
Abstract
The intermediate filament nestin is upregulated in response to cerebral ischemia; the significance of this, however, is incompletely understood. Here, we used transgenic mice that express green fluorescent protein (GFP) under control of the nestin promotor to characterize the fate of nestin-expressing cells up to 8 weeks after 30 mins occlusion of the middle cerebral artery (MCAo) and reperfusion. The population of nestin-GFP + cells increased in the ischemic lesion rim and core within 4 days, did not become TUNEL-positive, and was detectable up to 8 weeks in the lesion scar. Nestin-GFP + cells proliferated in situ and underwent approximately one round of cell division. They were not recruited in large numbers from the subventricular zone (SVZ) as indicated by absence of colabeling with intracerebroventricularly injected dye DiI in the majority of nestin-GFP + cells. Nestin-GFP + cells expressed the chondroitin sulfate proteoglycan NG2 and nestin protein, but typically lacked mature astrocytic markers, that is, glial fibrillary acid protein (GFAP) or S100 β. Vice versa, the majority of GFAP + cells lacked nestin-expression and surrounded the ischemic lesion by 4 days. Whole-cell patch-clamp recordings in acute brain slices from controls showed that only about half of nestin-GFP + cells displayed complex membrane properties. In contrast, 4 days after the insult all nestin-GFP + cells expressed these properties. We hypothesize that the change in physiologic properties induced by the ischemic insult is directed toward a specific function of nestin-expressing cells.
Introduction
Nestin represents a novel class of intermediate filament strongly expressed in multipotent CNS precursor cells (Lendahl et al, 1990). While nestin immunoreactivity in the adult brain is generally low and mainly observed in certain proliferative regions such as the subventricular zone (SVZ) or the hippocampus (e.g., Wei et al, 2002), marked upregulation of nestin has been shown in the lesioned brain (Lin et al, 1995; Duggal et al, 1997; Li and Chopp, 1999; Bond et al, 2002). Increased nestin expression persists for many weeks after brain ischemia and nestin + cells have been reported to typically colabel with glial fibrillary acid protein (GFAP), particularly in the ischemic border zone (Li and Chopp, 1999). Nestin was therefore considered a marker for reactive astrocytes, although nestin expression in neurons and oligodendrocytes has also been claimed (Li and Chopp, 1999). Reactive astrocytes are thought to play a major role in the healing process after brain injury and it has even been proposed that ‘recovery from ischemic damage may recapitulate ontogeny’, that is, the reexpression of developmental proteins may indicate active reconditioning that could promote cell survival (Cramer and Chopp, 2000). Given the fact that GFAP + astrocytes in the SVZ as well as in the subgranular zone of the dentate gyrus have been identified as neural precursor cells, this may indicate that in response to injury, subpopulations of astrocytes possess the ability to return to a more undifferentiated developmental stage (Doetsch et al, 1999; Laywell et al, 2000; Seri et al, 2001).
Adult neurogenesis is well characterized in the olfactory bulb (Alvarez-Buylla and Garcia-Verdugo, 2002; Doetsch et al, 1999) as well as in the dentate gyrus of the hippocampus (e.g., Altman and Das, 1965; Kempermann et al, 1998). This finding has energized research into the ability of the central nervous system (CNS) for self-repair through neuronal cell replacement (Lowenstein and Parent, 1999). In fact, neurogenesis in otherwise nonneurogenic regions has recently been reported after ischemic tissue damage (e.g., Nakatomi et al, 2002; Arvidsson et al, 2002). We and others have characterized progenitor cell heterogeneity in the dentate subgranular zone taking advantage of transgenic mice that express green fluorescent protein (GFP) under a neural-specific enhancer region of the nestin promotor (Yamaguchi et al, 2000; Kronenberg et al, 2003; Filippov et al, 2003; Fukuda et al, 2003).
The present study was designed to describe in a comprehensive immunohistochemical and electrophysiologic approach the effects of mild transient ischemia on nestin-expressing cells in the striatum up to 8 weeks after occlusion of the middle cerebral artery (MCAo)/reperfusion. We show that nestin-expressing cells are biologically distinct from GFAP + reactive astrocytes and adopt complex membrane properties at early time points after mild cerebral ischemia.
Materials and methods
Animals and Drug Administration
All experiments were performed according to the national and institutional guidelines and were approved by an official committee (G0113/00 LaGeTSi, Berlin). Mice expressing GFP under nestin gene regulatory elements were generated on a C57/BL6 background as described elsewhere (Yamaguchi et al, 2000; Fukuda et al, 2003). In some experiments, 129/Sv mice were used. Animals were 10 to 12 weeks old and weighed 20 to 25 g at the beginning of the experiments. Bromodeoxyuridine (BrdU; Sigma-Aldrich, Deisenhofen, Germany) was administered at a dose of 1 mg/h kg via subcutaneously implanted osmotic minipumps (flow rate 1 µL/h; Alzet, Cupertino, CA, USA). This dose is nontoxic and labels approximately 4 times more cells after cerebral ischemia than twice daily intraperitoneal injections at 50 mg/kg (Katchanov et al, 2001). To study cell migration from the SVZ to the ischemic striatum, we injected DiI (0.5 µL of 0.2% wt/vol DiI in DMSO) into the right ventricle (0.75 mm lateral to bregma and 2.8 mm from the dural surface) using a Hamilton syringe (Jin et al, 2003).
Induction of Cerebral Ischemia
Animals were anesthetized for induction with 1.5% isoflurane and maintained in 1.0% isoflurane in 69% N2O and 30% O2 with the use of a vaporizer. Left MCAo for 30 mins was induced with a silicone-coated 8.0 nylon monofilament and was essentially performed as described (Endres et al, 1998). To insure equivalent levels of ischemia, regional cerebral blood flow was measured using laser Doppler flowmetry and flexible skull probe. Core temperature was maintained at 36.5°C ± 0.5°C during the monitoring period until 1 h after reperfusion using a heating pad. After a survival time ranging from 24 h to 8 weeks animals were killed for histologic or electrophysiologic analysis.
Immunohistochemistry and Immunofluorescence
Mice were killed with an overdose of pentobarbital and perfused transcardially with 0.9% saline followed by 4% paraformaldehyde in 0.1 mol/L phosphate buffer. Brains were stored in the fixative for 48 h and then transferred into 30% sucrose in 0.1 mol/L phosphate buffer for 24 h. Coronal sections of 40 µm thickness were cut from a dry ice-cooled block on a sliding microtome (Leica, Bensheim, Germany). Sections were stored at −20°C in cryoprotectant solution containing 25% ethylene glycol, 25% glycerin, and 0.05 mol/L phosphate buffer. All antibodies were diluted in Tris-buffered saline containing 0.1% Triton X-100 and 3% donkey serum.
For
Cell Counts
Densities of nestin-GFP + or Ki67 + cells in ischemic striatum were quantified with StereoInvestigator (Microbrightfield Europe, Magdeburg, Germany) by circling the ischemic striatum in a reference section at approximately bregma 1.32 excluding the SVZ and the alveus. All cells within the ischemic striatum were marked and counted and this cell count was then divided by the corresponding volume of tissue.
Tissue Preparation for Electrophysiology
The procedure for the preparation of striatal slices from nestin-GFP transgenic mice after MCAo was performed as described previously (Filippov et al, 2003). Briefly, mice were killed by an overdose of pentobarbital, decapitated and their brains were dissected, washed, and the hemispheres were cut into 150 µm-thick slices in frontal orientation using the Leica VT1000S vibratome (Leica, Nussloch, Germany) at 4°C external solution (vide infra). Subsequently, the brain slices were gently transferred with a pipette to a holding chamber containing the standard bath solution until used for recording. Slices were stored at room temperature (21°C to 26°C) for at least 30 mins before recording.
Patch-Clamp Recordings
For patch-clamp analysis in situ, slices were gently transferred to a perfusing chamber installed on the stage of a microscope (Axioskop, Zeiss, Oberkochen, Germany; × 5 Zeiss objective, numerical aperture 0.15). The chamber was continuously perfused with bicarbonate-buffered bath solution (ACSF) composed of (in mmol/L): 134 NaCl, 2.5 KCl, 1.3 MgCl2, 2 CaCl2, 1.25 K2HPO4, 26 NaHCO3, 10
Identification of Nestin-GFP + Cells
Using standard transmission optics (Zeiss Axioskop, Zeiss, Oberkochen, Germany; × 5 Zeiss objective, numerical aperture 0.15), the striatum was identified. Images were recorded with a CCD camera and stored on computer (Variocam, PCO Computer Optics, Kelheim, Germany). GFP + cells were identified by fluorescence optics (excitation at 488 nm using a monochromator, Polychrome IV, Till Photonics, Martinsried, Germany). The emitted light was collected at 530 ± 10 nm with a CCD camera QuantiCam (b/w VGA, Phase, Lübeck, Germany; × 60 water immersion Olympus objective, numerical aperture 0.8). After patch-clamp recording, Alexa Fluor 594 fluorescence was detected at an excitation wavelength of 589 nm and an emission at 616 ± 4 nm.
Statistics
All values are expressed as mean ± s.d. Differences between groups were evaluated by unpaired Student's t-test or by ANOVA followed by Post hoc testing as indicated. P-values of < 0.05 were considered statistically significant.
Results
Nestin-GFP + Cells are Upregulated After 30 mins MCAo/Reperfusion
Nestin-GFP mice were subjected to 30 mins left MCAo/reperfusion or sham operation and were killed 1, 2, 4, 7, 28, or 56 days later. Nestin-GFP + cells were visualized on serial coronal microtome sections (40 µm thickness) immunohistochemically using an antibody directed against GFP (Figure 1). In sham-operated animals as well as in the contralateral, intact hemisphere a considerable number of weakly fluorescent nestin-GFP + cells with a delicate morphology and thin processes were found. The overall intensity of GFP-immunoreactivity was upregulated as early as 24 h after ischemia while the absolute number of nestin-GFP + cells increased by 4 days (Table 1). This increase in nestin-GFP + cell number and of GFP-immunoreactivity after stroke was restricted to the territory of the occluded middle cerebral artery. By 4 days after MCAo/reperfusion, GFP-immunoreactivity yielded a clear demarcation between the lesioned area and adjacent tissue both in the striatum and overlying cerebral cortex (boxed areas in Figures 1A and 1B). The majority of GFP + cells were located in the ischemic core and border zone at earlier time points (i.e., 2 to 7 days after MCAo). A scar-like conglomerate composed of nestin-GFP + cells was still evident within the lesion core after 8 weeks (Figure 2).
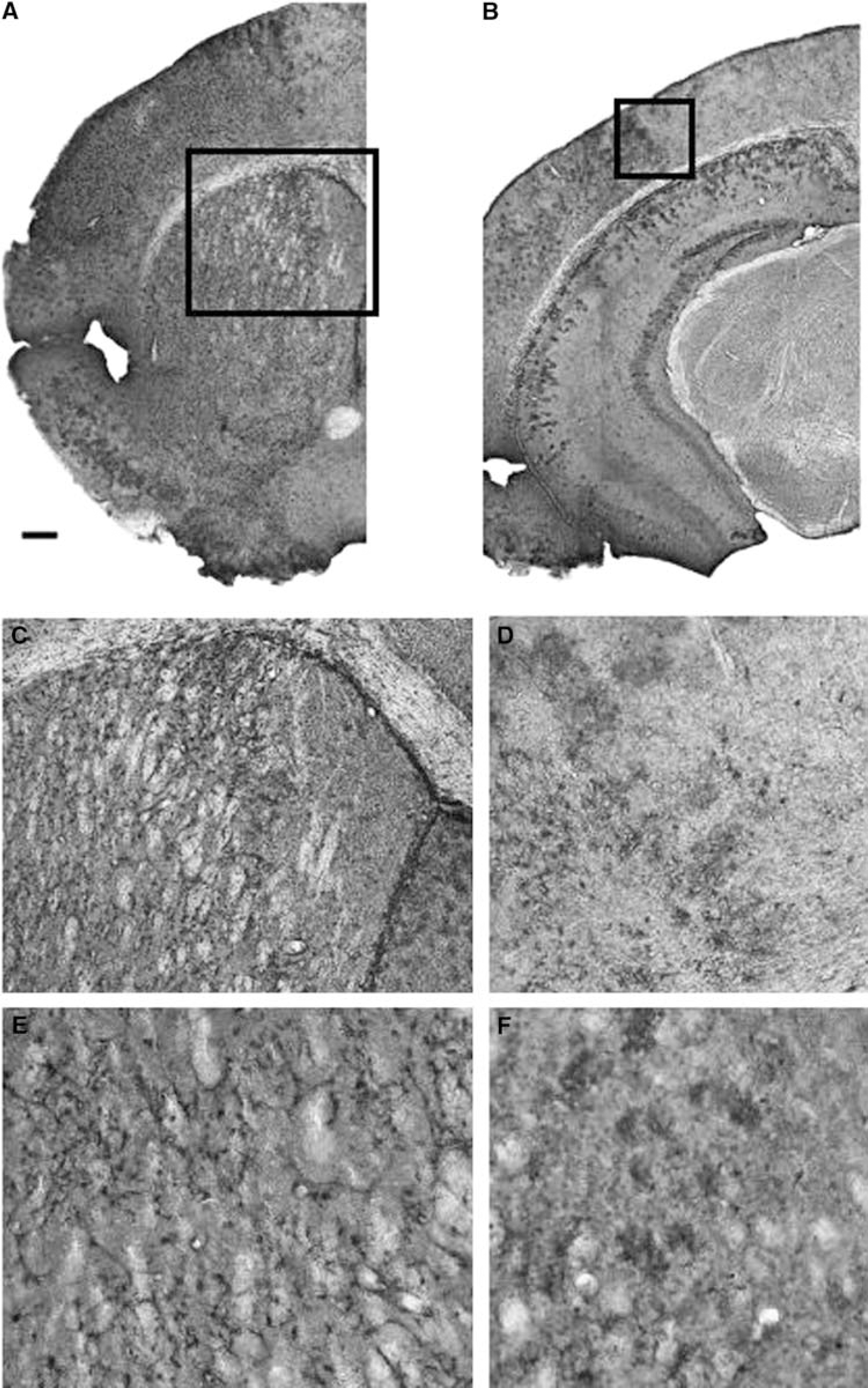
Nestin-GFP-immunoreactivity after cerebral ischemia. Nestin-GFP mice were subjected to 30 mins left MCAo and 4 days reperfusion. Nestin + cells were identified by anti-GFP-immunostaining and visualization with diaminobenzidine (DAB) on coronal brain sections (40 µm). Several pictures at × 2.5 objective have been joined together to yield composite images in (
Number of nestin-GFP + and of Ki67 + cells (×102/mm3) at different time points after 30 mins MCAo/reperfusion in the ischemic striatum
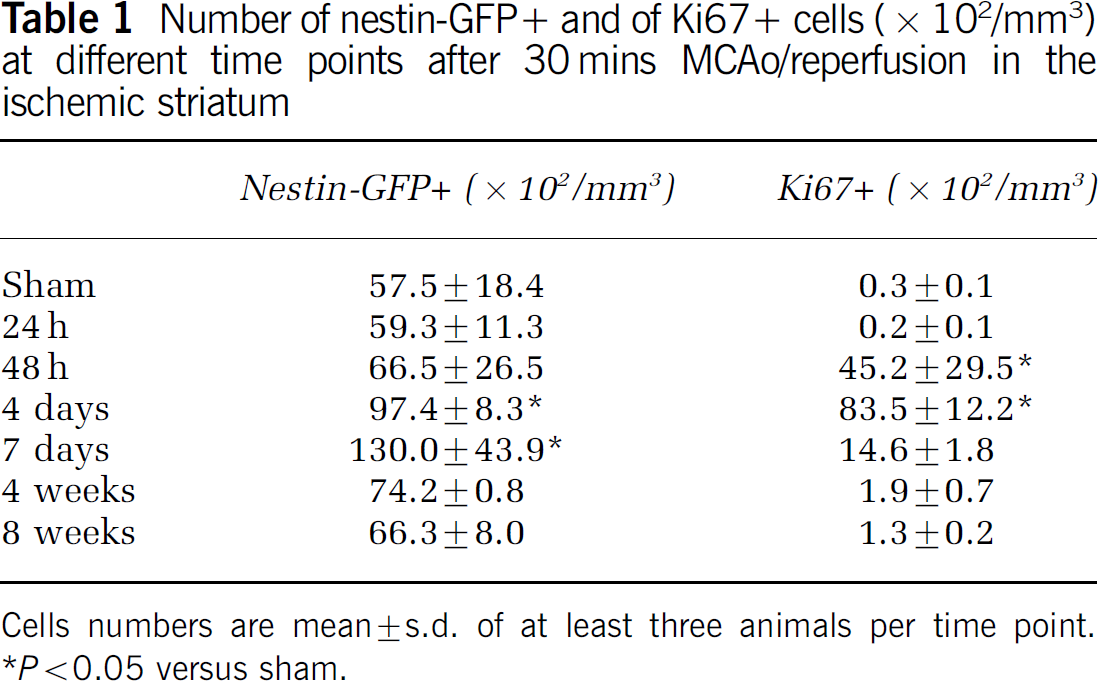
Cells numbers are mean ± s.d. of at least three animals per time point.
P < 0.05 versus sham.
Nestin-GFP + Cells Typically Lack Mature Astrocytic Markers but Express the Chondroitin Sulfate Proteoglycan NG2
To further characterize the phenotype of nestin-GFP + cells, we performed double-immunostaining with antibodies against the nestin protein, NG2, GFAP, and S100β (Table 2). Whereas in control animals and after sham operation, striatal nestin expression as detected by an antibody against the nestin protein was very weak, immunohistochemical analysis at 4 days after MCAo/reperfusion confirmed that the majority of nestin-GFP + cells were also strongly nestin-immunopositive. Vice versa, only a minority of nestin-immunoreactive cells lacked GFP (~15%); these cells were typically located toward the border (rather than the core) of the lesion and frequently coexpressed GFAP. In addition, some nestin +/GFP–cells displayed an endothelial morphology. Only 16% of nestin-GFP + cells expressed astrocytic marker GFAP and less than 2% expressed more mature astrocytic marker S100β 4 days after MCAo/reperfusion (Table 2). Nestin-GFP/GFAP double-labeled cells were typically located in the lesion perimeter. In fact, the spatial distribution of nestin-GFP + cells (i.e., primarily within the ischemic lesion) differed considerably from that of GFAP + cells (i.e., located mainly around the lesion) in the ischemic hemisphere, particularly at early time points after MCAo (Figures 2B–2E, 2G–2I). A similar pattern was observed when GFAP and nestin double-immunostaining was performed in 129/Sv mice after 30 mins MCAo/reperfusion. Only a few cells within the (inner) ischemic border zone coexpressed both markers after 72 and 96 h; GFAP +/nestin− cells were located in the outer border zone, and nestin +/GFAP− cells were located within the ischemic lesion (Figure 3A). At 4 days after MCAo/reperfusion, all nestin-GFP + cells analyzed within the lesion coexpressed the chondroitin sulfate proteoglycan NG2 (Lin and Bergles, 2002; Figure 3B). Relatively few nestin-GFP + cells displayed coexpression of both GFAP and NG2 (i.e., 15 out of 100 cells investigated) (Figure 3C). In comparison, under sham conditions the proportion of nestin-GFP + cells that were immunopositive for GFAP, S100β, or NG2 was 3%, 34%, or 58%, respectively (100 randomly selected cells from three animals for each staining). Although there was considerable overlap both in the sham condition and at earlier time points after MCAo, not all NG2 + cells expressed nestin-GFP (e.g., only 92 out of 100 cells investigated at 4 days after MCAo).
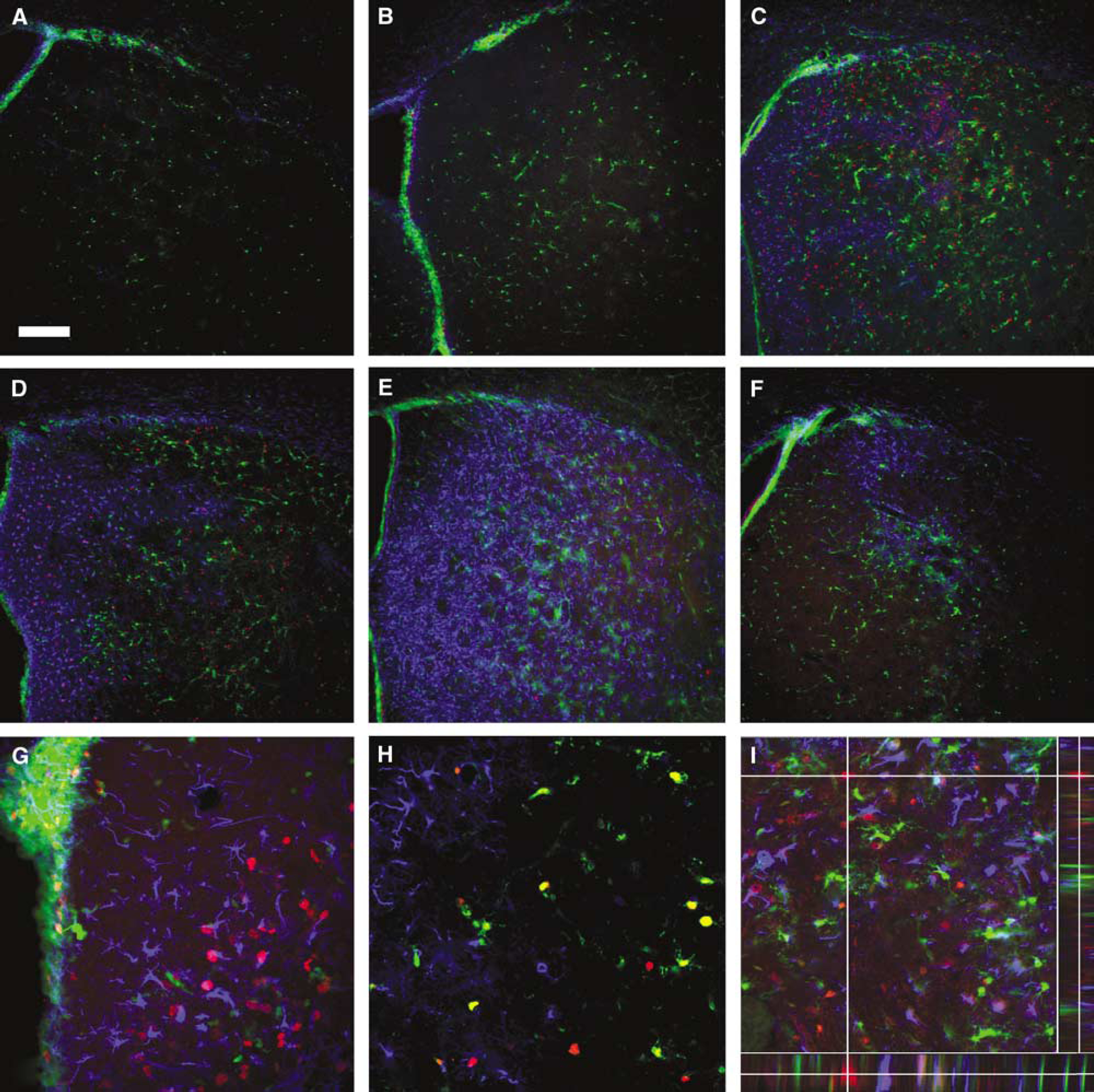
Temporal and spatial distribution of nestin-GFP +, GFAP + and Ki67 + cells after cerebral ischemia. Nestin-GFP mice were subjected to 30 mins left MCAo/reperfusion (
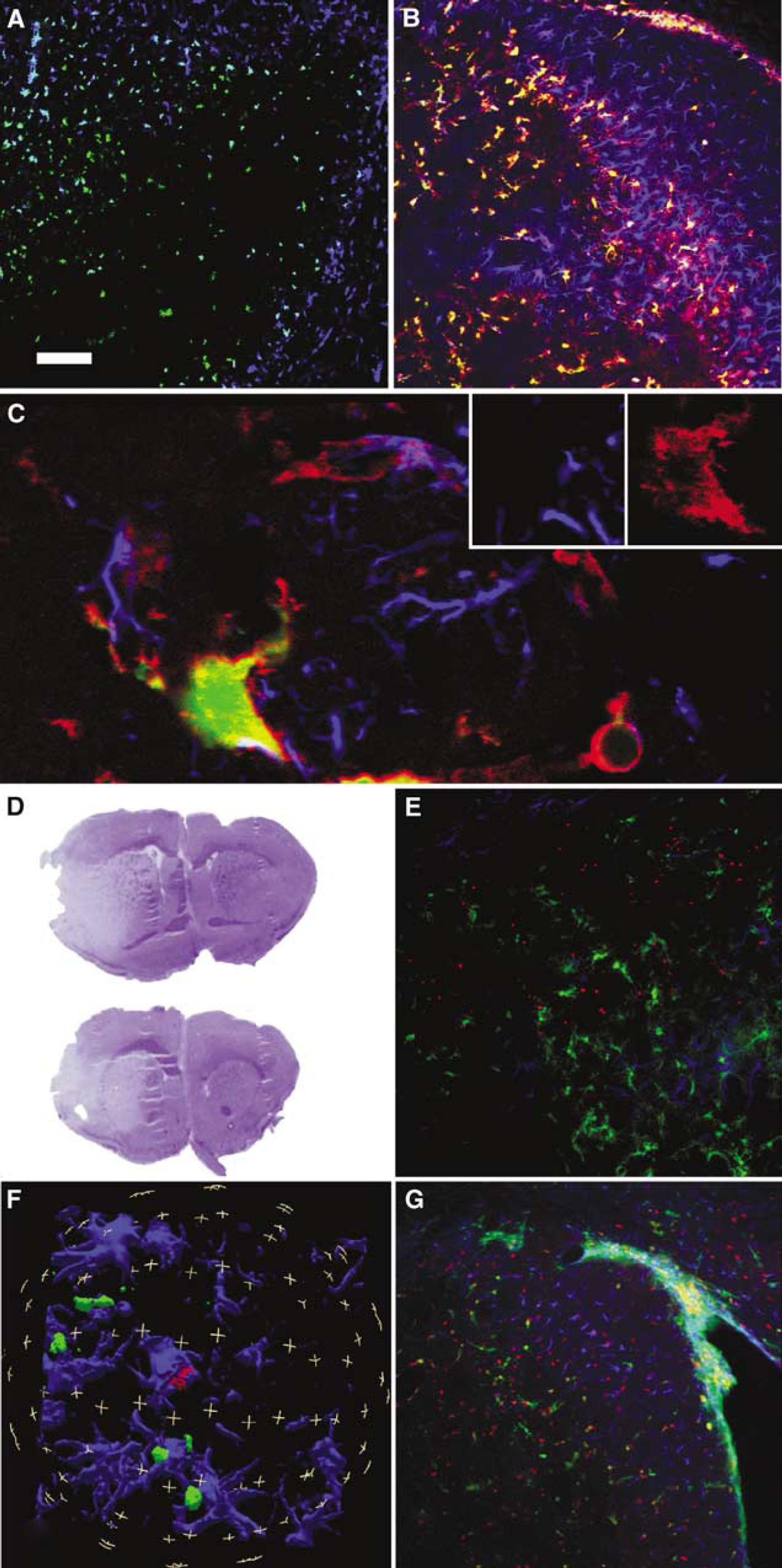
Different spatial distribution of nestin-GFP + and of GFAP + cells at early time points after stroke. (
Immunohistochemical characteristics of nestin-GFP + cells at 4 days after MCAo/reperfusion

More than 300 randomly selected nestin-GFP + cells located within the ischemic striatum from three different animals were analyzed. A total of 100 cells in the striatum of three sham-operated animals were analyzed. ND = not determined.
GFAP + and Nestin-GFP + Cells are Distinct Cellular Populations
In addition, we characterized GFAP + cells over time. Similar to nestin-GFP + cells, the number of GFAP-immunoreactive cells increased in the ischemic hemisphere after MCAo/reperfusion. At 4 days after MCAo/reperfusion, only 5% of GFAP + cells were nestin-GFP + (vide supra). The distribution of GFAP + cells in the ischemic hemisphere differed from that of nestin-GFP + cells: whereas nestin-GFP + cells were predominantly located in the lateral striatum (MCA territory; Figure 1), GFAP-immunoreactivity was most pronounced in the medial striatum close to the lateral ventricle as well as above and below the corpus callosum (ACA territory; Figure 2). As evidenced in hematoxylin-stained adjacent sections, the lateral striatum was subject to ischemic cell death while the medial striatum was spared (Figure 3D). Further phenotypic analysis of GFAP + cells within the ischemic border zone at 4 days after MCAo/reperfusion revealed that 66% of GFAP + cells were also S100β positive (165 out of 250 randomly selected cells from 3 different animals). At later time points, that is, 4 and 8 weeks after the insult, GFAP + cells were also found within the ischemic lesion core.
Nestin-GFP + Cells do not Undergo Cell Death
TUNEL + cells emerged within the ischemic lesion by 48 h, peaked at approximately 4 days and decreased thereafter. TUNEL + cells colocalized with the neuronal marker NeuN while GFAP costaining revealed no double-labeling at 72 and 96 h after 30 mins MCAo/reperfusion (see also Katchanov et al, 2001). Indeed, the majority of GFAP + cells were located outside the area of TUNEL-positive staining at early time points after MCAo (vide supra). In contrast, nestin-GFP + cells were intermingled with abundant TUNEL signal within the ischemic lesion (Figure 3E). However, careful confocal high-resolution microscopy (50 randomly selected TUNEL-positive nuclei each from 3 different animals) did not reveal any colabeling of nestin-GFP + cells and TUNEL signal.
Nestin-GFP + Cells Proliferate In Situ
Compared with sham, the number of nestin-GFP + cells almost doubled during the first week after MCAo/reperfusion (Table 1). To assess cell proliferation in the ischemic striatum, we used an antibody against Ki67, a nuclear protein expressed for most part of the cell cycle (Gerdes et al, 1984). The growth fraction of nestin-GFP + cells within the lesion (i.e., number of nestin-GFP + cells, which are also Ki67 +, divided by the total number of nestin-GFP + cells) increased significantly to > 10% at earlier time points after MCAo/reperfusion (see Table 3, the total number of Ki67 + cells is given in Table 1).
Growth fraction of nestin-GFP + cells (as assessed by Ki67 co-expression) at different time points after 30 mins MCAo/reperfusion in the ischemic striatum

More than 100 randomly selected nestin-GFP + cells in three animals per time point were analyzed. *P < 0.05 versus sham.
In addition, we cumulatively labeled all newly generated cells with BrdU via continous administration by osmotic minipump (e.g., Farah, 2004; Katchanov et al, 2001) and examined the animals 4 days after MCAo/reperfusion. The percentage of nestin-GFP + cells situated within the lesion that had incorporated BrdU at this time point was 83.5 ± 7.55 (n = 4 animals; analysis of 100 cells per animal).
We compared the proliferative activity of nestin-GFP + cells to that of GFAP-immunoreactive cells by both BrdU labeling and Ki67 expression. In the continuous BrdU labeling paradigm described above, BrdU was only found in 22.3% ± 3.2% of GFAP + cells surrounding the lesion (n = 3 animals, analysis of 100 cells per animal). Proliferating astrocytes (as defined by expression of Ki67 and GFAP) around the lesion typically lacked nestin-GFP. Because GFAP-immunoreactivity is observed especially in primary and secondary astrocytic processes whereas Ki67 gives a nuclear staining, true double-staining of an individual cell needed to be confirmed by 3D-reconstruction of GFAP-Ki67 z-stacks (Figure 3F). Indeed, less than 5% of proliferating cells within the lesion core displayed GFAP-immunoreactivity. In contrast to the dividing astrocytes surrounding the ischemic lesion, these few proliferating GFAP + cells within the lesion core typically coexpressed nestin-GFP.
To investigate whether nestin-GFP + cells in the ischemic lesion were recruited from the SVZ, we intracerebroventricularly injected the dye DiI in some animals (n = 4). At 4 days after 30 mins MCAo/reperfusion, we detected only a very small number of nestin-GFP + cells (less than 1%) that displayed DiI colabeling (Figure 4). In addition, we did not detect any obvious migration of nestin-GFP + cells from the SVZ toward the ischemic striatum within the first week after MCAo. Rather, nestin-GFP + cells were evenly distributed in the lateral and medials aspects of the lesion.
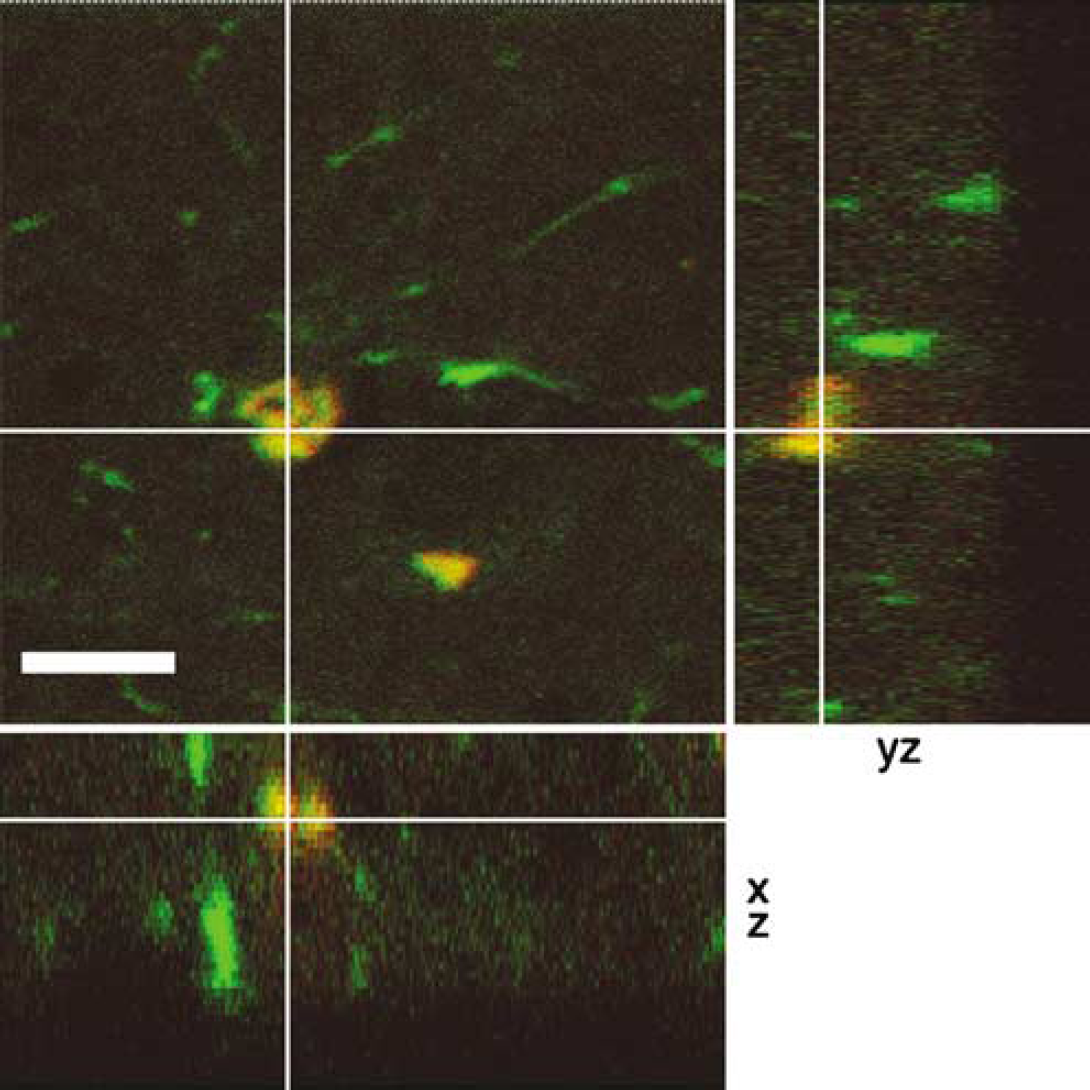
Nestin-GFP + cells were analyzed for DiI-labeling 4 days after intracerebroventricular administration of the dye and 30 mins MCAo/reperfusion. DiI-labeling was rare in nestin-GFP + cells. The image represents a three-dimensional reconstruction of a z series through nestin-GFP + cell along the yz-axis (right narrow panel) and xz-axis (bottom narrow panel), which confirms that DiI (red) and GFP (green) are indeed present in the same cell, which thus appears yellowish. Scale bar 15 µm.
Nestin-GFP + Cells Acquire the ‘Complex’ Electrophysiologic Phenotype after Stroke
Next, we analyzed the electrophysiologic properties of nestin-GFP + cells in acute brain slices at different time points after MCAo/reperfusion. After establishing the whole-cell recording configuration (Hamill et al, 1981), the membrane currents were studied by clamping the membrane at different potentials (Figure 5). We selected for cells with a stable membrane potential, which was between −50 and −85 mV (n = 111). The membrane was clamped from the holding potential of −70 mV to hyper-and depolarizing potentials ranging from −180 to + 80 mV (50 ms, 10 mV increment). Based on the membrane currents, two populations of nestin-GFP + cells were identified. (1) A ‘complex’ current pattern was characterized by outwardly rectifying currents activated by depolarization and inward rectifying, inactivation currents induced with hyperpolarization (Figure 5E). (2) The current profile of the second subpopulation was characterized by time and voltage-independent ‘passive’ currents. The resulting current/voltage curve was linear with a reversal potential at −74 mV close to the K+ equilibrium potential (Figure 5F). In control and sham-operated animals, ‘complex cells’ comprised 42% of all nestin-GFP + cells analyzed. The relative number of complex cells increased to 78.6% of nestin-GFP + cells after 2 days and to 100% at 3 and 7 days after MCAo/reperfusion. The percentage of complex cells then gradually decreased to 90% and 85% after 4 and 8 weeks, respectively (Figure 5G).
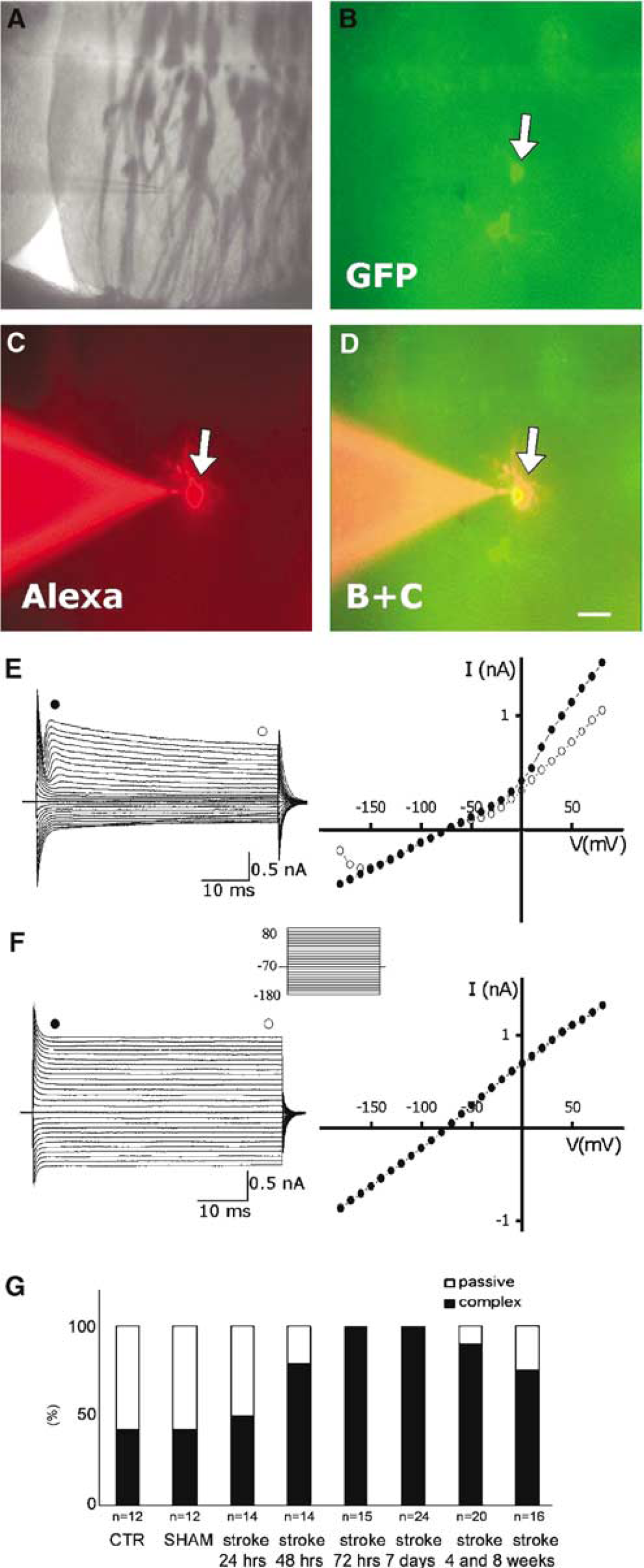
Current pattern of nestin-GFP + cells in the striatum of adult nestin promoter-GFP transgenic mice after MCAo/reperfusion. (
Discussion
This study has the following major findings: (1) Nestin-GFP + cells increase in number within the core of the lesion early after mild transient ischemia, apparently do not undergo cell death, and persist within the glial scar for at least 8 weeks. (2) The majority of nestin-GFP + cells are distinct from GFAP + reactive astrocytes that are set apart by immunohistochemical characteristics and different temporospatial distribution. (3) Nestin-GFP + cells proliferate in situ and undergo approximately one round of cell division. It is very unlikely that the increased number of nestin-GFP + cells in the ischemic striatum is because of cell migration from the SVZ. (4) Within the first week after MCAo/reperfusion, nestin-GFP + cells adopt a characteristic electrophysiologic phenotype characterized by complex membrane properties.
Nestin Immunoreactivity versus Nestin-GFP Expression
In the reporter mice used in this study, nestin-GFP + cells can be visualized in their three-dimensional structure even in acute tissue slices. In contrast, nestin staining by antibody yields an unevenly distributed cytosolic signal observed especially in the primary processes of a nestin-expressing cell. Therefore, a major additional advantage of the transgenic approach reported here is that by diffusing freely throughout the entire cell, GFP facilitates an analysis of double-labeling experiments, especially if the second antigen gives a nuclear staining like Ki67 (or in the case of TUNEL). Also, nestin antibodies crossreact with an endothelial antigen (e.g., Nacher et al, 2003, Yagita et al, 2002) whereas in the genetic construct we used GFP-expression is driven by a neural-specific enhancer region of the nestin promotor. Importantly, we found high levels of GFP and nestin-protein coexpression after ischemia.
Nestin-GFP + and GFAP + Cells are Distinct Glial Populations
Reactive astrocytosis—the proliferation and hypertrophy of astrocytes in response to brain injury—may play a major functional role in tissue recovery (Lin et al, 1995). Both nestin and GFAP are generally regarded as markers of reactive astrocytes that are upregulated after brain injury. In fact, a number of reports have shown that GFAP + reactive astrocytes coexpress nestin (e.g., Lin et al, 1995; Duggal et al, 1997; Li and Chopp, 1999). Here we show in a model of mild cerebral ischemia that—although there is indeed minor overlap—the majority of nestin-GFP + versus GFAP + cells are part of distinct glial populations. At early time points after MCAo/reperfusion (1) only approximately 15% of nestin-GFP + cells were colabeled with GFAP, and vice versa only approximately 5% of GFAP + cells were also nestin-GFP-immunoreactive. (2) Nestin-GFP + cells were located primarily within the ischemic lesion whereas GFAP + cells were located in the surrounding tissue that had remained intact. We found a high level of S100β coexpression (a marker of more mature astrocytes; e.g., Filippov et al, 2003) in GFAP + cells around the lesion while S100β immunoreactivity was virtually absent from nestin-GFP + cells in the core. Differences to earlier reports may relate to differences in the models used and to the fact that in the transgenic animal model GFP expression is driven by the nestin promotor whereas in earlier studies commercially available nestin antibodies were used. However, we did observe GFAP/nestin-GFP double-positive cells. These cells were located primarily in a small transitional zone between the lesion core and perimeter, which is in accordance with earlier studies (e.g., Duggal et al, 1997; Li and Chopp, 1999).
After a CNS stab lesion, resident NG2 + cells typically lack GFAP but frequently express vimentin and nestin and represent the main proliferative glial cell type (apart from microglia/macrophages), which subsequently differentiates further and contributes to the development of a glial scar (Alonso, 2005). The Alonso study is in general agreement with our data: all nestin-GFP + cells analyzed at earlier time-points after MCAo were also NG2 +, while NG2 + cells were scarcely GFAP + (i.e., only ~15%). Recent studies have shown both in vivo and in vitro that NG2 + cells give rise to oligodendrocytes, astrocytes, and neurons (Belachew et al, 2003; Aguirre and Gallo, 2004). Alonso showed in his model that the large majority of structures immunostained for either vimentin or nestin present in the lateral zone of the lesion (characterized by modest cell proliferation) was double-immunostained for GFAP, whereas such structures located in the border zone (i.e., a 100 µm-wide tissue portion extending along the lesion border) were GFAP—. Numerous NG2 immunoreactive cells present in the border zone were double-labeled for either vimentin or nestin (Alonso, 2005). In our study, we found that only 16% of all nestin-GFP + cells were also GFAP +, and these cells were mainly localized around the lesion.
Nestin-GFP + Cells Proliferate In Situ
Colabeling of nestin-GFP with proliferation marker Ki67 revealed a relatively low proliferation rate of roughly 10% at early time points after ischemia (Table 3). By cumulatively labeling newly generated cells with continuous administration of BrdU, we determined that by 4 days after the insult the vast majority of nestin-GFP + cells had become BrdU +. Considering the fact that by 4 days after MCAo/reperfusion the number of nestin-GFP + cells had almost doubled and that both the mother and the daughter cell become BrdU + after cell division, we infer that each nestin-GFP + cell must have undergone approximately one round of cell division after the ischemic insult (number of nestin-GFP + cells peaks within the first week after ischemia). We furthermore infer from our data that the rise in nestin-GFP + cells after the insult is due primarily to the in situ proliferation of resident nestin-GFP + cells. This fits well with the fact that DiI labeling from dye uptake at the ventricular surface was rare in nestin-GFP + cells at 4 days after MCAo/reperfusion. In contrast, GFAP + reactive astrocytes showed considerably less BrdU-incorporation than nestin-GFP + cells. In our view, this indicates hat many astrocytic cells surrounding the lesion merely upregulate GFAP without a preceding cell division.
Nestin-GFP + Population Aquires a Uniform Physiological Phenotype After Lesion
Based on their expression of membrane currents, nestin-GFP + cells can be classified as displaying either ‘complex’ or ‘passive’ properties and in control animals approximately half of the cells belonged to either population. We were unable to predict the electrophysiologic properties of a cell from either its morphology or its fluorescence level. The complex current pattern observed in nestin-GFP + cells in the striatum is also described for glial progenitor cells (Sontheimer et al, 1989), a newly described subpopulation of astrocytes in the CA1 region of the hippocampus (Matthias et al, 2003) and a subpopulation of precursor cells in the dentate gyrus (Filippov et al, 2003). The passive current profile is characteristic for the majority of astrocytes (Steinhauser et al, 1992; D'Ambrosio et al, 1998). In response to the ischemic insult, the population of nestin-GFP + cells shifts toward the complex physiologic phenotype. In fact, by 4 days after the insult the entire population exhibits a complex physiologic phenotype. The absence of TUNEL-reaction in nestin-GFP + cells and the fact that 24 h after MCAo/reperfusion, nestin-GFP + cells with passive membrane properties were still detectable suggest that passive nestin-GFP + cells had not undergone apoptosis or selective necrosis (so that complex cells would have selectively survived over time). We infer from our data that in response to the ischemic injury nestin-GFP + cells with passive membrane properties transform into complex cells. In addition, in preliminary experiments we have shown that complex nestin-GFP + cells express AMPA/kainate receptors but lack glutamate transporter currents (data not shown) similar to the hippocampal ‘complex astrocytes’ (Matthias et al, 2003). It has been hypothesized that complex astrocytes might represent an intermediate ‘astron’ cell type that retains some glial properties but has already switched on neuronal genes (Matthias et al, 2003). Importantly, a subset of astrocytes with complex membrane properties has been reported to express NG2 (vide supra; Matthias et al, 2003).
Taken together, nestin-GFP + cells appear as a highly versatile cellular population largely distinct from GFAP + reactive astrocytes. It is tempting to speculate that the electrophysiologic changes after ischemia might be directed toward a specific network function of nestin-GFP + cells.