhelmsson et al, 2004).
In this study, we subjected GFAP−/−Vim−/−, GFAP−/−, and Vim−/− mice to brain ischemia induced by middle cerebral artery (MCA) transection by two independent paradigms. After 7 days of ischemia, infarct volumes were 2- to 3.5-fold larger in GFAP−/−Vim−/− mice than in WT, GFAP−/−, or Vim−/−mice. This increase correlated with reduced glutamate uptake in cortical slices from GFAP−/−Vim−/− mice, altered endothelin B receptor (ETBR)-mediated control of gap junctions, and downregulation of plasminogen activator inhibitor-1 (PAI-1) in GFAP−/−Vim−/− cultures. These findings suggest that reactive astrocytes have a protective role in brain ischemia.
Materials and methods
Mice
Single-mutant (GFAP−/− or Vim−/−), double-mutant (GFAP−/−Vim−/−), and WT control mice were maintained on a mixed C57Bl/129 genetic background as described (Eliasson et al, 1999; Pekny et al, 1999b). All mice were age-matched adults except for the preparation of astrocyteenriched cultures for which postnatal day 1 to 2 mice were used. The mice were kept in a barrier animal facility and fed ad libitum.
Induction of Ischemia: Proximal Middle Cerebral Artery Transection
Focal cerebral ischemia was induced in six WT and six GFAP−/−Vim−/− mice by proximal MCA transection as described (Fotheringham et al, 2000) with slight modifications. Mice were anesthetized with isoflurane in oxygen, and body temperature was maintained at 37°C with a heating pad. Under the operating microscope, the left MCA was exposed, occluded at two points by bipolar coagulation, and transected to ensure permanent disruption (Figure 1E). The proximal end of the MCA (A) was coagulated approximately 2.5 mm from the MCA/anterior cerebral artery branch (as determined after brain dissection). After surgery, mice were housed individually in cages that were placed on a heating pad. After 1 h, the mice were returned to their normal environment.
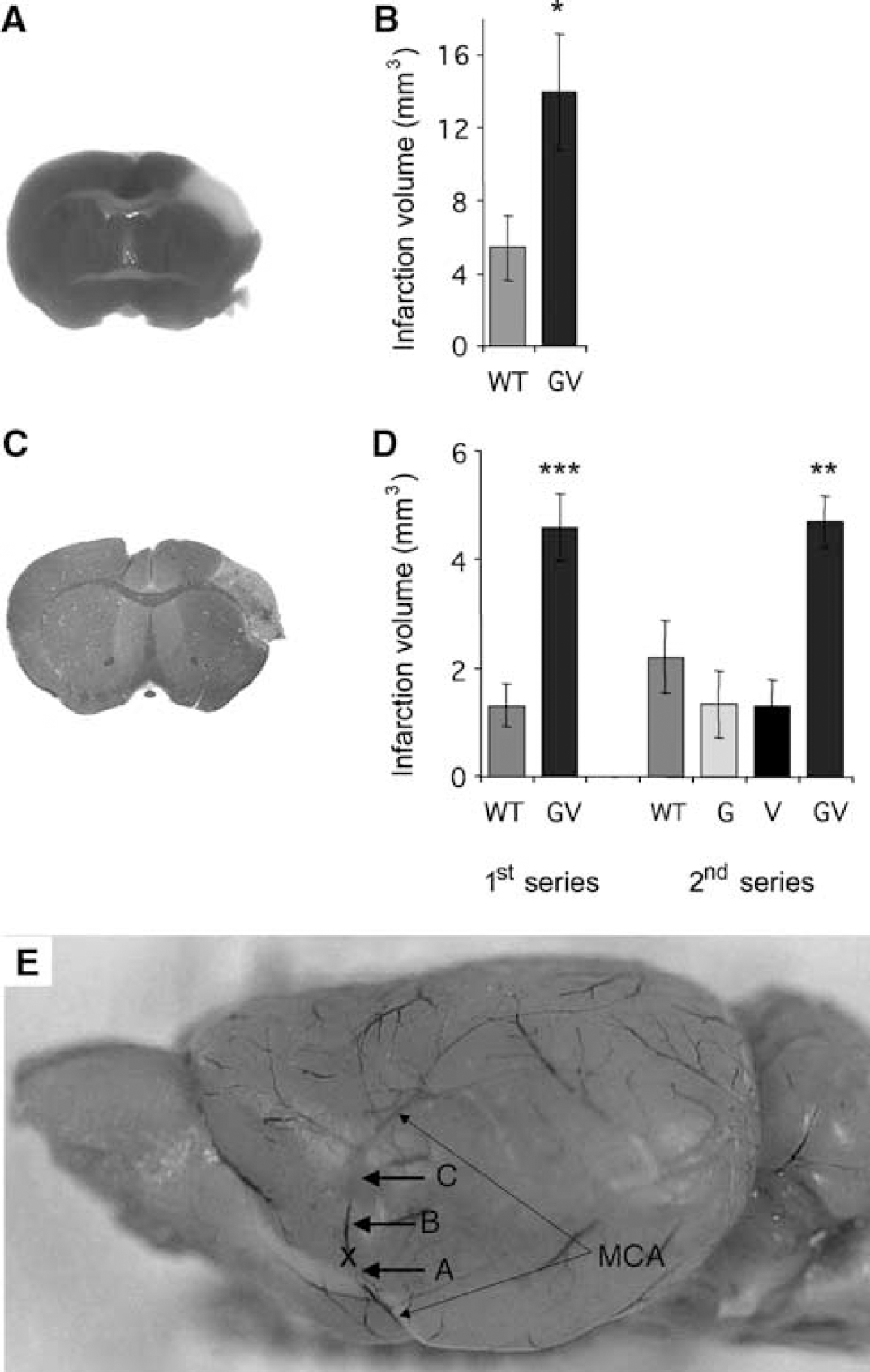
Infarct area is 2.5- to 3.5-fold larger in GFAP−/−Vim−/−(GV) than in WT mice 7 days after MCA transection. The infarct area was visualized by triphenyltetrazolium chloride and hematoxylin/erythrosin staining 7 days after proximal (A) and distal (
Induction of Ischemia: Distal Middle Cerebral Artery Transection
Distal MCA transection was performed as described (Nagai et al, 1999) in two independent series of experiments. One set of experiments was performed in 10 WT and 11 GFAP−/−Vim−/− mice. To determine if partial deficiency of astrocyte IFs or a deficiency of IFs in endothelial cells also affected the infarct volume, a second set of experiments was performed in nine WT, nine GFAP−/−, eight Vim−/−, and nine GFAP−/−Vim−/− mice. Briefly, mice were anesthetized by intraperitoneal injection of ketamine (75 mg/mL; Apharmo, Arnhem, The Netherlands) and xylazine (5 mg/mL; Bayer, Leverkusen, Germany), and body temperature was maintained at 37°C with a heating pad. Under the operating microscope, the MCA was ligated with a 10-0 suture Ethylon nylon thread (Ethylon, Neuilly, France) and transected distally (Figure 1E). After surgery, the mice were returned to their cages, which were placed on a heating pad (37°C) for 1 h.
Measurement of Infarct Volume 7 Days after Proximal Middle Cerebral Artery Transection
Deeply anesthetized mice were killed by decapitation. Fresh frontal brain slices (0.5 mm thick) were cut on a Vibratome, incubated for 30 mins in 0.125% triphenyltetrazolium chloride solution (T4375; Sigma-Aldrich, St Louis, MO, USA) in buffer containing 1.35% dimethylsulfoxide, 2 mmol/L MgCl2, 0.1 mol/L Na2HPO4, and 0.1 mol/L NaH2PO4 (pH 7.4) at 37°C, and fixed in 4% buffered formaldehyde (pH 7.4) (Fotheringham et al, 2000). In each slice, the infarct area in white matter was measured with a Nikon SMZ-U stereomicroscope and image analysis software (Easy Image, Bergström Instrument, Göteborg, Sweden). The infarct volume was calculated by integrating infarct areas on all adjacent brain slices with detectable infarction. The extent of brain edema was judged by comparing the areas of the ischemic and contralateral hemispheres.
Measurement of Infarct Volume 7 Days after Distal Middle Cerebral Artery Transection
Deeply anesthetized mice were perfused through the left ventricle with 4% phosphate-buffered formaldehyde (pH 7.4). The brains were postfixed overnight and embedded in paraffin. Frontal sections (8 μm thick) were stained with hematoxylin and erythrosine. Infarct areas were assessed by delineating the ischemic region, which could be clearly discriminated by its lighter appearance and high proportion of cells with pycnotic nuclei. The infarct area was measured on all sections (320 μm apart) on which it was detectable (4 to 10 sections per mouse) as described above, and the total infarct volume was calculated.
Blood Pressure and Heart Rate Monitoring
Blood pressure and heart rate were monitored in five WT and four GFAP−/−Vim−/− mice during the distal MCA transection. Under isoflurane anesthesia, the 10-0 suture Ethylon nylon thread (Ethylon, France) was passed under the MCA. The mouse was turned on its back, an incision was made in the neck, the left common carotid artery was exposed, and a 2.5F catheter was inserted and connected to a clinical fluid-filled discardable transducer (HLT0698; Fysicon, Oss, The Netherlands). Signal from the transducer was collected with an AD converter (PowerLab, ADInstruments, Chalgrove, UK). After measurement of baseline blood pressure and heart rate, the MCA was occluded by tightening the suture around the vessel. Blood pressure and heart rate were recorded 10 s, 15 mins, 30 mins, and 1 h after the MCA occlusion. After the operation, the mice were kept on a heating pad (37°C) for 1 h and were given free access to water and food. After 7 days, the mice were anesthetized with ketamine (75 mg/mL; Apharmo) and xylazine (5 mg/mL; Bayer) and perfused first with phosphate-buffered saline (PBS) for 7 mins and then with 4% phosphate-buffered formaldehyde (pH 7.4). The brains were postfixed overnight and embedded in paraffin.
Comparison of Cerebrovascular Architecture
The MCA territory was determined as described (Maeda et al, 1998). Briefly, the cerebrovasculature of four WT and four GFAP−/−Vim−/− mice, 8-month-old, was visualized by detection of endothelial cells with a rat anti-CD-31 antibody (1:200; BD Biosciences, San Jose, CA, USA) and a horseradish peroxidase-conjugated secondary antibody (P0162, 1:100; Dako, Glostrup, Denmark) according to standard immunohistochemical procedures; endogenous peroxidase activity was blocked by incubating the brains with PBS containing 0.6% H2O2 for 30 mins, and antibody incubations were performed at 4°C overnight. The distance between the dorsal midline and the line of anastomoses connecting the peripheral branches of the MCA and the anterior cerebral artery was measured at three points (2, 4, and 6 mm from the frontal pole) with the Nikon SMZ-U stereomicroscope and image analysis software (Easy Image).
Data Analysis
Data are expressed as mean±s.e.m. The two-tailed t-test was used for statistical analysis. Differences were considered significant at P < 0.05.
Immunohistochemistry
Coronal cryosections (35 μm), made from perfused and postfixed brains of four WT and three GFAP−/−Vim−/− mice, were incubated in 0.05% glycine in PBS for 1 h at room temperature and permeabilized overnight in PBS containing 0.5% Tween 20 and 1% bovine serum albumin at room temperature. S100β and ETBR were detected with rabbit antibodies against S100β (Z0311, 1:200; Dako) and ETBR (1:100; Alomone Labs, Jerusalem, Israel) and Alexa 488-conjugated anti-rabbit antibodies (1:500; Molecular Probes, Eugene, OR, USA). Glial fibrillary acidic protein and glutamine synthase (GS) were detected with mouse antibodies against GFAP (clone GA5, 1:100; Sigma-Aldrich) and GS (1:100; Chemicon Europe Ltd, Hampshire, UK), and Alexa 568-conjugated anti-mouse antibodies (1:500; Molecular Probes). Glutamate transporter-1 (GLT-1) was detected with guinea-pig antibodies against GLT-1 (1:100; Chemicon Europe Ltd.) and Alexa 488-conjugated anti-guinea pig antibodies (1:100; Molecular Probes). Microglia were detected with biotinylated tomato lectin (30 μg/mL; Sigma-Aldrich) and Cy3-conjugated streptavidin (1:100; Sigma-Aldrich). Antibodies were diluted in 1% bovine serum albumin and 0.01% Tween 20 in PBS. Cell nuclei were stained with ToPro-3 (1:1,000; Molecular Probes).
Astrocyte-Enriched Cultures
Primary astrocyte-enriched cultures were prepared from postnatal day 1 GFAP−/−Vim−/− and WT mice as described (Pekny et al, 1998a). The cultures for immunocytochemistry were grown in 16-chamber slides (Nalge Nunc International, Naperville, IL, USA) in Dulbecco's modified Eagle's medium (D5671; Sigma-Aldrich) containing 10% fetal calf serum, 2 mmol/L L-glutamine, and penicillin—streptomycin (Invitrogen, Paisley, UK). Cultures for quantitative real-time PCR analysis of mRNA were maintained in 10% fetal calf serum (ETBR and connexin 43 (Cx43)) or 1 or 10% fetal calf serum (PAI-1).
Immunocytochemistry
Cells were fixed in methanol at −20°C for 5 mins and washed with PBS. After nonspecific binding was blocked with 5% normal goat serum (Dako) for 15 mins at room temperature, cells were incubated with rabbit antibodies against ETBRs (1:100; Alomone Labs) and mouse antibodies against GFAP (clone GA5, 1:100; Sigma-Aldrich) and then with Alexa 488-conjugated anti-rabbit and Alexa 568-conjugated anti-mouse antibodies (1:500; Molecular Probes). All antibodies were diluted in PBS.
Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis and Immunoblotting for Endothelin B Receptors
Medium was removed from confluent cultures of astrocytes (10 to 14 days after plating). After two washes with PBS, cells were harvested by scraping into sodium dodecyl sulfate-extraction buffer consisting of protease inhibitor cocktail (Roche, Mannheim, Germany), 1 mmol/L Na+ orthovanadate, and 1 mmol/L phenylmethanesulfonyl in 2% sodium dodecyl sulfate, and the cell lysate was frozen at −80°C. Thawed lysates were sonicated for 5 secs, and the protein content was determined with the Bicinchoninic Acid Protein Assay (Sigma-Aldrich). Protein (50 μg) was separated on 4 to 12% bis-Tris gels (Invitrogen, Carlsbad, CA, USA) and transferred to a nitrocellulose membrane. The membrane was stained with Ponceau S solution (Sigma-Aldrich) to control for equal protein loading. ETBRs were detected with polyclonal rabbit antibodies (1:500; Alomone Labs), and β-actin (internal loading control) was detected with a mouse monoclonal antibody (1:1,000; Abcam, Cambridge, UK). To enhance the signal for ETBR, the immunoblot was incubated with biotin-conjugated monoclonal anti-rabbit immunoglobulins (Sigma-Aldrich) and probed with peroxidase-conjugated streptavidin (Dako) or with peroxidase-conjugated donkey anti-mouse immunoglobulins for β-actin detection (Jackson Immunoresearch, Suffolk, UK). Immunoreactivity was visualized by chemiluminescence and quantified with Multi Gauge v2.3 software (Fuji Photo Film Co., Ltd, Tokyo, Japan). Band densities were compared by one-way analysis of variance followed by Bonferroni's multiple comparison test performed with the SPSS statistical package v11.5.1 (SPSS, Chicago, IL, USA).
Reverse Transcription and Quantitative Real-Time PCR
cDNA was generated using the iScript cDNA Synthesis Kit (Bio-Rad Laboratories, Hercules, CA, USA) with a mixture of random hexamers and oligo(dT) primers, according to the manufacturer's instructions, except that for ETBR and Cx43 expression analyses, the incubation time at 42°C was increased from 30 to 60 mins. The reverse transcription was run in duplicate in 10-μL reactions (Stahlberg et al, 2004) using 1.5 μg of total RNA extracted from primary astrocyte cultures as described below. The cultures were prepared from four WT and four GFAP−/−Vim−/− mice (for ETBR and Cx43), and five WT and three GFAP−/−Vim−/−mice (for PAI-1). Gene-specific SYBR-Green I-based PCR assays were designed for ETBR (GenBank accession number NM_007904), PAI-1 (GenBank accession number M33960), and Cx43 (GenBank accession number M63801). Formation of expected PCR products was confirmed with agarose gel electrophoresis and melting curve analysis. The primer sequences were as follows: 5′-AAGAATGCCCAAGAGAAAAC-3′ (forward) and 5′-AAAAAGGAAGGAAGGAAAATC-3′ (reverse) for ETBR; 5′-CAGACAATGGAAGGGCAACA-3′ (forward) and 5′-GCGGGCTGAGATGACAAA-3′ (reverse) for PAI-1; and 5′-ACCCAACAGCAGCAGACTT-3′ (forward) and 5′-ACCGACAGCCACACCTTC-3′ (reverse) for Cx43. Real-time PCR experiments were run on a Rotor-Gene 3000 (Corbett Research, Sydney, Australia) and were analyzed as described (Stahlberg et al, 2004). All gene expression data were normalized to total RNA concentration. The statistical significance of differences between GFAP−/−Vim−/− and WT mice was tested with two-tailed t-tests. Polymerase chain reaction run conditions and thermocycler programs are available on request.
Scrape Loading/Dye Transfer
Astrocyte gap-junctional communication (AGJC) was assessed by using the scrape loading/dye transfer technique as described (Blomstrand et al, 1999). In brief, confluent astrocyte cultures from five WT and four GFAP−/−Vim−/− mice were incubated for 7 mins in HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid)-buffered salt solution (137 mmol/L NaCl, 5.4 mmol/L KCl, 0.41 mmol/L MgSO4, 0.49 mmol/L MgCl2, 1.26 mmol/L CaCl2, 0.64 mmol/L KH2PO4, 3 mmol/L NaHCO3, 5.5 mmol/L glucose, and 20 mmol/L HEPES (pH 7.4)), with or without a nonsaturating dose of 10 nmol/L of endothelin-3 in this and subsequent steps, and followed by four quick washes in Ca2+-free buffer. Lucifer yellow (0.1% w/v) in Ca2+-free buffer was added, and two parallel scrapes were performed with a scalpel. The Lucifer yellow was removed after 1 min, and Ca2+-containing buffer was added. 8 mins after scraping, four images from each culture were captured with a Nikon Optiphot 2 equipped with a Hamamatsu C5810 chilled three-chip color charge-coupled device camera. The dye spread was quantitated by using Easy Image Analysis 2000 (Tekno Optik AB, Stockholm, Sweden) to measure the fraction of a predefined area occupied by Lucifer yellow-positive cells.
Glutamate Uptake
Fresh brain slices microdissected from five WT and nine GFAP−/−Vim−/− mice were homogenized in 5 mmol/L Tris/ 320 mmol/L sucrose with a protease inhibitor cocktail (Complete; Roche, Indianapolis, IN, USA), using a handheld pellet pistol. Samples were then washed three times in ice-cold PBS (pH 7.4) and resuspended in Na+-containing Krebs buffer (120 mmol/L NaCl, 25 mmol/L Tris—HCl, pH 7.4, 5 mmol/L KCl, 2 mmol/L CaCl2, 1 mmol/ L KH2PO4, 1 mmol/L MgSO4, and 10% glucose) or in Na+-free Krebs (120 mmol/L choline-Cl substituted for NaCl). The samples were then incubated with 10 μmol/L 3H-glutamate for 4 mins at 37°C, harvested with a tissue harvester (Brandel, Gaithersburg, MD, USA) on a Whatman GF/B paper (FPD-100; Brandel), and washed three times in Tris buffer (pH 7.4). Radioactivity was determined by standard scintillation counting. Mean Na+-dependent uptake was determined in duplicate by subtracting the mean uptake in a Na+-free duplicate. To determine the contribution of GLT-1-mediated transport, a separate set of samples was also incubated with 500 μmol/L dihydrokainate and 10 μmol/L 3H-glutamate for 4 mins at 37°C, harvested, and washed three times in Tris buffer (pH 7.4) on the Whatman GF/B paper using a tissue harvester. Radioactivity and mean Na+-dependent uptake were determined as described above.
Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis and Immunoblotting for Glutamate Transporter-1
Brain slices (approximately 2 × 2 mm) taken from the cortex of the left hemisphere of six WT and nine GFAP−/−Vim−/− mice were homogenized by sonication in Tris-EDTA (ethylenediaminetetraacetic acid) buffer containing protease inhibitors (aprotinin, pepstatin, and leupeptin). Samples were stored at −70°C until use. Aliquots of homogenized samples (1 to 20 μg protein) were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (8% polyacrylamide gels) and transferred to nitrocellulose membranes (Hybond ECL, Amersham, Buckinghamshire, UK) by electroblotting (100 V, 60 mins). Western blot analysis was performed as described (Rothstein et al, 1994). In brief, blots were blocked and washed in 50 mmol/L Tris-buffered saline containing 5% nonfat milk and 0.5% Tween 20, and probed with affinity-purified polyclonal antibodies specific for GLT-1 (1:10,000; C terminus-directed antibody) using horseradish peroxidase-conjugated donkey anti-rabbit IgG (1:5,000; Amersham) as secondary antibody. The blots were also probed for actin (1:2,000) as a protein-loading standard. In all cases, protein levels varied by < 10% between lanes. Immunoreactivity was visualized by enhanced chemiluminescence and quantified densitometrically with Quantity One v4.0 software (Bio-Rad Laboratories).
DNA Array Analysis
RNA extraction, DNase treatment, poly (A+) RNA enrichment, probe synthesis, column chromatography, and hybridization of the probes to the array were performed as described in the Atlas Pure Total RNA Labeling System User Manual and the Atlas cDNA Expression Arrays User Manual (Clontech Laboratories, Palo Alto, CA, USA). Confluent astrocyte cultures were harvested by scraping into RNase-free PBS and centrifuged at 500g for 5 mins at 4°C. The arrays were exposed to phosphor screens (Molecular Dynamics, Buckinghamshire, UK) overnight. The screens were scanned in a Storm instrument (Storm 820, Molecular Dynamics), and the images were analyzed with AtlasImage 1.5 (Clontech Laboratories).
Results
Larger Infarct Volume in GFAP–/–Vim–/– than WT Mice after Proximal Middle Cerebral Artery Transection
Seven days after proximal MCA transection, infarct volume was 257% higher in GFAP−/−Vim−/− mice than in WT controls (14.0±3.20 vs 5.43±1.81 mm3, P < 0.05; Figures 1A and 1B). No difference was found in the extent of brain edema (1.00±0.08 vs 1.04±0.08, respectively).
Larger Infarct Volume in GFAP–/–Vim–/– than WT Mice after Distal Middle Cerebral Artery Transection
To determine if the transection point affects the difference in the infarct volume between WT and GFAP−/−Vim−/− mice, we transected the MCA more distally (Figure 1E). In the first set of experiments, the infarct volume was 351% larger in GFAP−/−Vim−/− than in WT mice 7 days after distal MCA transection (4.60±0.62 vs 1.31±0.40 mm3, P < 0.001; Figures 1C and 1D). To determine if partial deficiency of IFs in astrocytes or deficiency of IFs in endothelial cells also affected the infarct volume, single GFAP−/− and Vim−/− mice were included in a second experimental series. The infarct volume was 214% larger in GFAP−/−Vim−/− than in WT mice (4.71±0.62 vs 2.20±0.40 mm3, P < 0.01; Figure 1D), and there were no significant differences in the infarct volume between GFAP−/− and Vim−/− mice (1.34±0.62 and 1.32±0.46 mm3, respectively) and WT mice (Figure 1D).
Median Blood Pressure and Heart Rate and Cerebrovascular Architecture are not Altered in GFAP–/–Vim–/– Mice
To address a possible primary effect of the absence of GFAP and Vim on blood pressure and cerebrovascular architecture, we recorded blood pressure and heart rate during and after the MCA transection and compared the anastomoses between branches of the MCA and the anterior cerebral artery. During the MCA transection and for 60 mins thereafter, mean blood pressure and heart rate did not differ in WT and GFAP−/−Vim−/− mice (data not shown). The lines of anastomoses between branches of the MCA and the anterior cerebral artery were comparable in WT and GFAP−/−Vim−/− mice (Figure 2).
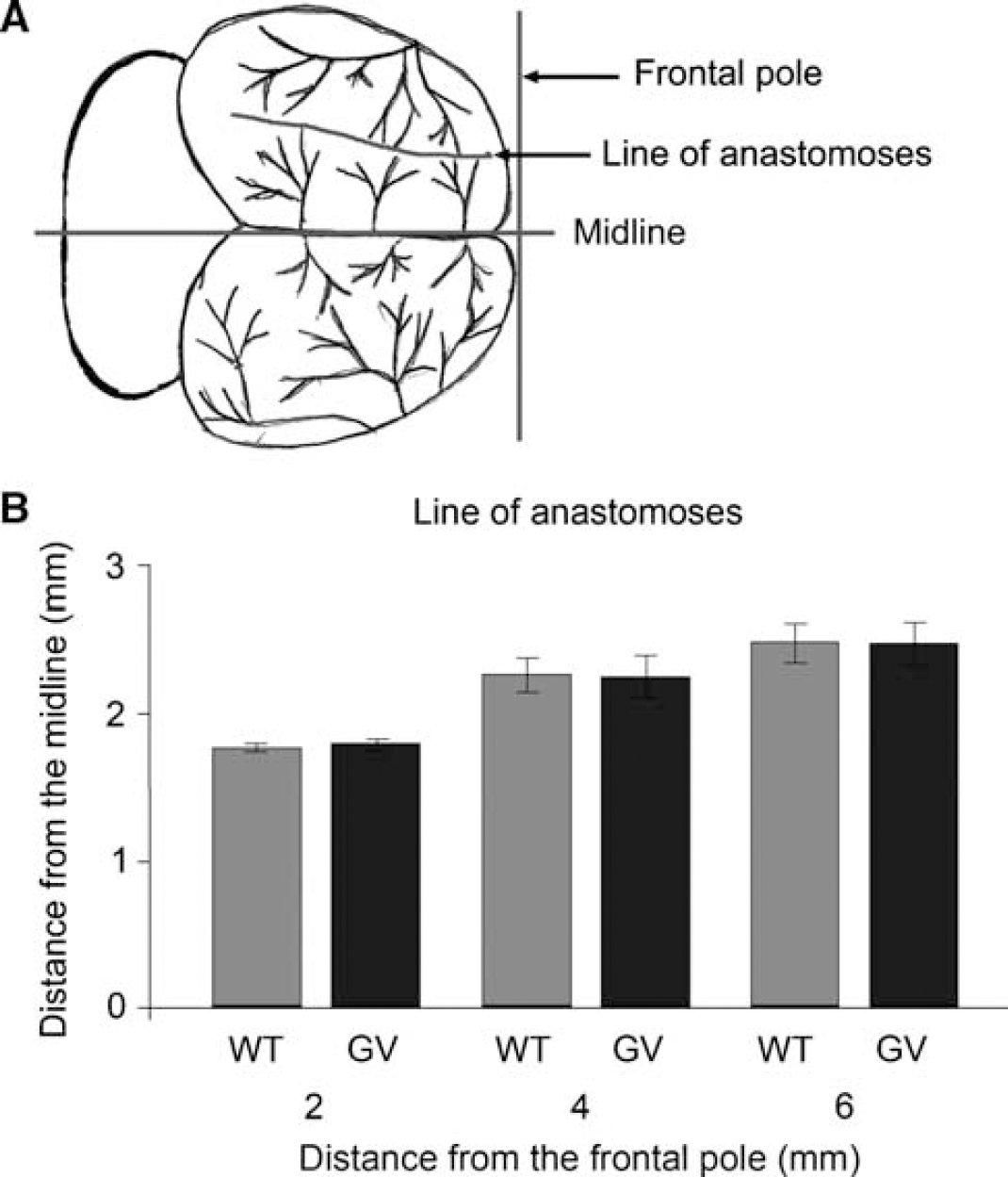
The territory supplied by the MCA is comparable in WT and GFAP−/−Vim−/− (GV) mice. Comparison of the line of anastomoses (
Normal Number of Astrocytes and Microglia in the Penumbra of GFAP–/–Vim–/– Mice
Next, we examined the penumbra and the infarct area histologically. Except for the larger infarct area in GFAP−/−Vim−/− mice, hematoxylin/erythrosine staining showed no difference in the appearance of the penumbra and the infarct region between WT and GFAP−/−Vim−/− mice. Immunostaining for S100β revealed comparable numbers of astrocytes in the two groups (62.13±3.29 vs 57.33±3.44 per 0.25 mm2 for WT and GFAP−/−Vim−/− mice, respectively; P = 0.37), although GFAP−/−Vim−/− astrocytes had less prominent hypertrophy of the main cellular processes (Figures 3A and 3B). Comparable numbers of astrocytes in the two groups were also confirmed by GS immunostaining (data not shown). Tomato lectin staining showed no differences in the appearance, distribution, and number of microglial cells in the penumbra and the infarct region between WT and GFAP−/−Vim−/− mice (54.5±2.84 vs 61.33±5.24 per 0.0625 mm2, respectively; P = 0.27 and data not shown).
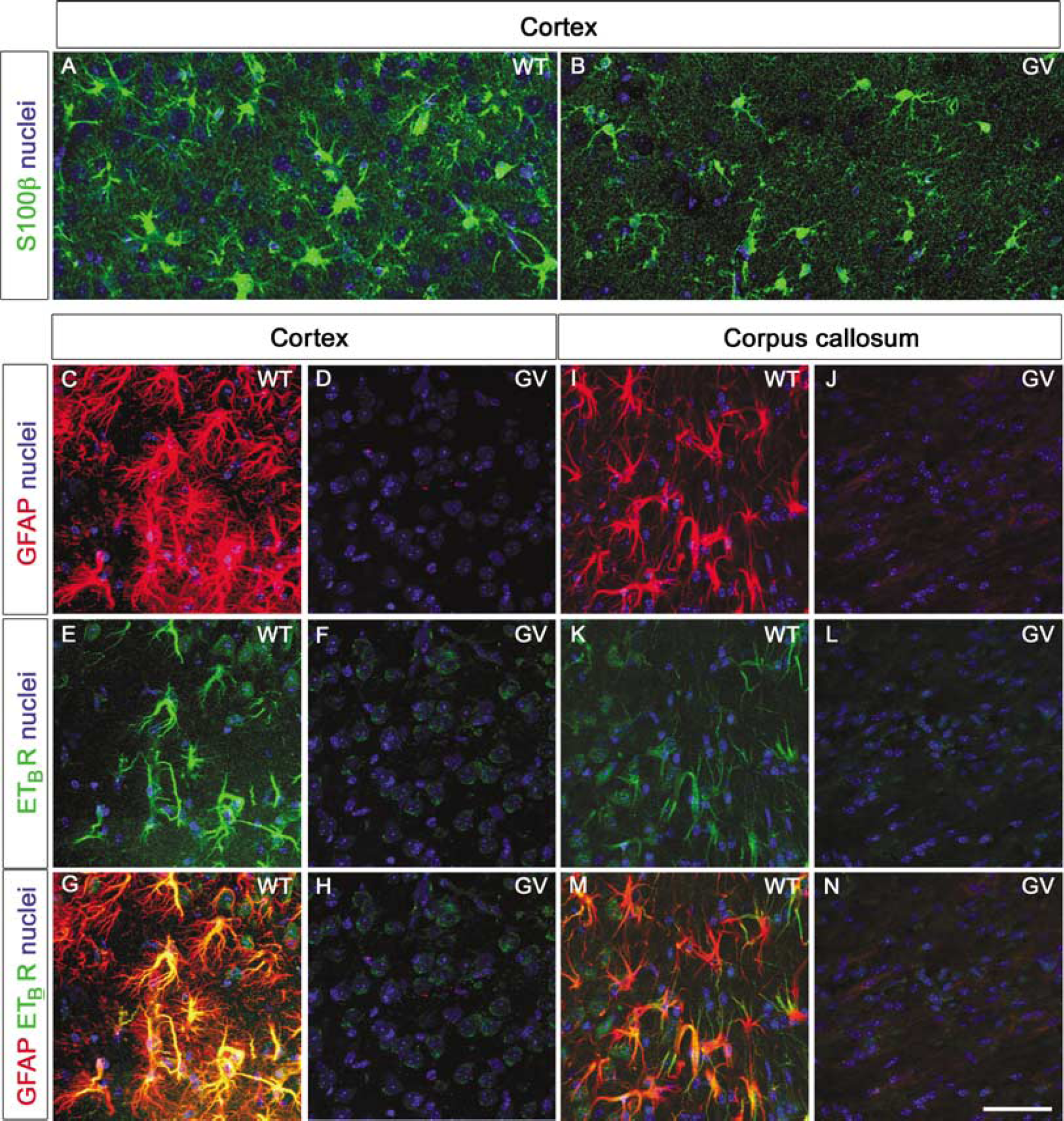
Astrocytes around the ischemic lesion show ETBR immunoreactivity in WT but not GFAP−/−Vim−/− (GV) mice. (
Absence of Endothelin B Receptor-Immunoreactivity in Astrocytes in the Ischemic Penumbra of GFAP–/–Vim–/– Mice
Endothelin B receptor expression by astrocytes in the injured CNS was proposed as one of the steps leading to astrocyte activation and reactive gliosis (Koyama et al, 1999). In the ischemic penumbra and in the corpus callosum 7 days after MCA transection, ETBRs were highly expressed on reactive astrocytes in WT mice (Figures 3E and 3K) but were essentially undetectable on astrocytes of GFAP−/−Vim−/− mice (Figures 3F and 3L). Similarly, in WT astrocytes in cultures prepared from postnatal day 1 mouse brains, ETBRs were readily detectable and the ETBR immunoreactivity exhibited a characteristic filamentous appearance (Figure 4). In contrast, ETBR immunoreactivity was undetectable in the cytoplasm of GFAP−/−Vim−/−astrocytes (Figure 4). Interestingly, numerous GFAP−/−Vim−/− astrocytes exhibited a granular ETBR immunoreactivity in the cell nucleus (Figure 4). This finding suggests that IFs are required for the production, stability, or distribution of ETBRs in reactive astrocytes.
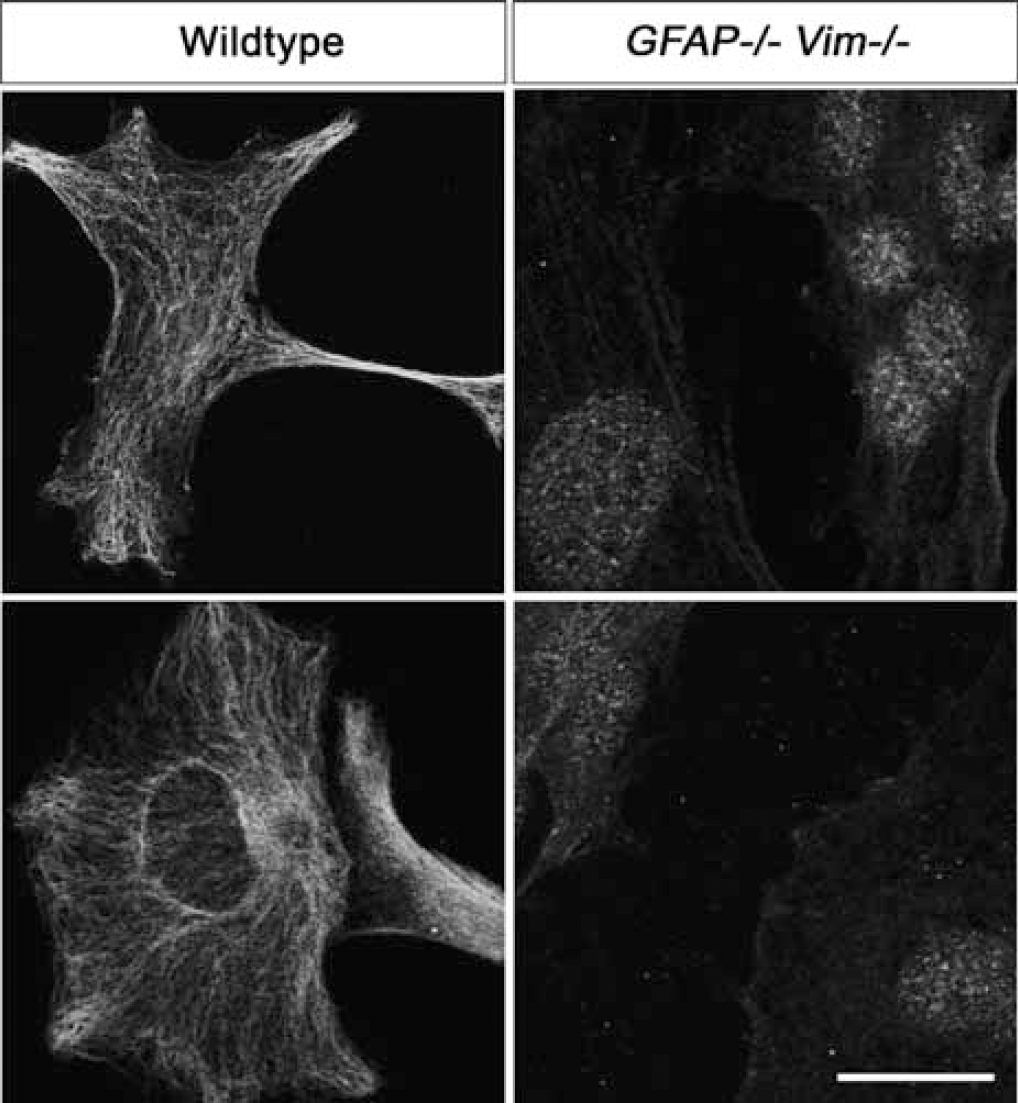
ETBR immunoreactivity had a filamentous appearance in the cytoplasm of cultured WT astrocytes but was absent in the cytoplasm of GFAP−/−Vim−/− astrocytes, where it was often confined to the cell nuclei. Green, ETBR. Scale bar = 50 μm.
Endothelin B Receptor Immunoreactivity Colocalizes with Intermediate Filament Bundles in Reactive Astrocytes
To investigate further the relationship between ETBR and astrocyte IFs, we analyzed their distributions in cultured astrocytes by laser-scanning confocal microscopy. This analysis revealed remarkable colocalization between bundles of IFs and ETBR immunoreactivity in WT astrocytes, with the ETBR immunoreactivity decorating bundles of IFs (Figure 5).
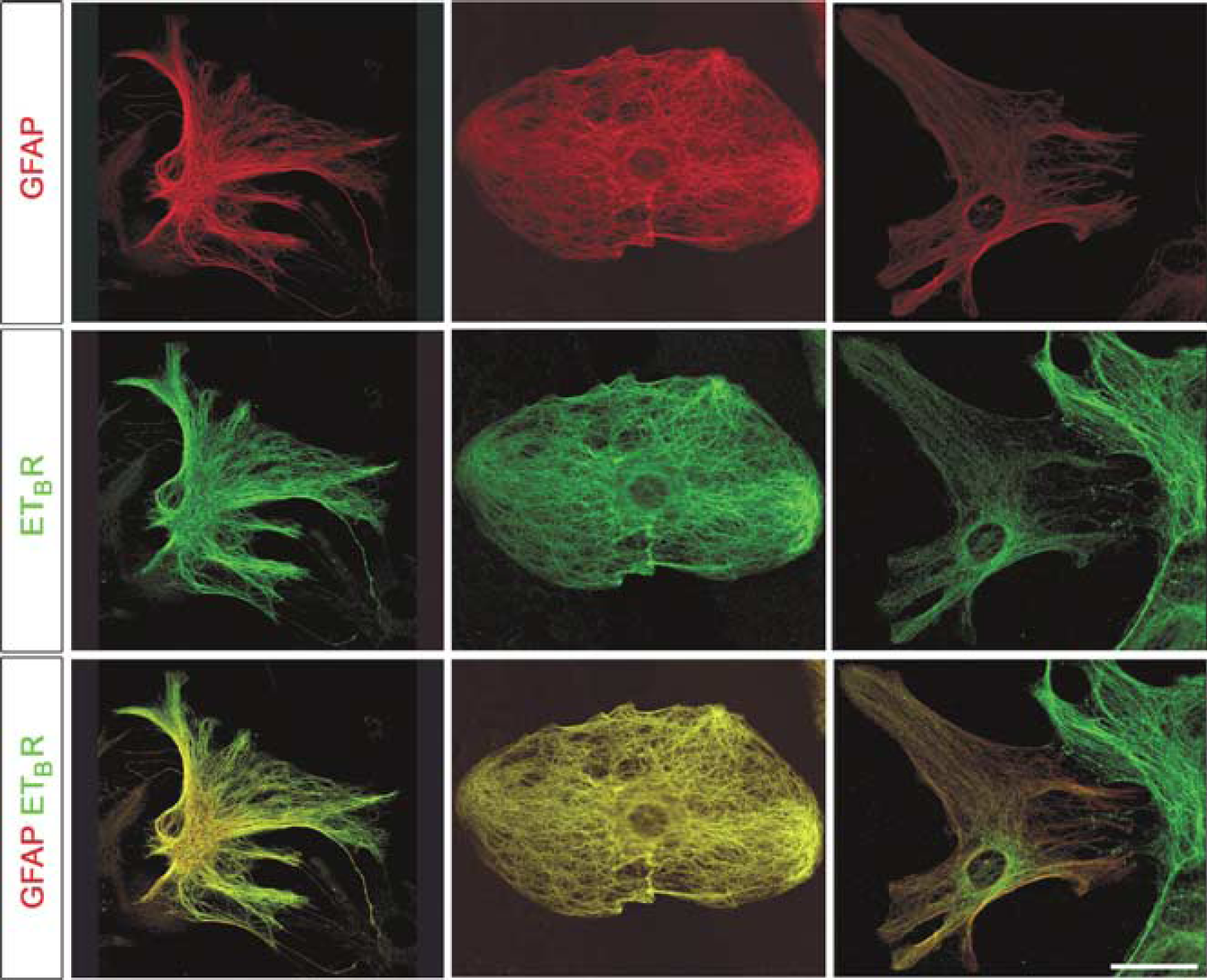
ETBR and bundles of IFs colocalize in cultured WT astrocytes. Laser-scanning confocal microscopy revealed a filamentous appearance of ETBR immunostaining, which colocalized with GFAP-positive bundles of IFs. Red, GFAP; green, ETBR. Scale bar = 20 μm.
Intermediate Filaments Determine Endothelin B Receptor Distribution in Reactive Astrocytes
To determine if IFs are required for the production, stability, or distribution of ETBR in reactive astrocytes, we performed quantitative real-time PCR and Western blot analyses on cultured astrocytes from GFAP−/−Vim−/− and WT mice. Endothelin B receptor mRNA levels were 43% higher in GFAP−/−Vim−/− than in WT astrocytes (P < 0.05), but the amounts of ETBR protein were comparable (Figure 6). Thus, in the absence of IFs, astrocytes contain a normal amount of ETBRs, which do not associate with IFs but seem to be distributed throughout the cell.
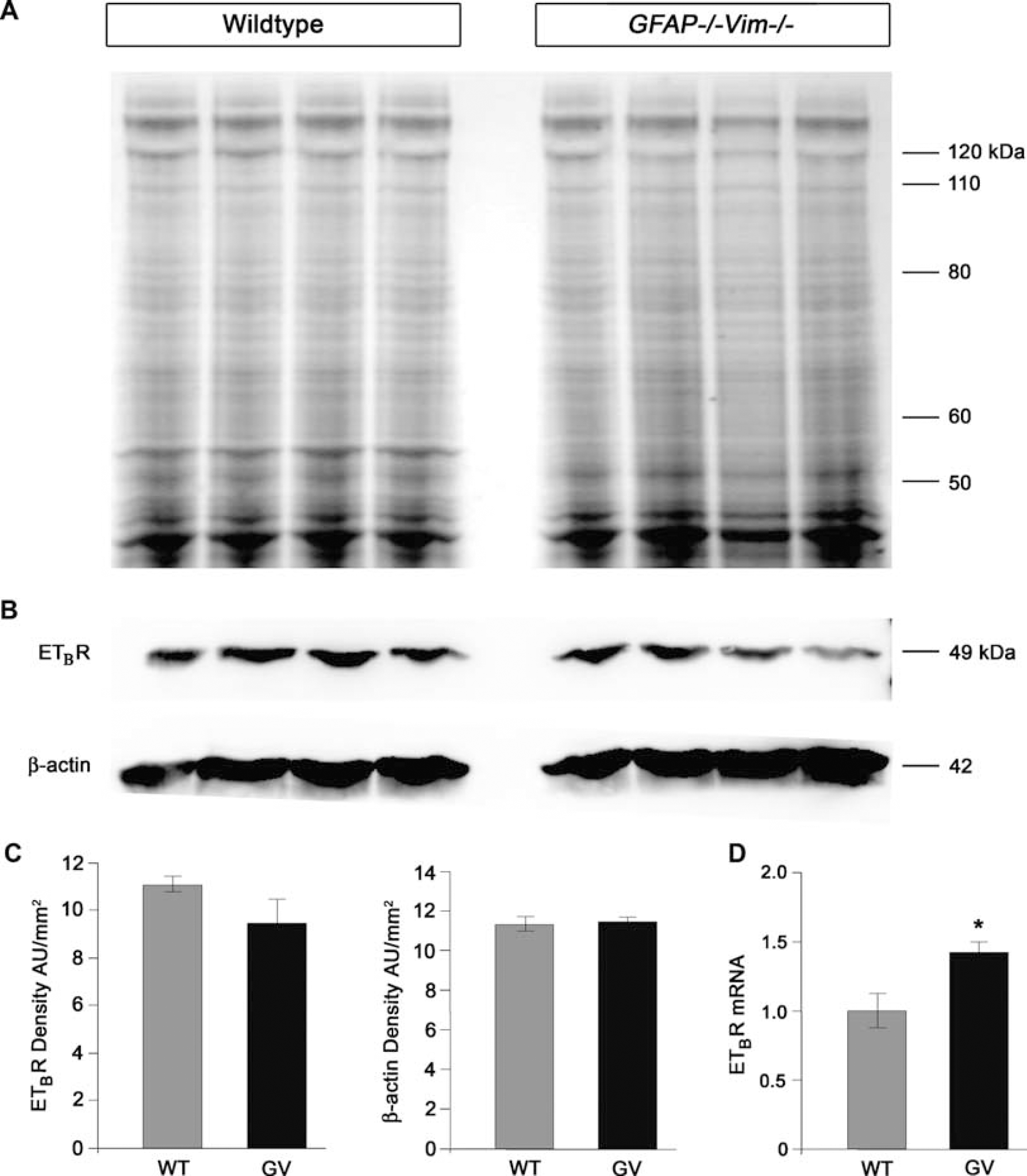
ETBR protein levels are comparable in WT and GFAF−/−Vim−/− (GV) cultured astrocytes. (
Endothelin-3-Induced Blockage of Gap Junctions is Attenuated in GFAP–/–Vim–/– Astrocytes
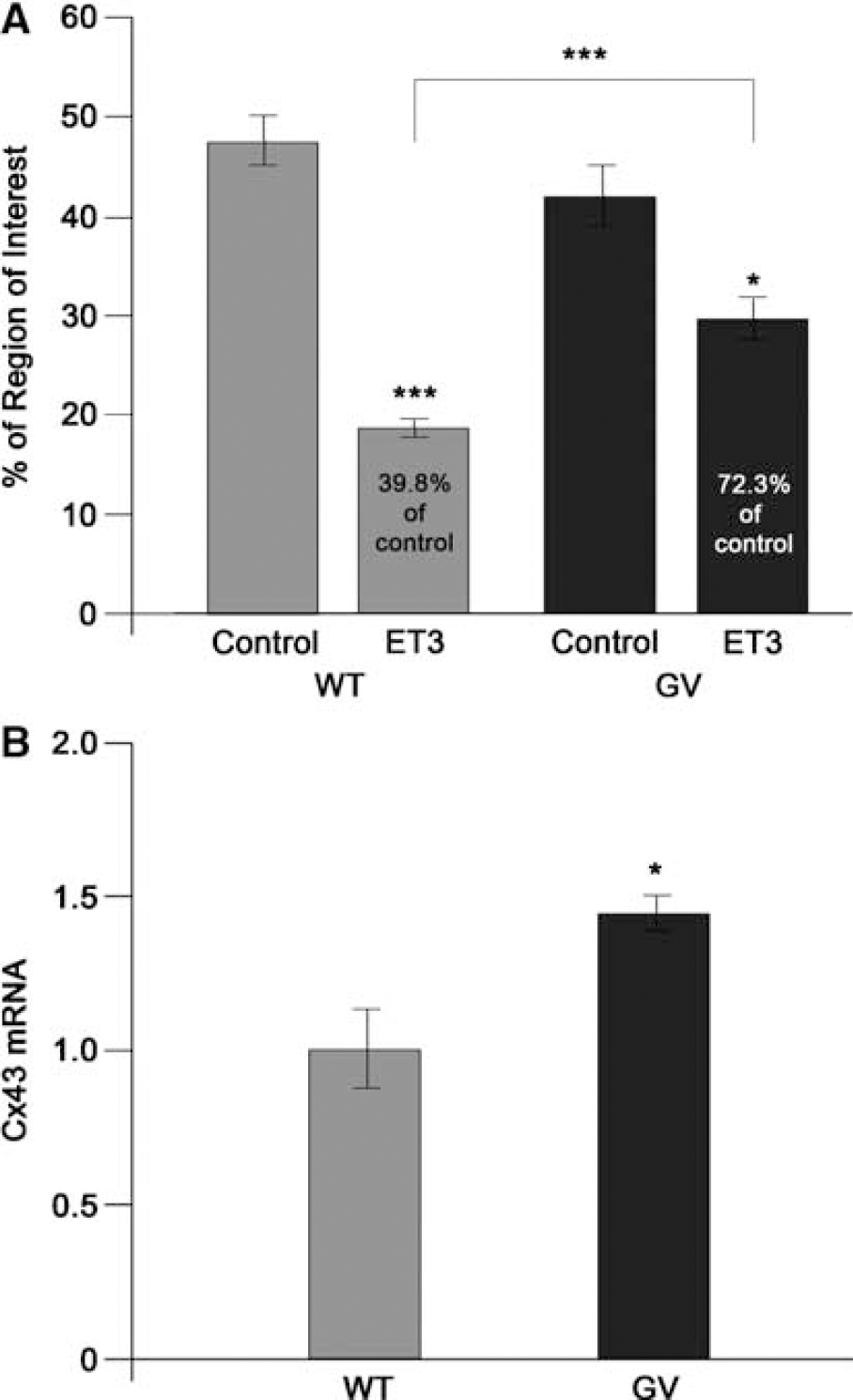
Endothelins are well-known blockers of astrocyte gap-junctional communication (AGJC) (Blomstrand et al, 1999). To determine if the altered cytoplasmic distribution of ETBRs affects AGJC and thus the function of the astrocyte syncytium, we stimulated ETBRs with the selective ligand endothelin-3. No significant difference in basal AGJC was observed between WT and GFAP−/−Vim−/− astrocytes, but endothelin-3-mediated inhibition of AGJC was less prominent in GFAP−/−Vim−/− than in WT astrocytes (72.3±8.9 vs 39.8±2.6% of control levels; P < 0.01; Figure 7A). Gap junctions are made of connexins, with Cx43 being the most predominant gap junction protein in astrocytes (Giaume and McCarthy, 1996). Using quantitative real-time PCR, we found that Cx43 mRNA expression was 44% higher in GFAP−/−Vim−/− in than WT astrocytes (P < 0.05; Figure 7B), further linking astrocyte IFs and AGJC.
Reduction in Glutamate Transport in GFAP–/–Vim–/–Mice
The ability of astrocytes to remove glutamate released by neurons was proposed to reduce infarct size by limiting the excitotoxic cell death (reviewed in Nedergaard and Dirnagl, 2005). To determine if glutamate transport is altered in GFAP−/−Vim−/− mice, we examined total glutamate uptake and glutamate uptake mediated by GLT-1, the primary glutamate transporter in astrocytes, in freshly dissected hemicortices of GFAP−/−Vim−/− and WT mice. Mean total glutamate uptake in GFAP−/−Vim−/− mice was 44% lower than in WT controls (1.99±0.79 vs 3.53 ± 0.17 nmol/mg per min; P < 0.05). Similarly, GLT-1-dependent transport was 50% lower than in WT mice (1.38±0.78 vs 2.85±0.29 nmol/mg per min, P < 0.05; Figure 8A). Western blot analysis showed comparable amounts of GLT-1 in WT and GFAP−/−Vim−/− cerebral cortex (22±3 vs 20±4U; Figures 8B and 8C) and immunohistochemical analyses of the ischemic penumbra showed less prominent GLT-1 immunostaining in astrocytes of GFAP−/−Vim−/− compared with WT mice (Figure 8D). Thus, astrocytes of GFAP−/−Vim−/− mice are less capable of removing glutamate from the ischemic brain tissue.
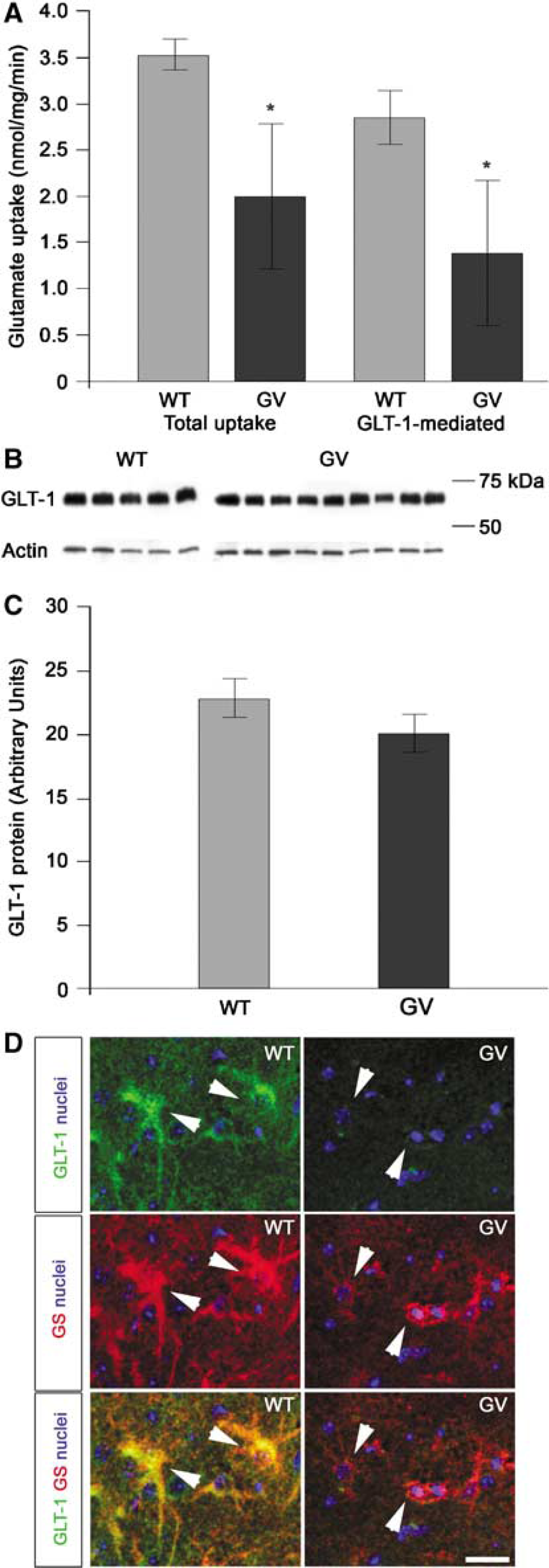
Total and GLT-1-mediated glutamate uptake is decreased in the cortex of GFAP−/−Vim−/− (GV) mice. (
Plasminogen Activator Inhibitor-1 is Downregulated in GFAP–/–Vim–/– Astrocytes
To address further, at a molecular level, the effect of attenuated reactive gliosis, we performed microarray analysis with the Atlas cDNA Expression Arrays (Clontech Laboratories). The expression of 1,200 genes was analyzed in primary astrocyte cultures derived from WT and GFAP−/−Vim−/− mice and maintained in the presence of 10% serum. Only a single gene, PAI-1, an inhibitor of tissue plasminogen activator (tPA), fulfilled the criteria of a threefold or higher downregulation in GFAP−/−Vim−/− than in WT astrocytes (the ratio was 0.3 for GFAP−/−Vim−/−/WT astrocyte cultures). Next, we performed quantitative real-time PCR analysis of PAI-1 mRNA in primary GFAP−/−Vim−/− and WT astrocytes maintained in the presence of either 1 or 10% serum, the latter mimicking some aspects of reactive gliosis. Compared with WT, GFAP−/−Vim−/− astrocytes showed a 69% reduction in PAI-1 mRNA in the presence of 1% serum (P < 0.05), and an 86% reduction in 10% serum (P < 0.01; Figure 9).
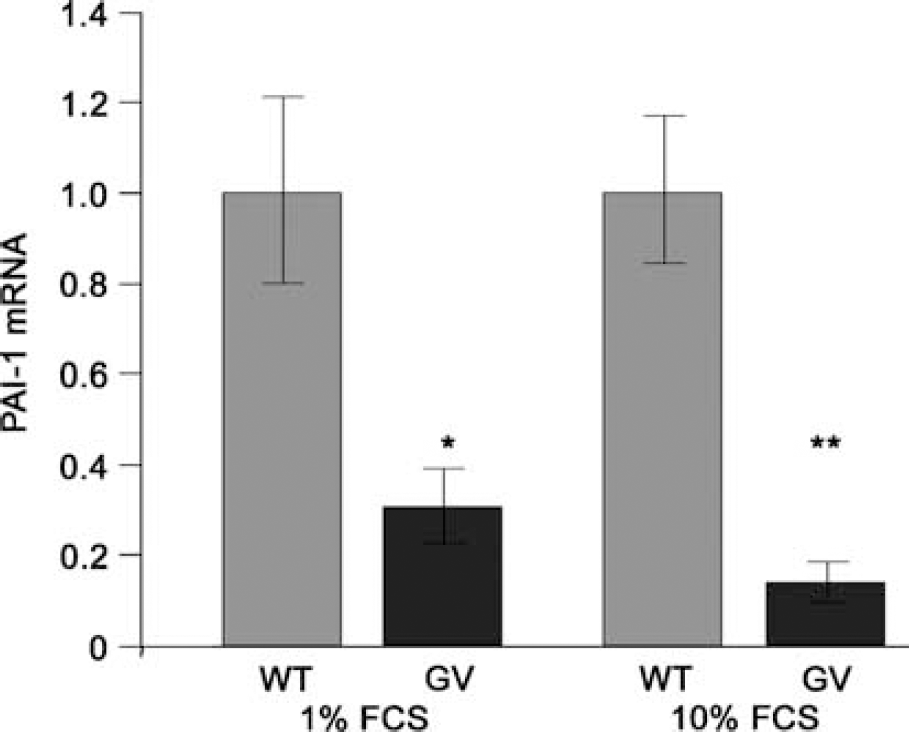
Plasminogen activator inhibitor-1 mRNA levels are lower in GFAP−/−Vim−/− (GV) than in WT astrocytes cultured in the presence of 1 or 10% fetal calf serum (**P < 0.01 and *P < 0.05). Values are mean±s.e.m.
Discussion
In the absence of a suitable experimental model, direct proof that astrocytes have a protective effect in the ischemic brain has largely been lacking. Attempts to generate mammalian models in which astrocytes would be globally or regionally eliminated resulted in phenotypes too severe or complex for studying the role of astrocytes in the pathogenesis of CNS diseases. In mice expressing herpes simplex virus thymidine kinase from the GFAP promoter, ablation of dividing astroglial cells by treatment with ganciclovir during early postnatal development led to severe developmental abnormalities (Delaney et al, 1996). The same approach was applied in adult mice to selectively eliminate the subpopulation of dividing reactive astrocytes after neurotrauma and resulted in a massive invasion of leukocytes in response to dying astrocytes, increased neurodegeneration, and failed blood—brain barrier (BBB) repair (Bush et al, 1999), implying neuroprotective activities of astrocytes in the lesioned area (Faulkner et al, 2004).
Upregulation of IFs is a hallmark of reactive gliosis after trauma or stroke and is a key step in astrocyte activation (Pekny et al, 1999b). To address the role of reactive astrocytes in stroke, we subjected GFAP−/−, Vim−/−, and GFAP−/−Vim−/− mice, which lack one or both of the key components of IFs in reactive astrocytes, to proximal or distal MCA transection. In three independent experiments, GFAP−/−Vim−/− mice had 2- to 3.5-fold larger infarct volumes after 7 days of ischemia than WT controls. The infarct volumes in GFAP−/− or Vim−/− mice were not significantly different from those in WT mice. The increased infarct volumes in GFAP−/−Vim−/− mice were linked to an altered distribution of ETBR, less efficient ETBR-mediated inhibition of AGJC, reduced ability to take up glutamate, and lower levels of tPA inhibitor PAI-1 in IF-free GFAP−/−Vim−/− astrocytes. These results provide evidence that reactive astrocytes have a protective role in brain ischemia.
In GFAP−/− mice, reactive astrocytes contain reduced amounts of IFs, which are composed of vimentin and nestin (Eliasson et al, 1999; Pekny et al, 1998a). Reactive astrocytes in Vim−/− mice also contain reduced amounts of IFs, and the IF bundles are abnormally compact, reflecting reduced space between individual filaments (Eliasson et al, 1999). Despite these abnormalities, reactive astrocytes in these single-mutant mice were not compromised with regard to their function in stroke. However, the complete absence of IFs in reactive astrocytes in GFAP−/−Vim−/− mice profoundly affected infarct volume. These results are consistent with the findings that healing after brain or spinal cord trauma was prolonged and synaptic loss was increased in GFAP−/−Vim−/− mice, which have a complete deficiency of IFs in reactive astrocytes, but there was normal healing in GFAP−/− or Vim−/−mice, which have only a partial deficiency (Menet et al, 2003; Pekny et al, 1999b; Wilhelmsson et al, 2004). However, we cannot exclude the possibility that partial IF deficiency had a subtle effect that escaped detection. In another study, GFAP−/− mice and WT controls had similar infarct volumes after 2 days of permanent MCA occlusion; however, when MCA occlusion was combined with transient occlusion of the carotid artery, the GFAP−/− mice had larger infarcts than the controls (Nawashiro et al, 2000).
Vimentin is also expressed in other cell types in the CNS, particularly endothelial cells, oligodendroglia, and microglia. Endothelial cells in Vim−/−mice are devoid of IFs because the other IF protein expressed in these cells, nestin, cannot form filaments on its own (Eliasson et al, 1999; Pekny et al, 1999b). Thus, with regard to IF production in reactive astrocytes, the only difference between Vim−/− and GFAP−/−Vim−/− mice is the partial or complete absence, respectively, of IFs. Since stroke volumes were not significantly different in Vim−/−mice and WT controls, the absence of IFs in endothelial cells and other cell types does not affect infarct size. GFAP−/−Vim−/− and WT mice had comparable cerebrovascular architecture, as well as comparable mean blood pressure and heart rate before and during the focal brain ischemia. This implies that the increased infarct size after MCA transection in GFAP−/−Vim−/− mice is a consequence of the absence of IFs in astrocytes.
How might the absence of IFs in reactive astrocytes explain the increased infarct size in GFAP−/−Vim−/− mice? The lack of IFs may compromise several functions of astrocytes, namely AGJC, uptake and transport of molecules such as glutamate, glucose, and ascorbate, protection against tPA-mediated neurotoxicity, reconstruction of damaged BBB, migration into the injured area, and regulation of cell volume.
We saw a close colocalization between bundles of IFs and ETBR immunoreactivity in cultured WT astrocytes and altered ETBR immunoreactivity in GFAP−/−Vim−/− astrocytes both in vitro and in vivo. These findings suggest that ETBRs associate with IFs and are freely distributed throughout the cell in IF-free astrocytes, which also exhibit less efficient ETBR-mediated inhibition of AGJC. Astrocyte gap-junctional communication was proposed to promote secondary expansion of focal injury, since open astrocytic gap junctions can mediate the propagation of cell death signals or undesirable backflow of ATP from living to dying cells (Lin et al, 1998). Thus, less efficient ETBR-mediated inhibition of AGJC in GFAP−/−Vim−/− mice may have contributed to larger infarct volume in these mice.
Astrocytes play a major role in the transport and metabolism of a range of molecules, in particular those utilized in nutrition, cell—cell signaling, and neurotransmission. A failure to carry out these functions may constitute a major pathogenic component in stroke and other CNS pathologies (reviewed in Kraig et al, 1995). In primary cultures of reactive astrocytes from WT, GFAP−/−, Vim−/−, and GFAP−/−Vim−/− mice, we found no differences in their ability to transport glucose and ascorbate (Pekny et al, 1999a). Previously, we showed that despite comparable availability of GS (Wilhelmsson et al, 2004), glutamine levels in reactive astrocytes from both GFAP−/− and GFAP−/−Vim−/− mice were increased by 70 to 100% (Pekny et al, 1999a). Our present data show that glutamate transport is reduced in brain cortices of GFAP−/−Vim−/− mice. It is tempting to speculate that the reduced ability of the GFAP−/−Vim−/− mice to remove glutamate from the ischemic brain tissue, possibly due to deficient intracellular trafficking of GLT-1, is the consequence of intracellular glutamine accumulation. It was shown recently that GLT-1 immunoreactivity in astrocytes colocalized with bundles of IFs (Shobha et al, 2007) and GLT-1 trafficking and cluster formation within astrocyte processes were proposed to be dependent on the cytoskeleton (Zhou and Sutherland, 2004). Notably, GLT-1−/− mice show more extensive ischemia-induced damage (Mitani and Tanaka, 2003), and when GLT-1+/− mice are crossed to superoxide dismutase-1 (G93A) mice, a model of amyotrophic lateral sclerosis, they have more prominent loss of motor neurons and muscle strength than SOD-1 (G93A) mice alone (Pardo et al, 2006). These findings indicate that glutamate transporter dysfunction influences astrocyte—neuron interactions after injury.
Among 1,200 genes whose expression was compared between WT and GFAP−/−Vim−/− astrocytes, PAI-1 was the only gene that showed at least threefold downregulation in the mutants. Quantitative real-time PCR confirmed the downregulation of PAI-1 mRNA in GFAP−/−Vim−/− astrocytes. Plasminogen activator inhibitor-1 inhibits tPA, which has a neurotoxic effect in the ischemic penumbra, probably mediated through the activation of both microglia and N-methyl D-aspartate receptors (Sheehan and Tsirka, 2005). Mice deficient in tPA have smaller infarcts than WT controls in a transient focal ischemia model, and administration of tPA to both tPA-deficient and WT mice increases infarct volume (Tsirka et al, 1995). Similarly, mice over-expressing PAI-1 have smaller infarcts than WT controls after focal brain ischemia (Nagai et al, 2005). Thus, reduced levels of PAI-1 mRNA could enhance the neurotoxic effects of tPA in the ischemic penumbra of GFAP−/−Vim−/− mice.
Reactive astrocytes play a key role in the reconstruction of damaged BBB after both trauma and stroke. In a previous study with the in vitro BBB model, we demonstrated that IF-deficient astrocytes were either unable to induce BBB properties in endothelial cells or did so less efficiently (Pekny et al, 1998b). Thus, less efficient reconstruction of the BBB in the penumbra may contribute to the increased infarct volume in GFAP−/−Vim−/− mice.
In response to CNS injury, astroglial cells migrate over considerable distances to the injured region, where they join the local pool of reactive astrocytes (Johansson et al, 1999). We have shown that partial or complete deficiency of IFs in reactive astrocytes in vitro reduces their ability to migrate (Lepekhin et al, 2001). However, since the overall number of astrocytes and microglia around the infarct area did not differ in GFAP−/−Vim−/− and WT mice, reduced migration does not seem to contribute to the increased infarct volume.
The ability of astrocytes to regulate their volume by releasing osmotically active molecules, such as taurine, is considered to be a mechanism for counteracting cytotoxic brain edema in stroke (Kimelberg, 1991). When subjected to hypotonic stress, GFAP−/−Vim−/− reactive astrocytes in vitro release 25 to 46% less taurine than WT controls, indicating a reduced capacity to counteract cell swelling (Ding et al, 1998) that could have contributed to the increased infarct volumes in GFAP−/−Vim−/− mice. Consistent with this possibility, we showed that mice with IF-free nonreactive astrocytes have more prolonged changes in extracellular space diffusion parameters than WT controls during cell swelling evoked by hypotonic stress or high [K+] (Anderova et al, 2001).
Previously, we showed that attenuation of reactive gliosis in GFAP−/−Vim−/− mice increases synaptic loss at the initial stage after neurotrauma, but allows complete synaptic regeneration later on (Wilhelmsson et al, 2004). Thus, reactive gliosis seems to play a positive role soon after CNS injury, although it seems to pose an obstacle to regeneration at a later stage (reviewed in Pekny and Pekna, 2004). The results of a recent study using conditional ablation of Stat3 and Socs3 also point to a dual role of reactive astrocytes after spinal cord injury (Okada et al, 2006). Here we show a similar neuroprotective effect of reactive gliosis in brain ischemia. Genetic ablation of astrocyte IFs led to altered intracellular distribution of ETBR immunoreactivity in reactive astrocytes, both in the dentate gyrus affected by neurodegeneration induced by entorhinal cortex lesion (Wilhelmsson et al, 2004) and in ischemic tissue as shown here. Thus, less efficient ETBR-mediated inhibition of AGJC in GFAP−/−Vim−/− astrocytes might contribute to both increased synaptic loss in the acute stage after entorhinal cortex lesion and to larger infarct volume after focal brain ischemia in these mice.
In summary, our findings provide in vivo evidence that reactive astrocytes play a protective role in ischemic stroke.
Footnotes
Acknowledgements
We thank Dr Anders Hamberger, Dr John Eriksson, and Hanna-Mari Pallari for their input in this project and Dr Martin Rydmark for his advice on stereomeric evaluation of the infarct volume.