
Abstract
The effects of statins on gene expression of cerebral endothelial cells (ECs) in vivo have not been investigated after stroke. We developed a rapid double immunofluorescent staining protocol with antibodies against von Willebrand factor (a marker for endothelium) and glial fibrillary acidic protein (a marker for astrocytes) for laser capture microdissection to isolate single ECs in brain tissue of the rat. Using this protocol in combination with real-time PCR, we found that stroke significantly increased mRNA levels of protease-activated receptor 1 (PAR-1) and tissue factor (TF) in ECs isolated from ischemic cerebral microvessels compared with nonischemic vessels. Treatment of embolic stroke with recombinant human tissue plasminogen activator (rht-PA) 4 h after stroke further elevated PAR-1 mRNA levels nearly 1000-fold in the core and 500-fold in the boundary above the nonstroke group 30 h after stroke, while TF mRNA levels were elevated approximately 10 fold above the nonstroke group. Furthermore, stroke significantly increased matrix metalloproteinase (MMP) 2 and 9 mRNA levels in the ischemic core and boundary regions 6 and 30 h after stroke. Treatment with rht-PA-upregulated MMP2 expression in the ischemic boundary and core. Atorvastatin completely blocked rht-PA upregulation of the above genes, when atorvastatin in combination with rht-PA was administered 4 h after stroke. Monotherapy of atorvastatin 4 h after stroke did not significantly reduce expression of genes examined in the present study. These data provide evidence that atorvastatin reduces exogenous tPA-aggravated cerebral endothelial genes that mediate thrombosis and blood-brain barrier permeability, which could contribute to the beneficial effects of statins on thrombolytic treatment of acute stroke.
Keywords
Introduction
Treatment of stroke with the 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase inhibitors (statins) reduces infarct volume independent of the effects of statin on lowering LDL cholesterol (Bedi and Flaker, 2002; Kawashima et al, 2003; Chen et al, 2003; Miida et al, 2004). Stroke patients in a clinical trial of Markers of Inflammation after Simvastatin in Ischemic Cortical Stroke (MISTICS) showed a trend for a favorable neurological outcome at day 90, when simvastatin was administered to stroke patients between 3 and 12 h from the symptom onset (Montaner et al, 2004). In vitro studies show that statins modulate the blood coagulation cascade leading to reduced thrombogenicity, which could contribute to the beneficial effects of statins on stroke (Krysiak et al, 2003, Undas et al, 2005). Upregulation of tissue factor and thrombin, which are key factors for thrombosis, in cerebral endothelial cells (ECs) has been shown after stroke (Krysiak et al, 2003). Thrombin exerts its effects via a family of G protein-coupled protease-activated receptors (PAR) PAR-1 to PAR-4 (Hirano and Kanaide, 2003; Xi et al, 2003). Statins block thrombin-induced tissue factor activity in cultured human aortic ECs (Eto et al, 2002). However, it is unknown whether treatment with statins affects the coagulation gene expression profile of cerebral ECs in vivo.
Endothelial cells of cerebral blood vessels display characteristics that differentiate them from the endothelia of other organs (Cleaver and Melton, 2003). The cerebral ECs and astrocytes are two very important components of neurovascular units that form the blood-brain barrier (BBB) (Abbott, 2002; Ballabh et al, 2004). Although anatomically these two types of cells are in close contact, the function of astrocytes is quite different from cerebral ECs in response to stroke (Schipke and Kettenmann, 2004). Laser capture microdissection (LCM) allows the isolation of individual cells from heterogeneous cell types within the tissue, which provides a homogeneous cell population for gene expression analysis (Banks et al, 1999; Luo et al, 1999; Suarez-Quian et al, 1999; DiFrancesco et al, 2000; Jacquet et al, 2005). In the present study, we developed a double immunofluorescence protocol to visualize cerebral ECs and astrocytes, which maximizes the cellular specificity during harvest of single ECs by LCM. We then used this protocol in combination with real-time RT-PCR to examine the effects of atorvastatin on expression of genes mediating thrombosis and BBB leakage in cerebral ECs after embolic stroke in the rat.
Materials and methods
Animal Model of Stroke and Tissue Preparation
All experimental procedures were approved by the Institutional Animal Care and Use Committee of Henry Ford Hospital. Male Wistar rats weighing 320 to 380 g were employed. The middle cerebral artery (MCA) was occluded by placement of an embolus at the origin of the MCA, as described previously (Zhang et al, 1997). Rats were killed 6 and 30 h after ischemia and brain coronal sections (8 μm thick) were cut on a cryostat set at −20°C and kept at −80°C until processing. Hematoxylin and eosin (H&E) staining was performed on frozen sections from each rat to detect the ischemic lesion.
Experimental Protocol
Rats were randomly assigned to the following groups: (1) the combination group: atorvastatin (Pfizer) was administered subcutaneously at a dose of 20 mg/kg 4 h after MCA occlusion (MCAo) and was followed by a second dose of 20 mg/kg 24 h after the first dose, and recombinant human tissue plasminogen activator (rht-PA) (20 mg/kg) was intravenously administered 4 h after MCAo. Animals were killed 6 (n = 4) and 30 h (n = 6) after MCAo; (2) the rht-PA group: rht-PA was administered (intravenously) 4 h after MCAo and rats were killed 6 (n = 4) and 30 h (n = 5) after MCAo; (3) the atorvastatin group: atorvastatin was administered subcutaneously at a dose of 20 mg/kg 4 h after MCAo and was followed by a second dose of 20 mg/kg 24 h after the first dose. These rats were killed 6 (n = 4) and 30 h (n = 5) after MCAo; (4) the MCAo group: rats were killed 6 (n = 4) and 30 h (n = 4) after MCAo; (5) the control group: nonstroke rats (n = 4).
Immunofluorescent Staining
For double immunofluorescent staining, brain coronal sections localized to the territory supplied by the MCA were air-dried for 30 secs and fixed in freshly prepared 4°C cold acetone for 2 mins. The coronal sections were incubated with the first primary antibody against glial fibrillary acidic protein (GFAP) (an astrocyte marker, DAKO, 1:50 dilution) for 5 mins and then incubated with CY3-conjugated F(ab′)2 anti-rabbit IgG mouse-adsorbed secondary antibody (DAKO, 1:50 dilution) for 5 mins. After that, the second primary antibody against von Willebrand factor (vWF, an endothelial marker, DAKO, 1:50 dilution) was incubated for 5 mins and followed by FITC-conjugated F(ab′)2 goat anti-rabbit IgG secondary antibody (DAKO, 1:50 dilution). The slides were subsequently dehydrated in graded ethanol solutions (75%, 95%, 100%, each once for 30secs) and cleared in xylene for 5 mins. For single immunofluorescent staining, the second primary antibody was omitted. All procedures were performed at room temperature and completed within 30 mins.
Laser Capture Microdissection
After air-drying for at least 10 mins, vWF or GFAP immunofluorescent reactive cells were captured onto a thermoplastic film mounted on optically transparent LCM caps using the PixCell II LCM System (Arcturus Engineering). The following parameters were used during LCM: 7.5-μm laser spot size, 60-mW power, 750-μs duration. The transfer film was examined under the microscope to ensure cell lysis. Caps with cells were immediately placed into Eppendorf tubes containing 350 μL of lysis buffer and stored in −80°C before RNA isolation. Approximately 500 cells were isolated in the ischemic core and boundary regions from each rat (Figure 1).
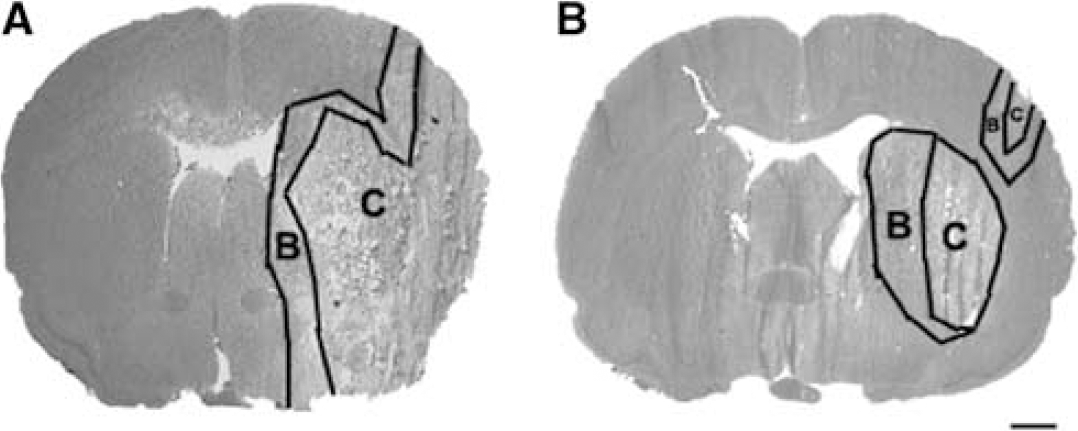
Hematoxylin and eosin-stained coronal sections illustrate the ischemic boundary and core regions from representative rats in the MCAo only (
RNA Isolation and Real-Time RT-PCR
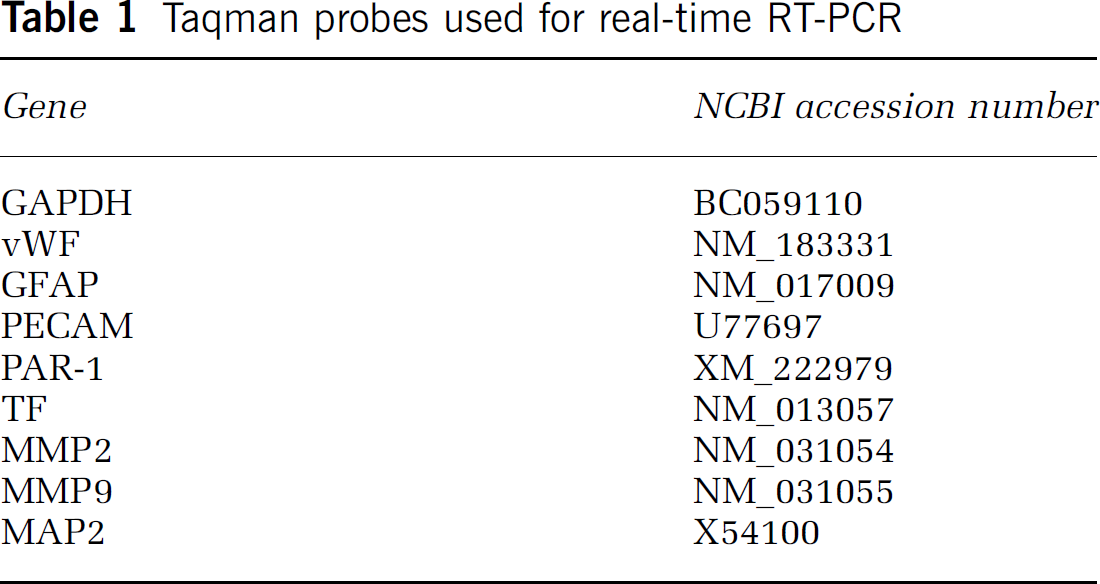
Total RNA from captured cells was isolated using RNeasy Micro Kit (Qiagen Incorporation) and was followed by reverse transcription. Total RNA from brain tissue was isolated using RNeasy Lipid Tissue Mini Kit (Qiagen Incorporation). Quantitative RT-PCR was performed on an ABI 7000 PCR instrument (Applied Biosystems) using three-stage program parameters provided by the manufacturer, as follows: 2 mins at 50°C to require optimal AmpErase uracil-N-glycosylase activity, 10 mins at 95°C to activate AmpliTaq Gold DNA polymerase, and then each cycle 15 secs at 95°C, 1 min at 60°C for 50 cycles; 2 mins at 50°C, 10 mins at 95°C, and then 40 cycles of 15 secs at 95°C and 1 min at 60°C. Table 1 lists Taqman probes (Applied Biosystems) specific for the genes examined in the present study. Each sample was tested in triplicate and data obtained from three independent experiments were expressed as a subtraction of the quantity of specific transcripts to the quantity of the control gene β-actin in mean arbitrary units. Threshold cycle (CT) values were quantified by the 2−ΔΔCt method (Livak and Schmittgen, 2001).
Taqman probes used for real-time RT-PCR
Statistical Analysis
Comparisons of mRNA levels of matrix metalloproteinase (MMP) 2, 9, TF, and PAR-1 were made between the nonstroke and MCAo rats using one-way analysis of variance (ANOVA). Analysis of variance also was used to test the effects of combination treatments rht-PA and atorvastatin on the mRNA levels of these genes. Analysis began with testing for rht-PA by atorvastatin interaction, followed by a subgroup analysis. If a significant treatment interaction between rht-PA and atorvastatin at the critical level of 0.05 was detected, all possible pairwise comparisons were made. Student's t-test was used to compare the RT-PCR measurement of mRNA levels in double and single stainings. The data are presented as mean ± standard deviation (s.d.). Differences were considered statistically significant if the P-value is < 0.05.
Results
Double Immunofluorescent Staining Enhances Cellular Specificity
Although a rapid single immunostaining protocol has been shown, a quick double-immunofluorescent staining protocol for LCM, to our knowledge, has not been developed. Figure 2 shows astrocytes (green) and microvessels (red) on double-immunofluorescent stained coronal sections. As expected, many cerebral microvessels were surrounded by astrocyte processes (Figures 2A to 2C, arrows). However, some vessels with relatively few astrocyte processes (Figures 2A to 2C, arrowhead) and individual astrocytes without connection with vessels were easily identified (Figures 2B and 2C, asterisks). In all, 29% of the microvessels (29 ± 0.03, n = 12 sections from 3 rats) exhibited connection with GFAP-positive processes. Thus, we can capture a relatively pure population of astrocytes and ECs from double-immunofluorescent stained coronal sections, in which astrocytes and microvessels were readily recognized, compared with single immunofluorescent stained ones (Figures 2D to 2I).
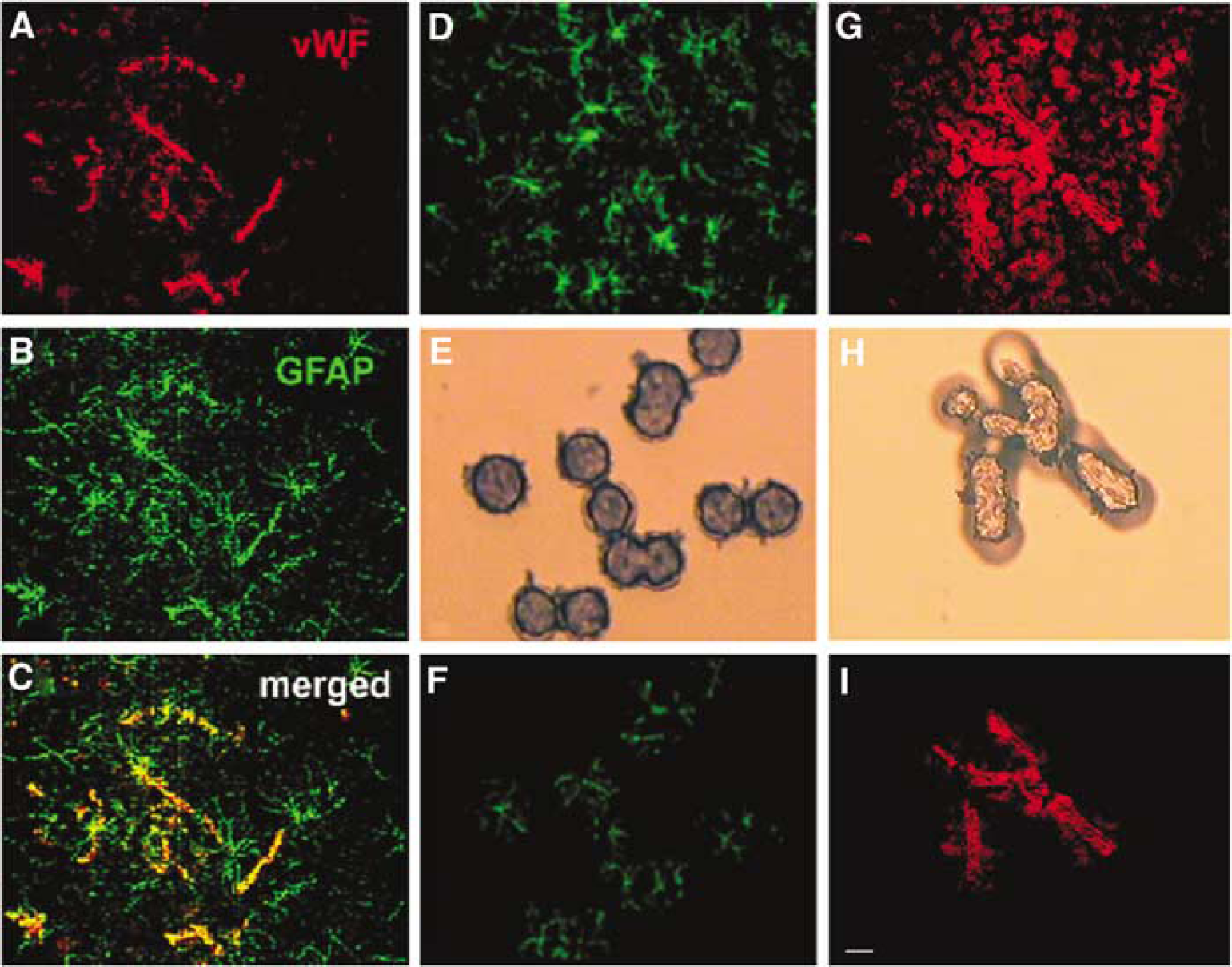
Microscopic images of single- and double-fluorescent staining with antibodies against GFAP and vWF. Panels A to C are double immunofluorescent staining images and show vWF (red)-positive vessels and GFAP (green)-positive astrocytes. Some vWF-positive vessels were contacted by GFAP-positive processes (arrows in
To examine efficiencies of amplification, standard curves with diluted samples were measured (Figure 3). The PCR efficiency of vWF from LCM and whole brain tissue was 0.88 ± 0.24 and 0.93 ± 0.03, respectively, while GAPDH from LCM and whole brain tissue was 1.18 ± 0.01 and 1.14 ± 0.13, respectively, which were not statistically different between LCM and brain tissue samples (P > 0.05).
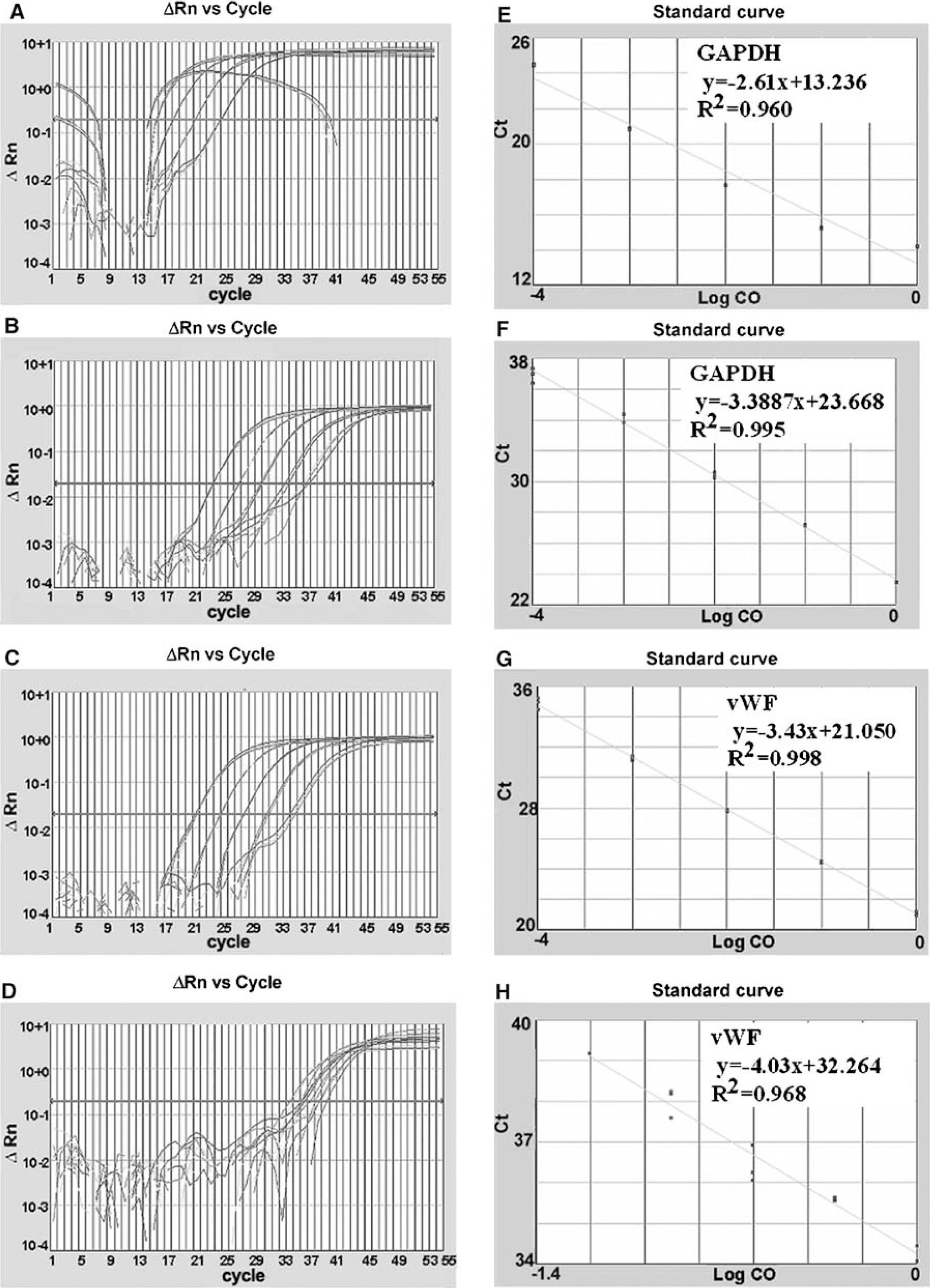
Efficiencies of RNA amplification from brain tissue and single ECs. Panels A to H show amplification plots (
Using TaqMan Probes for astrocytes, ECs, and neurons, cellular specificity was examined. Real-time RT-PCR analysis revealed that, in addition to the presence of vWF mRNA, samples captured from single vWF staining contained GFAP mRNA, while samples from vWF-positive cells in the tissue double-stained with vWF and GFAP exhibited a significant reduction in GFAP mRNA and increases in vWF mRNA compared with single vWF staining (P < 0.05) (Figures 4A and 4B). Samples from single GFAP staining showed the presence of mRNA of PECAMl (Figure 4C), a specific marker for ECs in the brain (Ulger et al, 2002). In contrast, samples from GFAP-positive cells in double-stained tissue did not exhibit mRNA of PECAMl (Figure 4C). MAP2 mRNA, a neuronal marker, was not detected in vWF-positive cells captured from single- or double-staining samples. These data indicate that double-immunofluorescent staining increases cellular specificity.
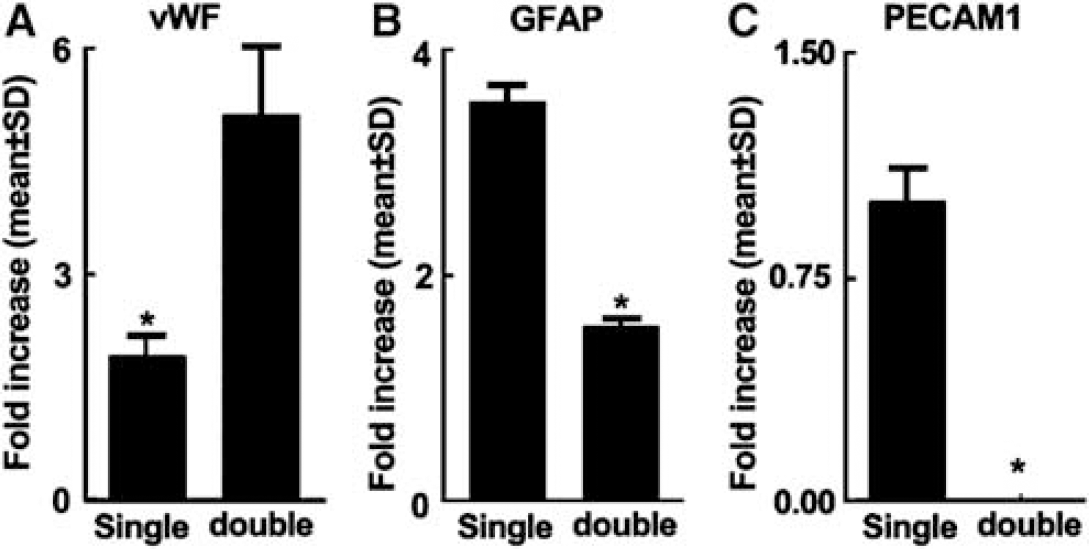
Double immunostaining increases endothelial and astrocytic specificities. Real-time RT-PCR data show vWF (
Atorvastatin Downregulates Recombinant Human Tissue Plasminogen Activator Aggravated Genes Mediating Thrombosis and Blood-Brain Barrier Leakage in Cerebral ECs
Thrombin and TF are key factors for induction of cerebral vascular thrombosis after stroke (Krysiak et al, 2003). Protease-activated receptor 1 is the prototype thrombin receptor and is activated when thrombin cleaves its NH2-terminal exodomain to unmask a new receptor NH2 terminus (Rohatgi et al, 2004). To examine the effects of atorvastatin on expression of PAR-1 and TF, vWF-positive cells were captured from brain tissues double immunofluorescently stained with antibodies against vWF and GFAP. Stroke significantly (P < 0.05) increased PAR-1 mRNA levels in the ischemic core and boundary regions 6 (Figure 5A) and 30 h (Figure 5B) after MCAo compared with the levels in nonstroke animals. Treatment with atorvastatin 4 h after MCAo did not significantly reduce PAR-1 expression (Figures 5A and 5B). Treatment with rht-PA 4 h after stroke substantially elevated PAR-1 mRNA levels in the core and boundary regions compared with the MCAo group 30 h after stroke (Figure 5B), although 6 h after MCAo, treatment with rht-PA did not significantly increase PAR-1 mRNA levels compared with the MCAo group (Figure 5A). However, treatment with atorvastatin in combination with rht-PA 4 h after stroke completely blocked rht-PA-induced upregulation of PAR-1 30 h after MCAo (Figure 5B, Table 2). In addition, stroke significantly increased TF mRNA levels in the ischemic boundary region 6 and 30 h after MCAo, and rht-PA further elevated TF mRNA levels in the ischemic core and boundary regions 30 h after stroke (Figure 5D) compared with nonstroke animals. Treatment of stroke with atorvastatin in combination with rht-PA, but not monotherapy of atorvastatin, significantly (P < 0.05) reduced rht-PA-induced TF expression in ECs of the cerebral microvessels (Figure 5D, Table 2). Matrix metalloproteinase 2 and 9 mediate BBB leakage after stroke (Asahi et al, 2001; Rosenberg, 2002). Stroke significantly (P < 0.05) increased MMP2 (Figure 6A), but not MMP9 (Figure 6C), mRNA levels in the ischemic core and boundary 6 h after MCAo. At 30 h after MCAo, significant increases of MMP2 (Figure 6B) and MMP9 (Figure 6D) expression were detected in the ischemic core and boundary regions. Atorvastatin alone significantly reduced MMP9 expression in the ischemic boundary region 30 h after MCAo (Figure 6D), but did not significantly reduce MMP2 (Figures 6A and 6B). Monotherapy of rht-PA significantly increased MMP2, but not MMP9, mRNA levels in the ischemic core and boundary regions 6 and 30 h after ischemia (Figures 6A to 6D). However, the combination treatment of atorvastatin with rht-PA significantly blocked rht-PA-upregulated MMP2 6 (Figure 6A) and 30 h (Figure 6B, Table 2) after stroke. The combination treatment also significantly decreased MMP9 expression 30 h after stroke compared MMP9 levels in the stroke group (Figure 6D, Table 2). These data indicate that the temporal and spatial profiles for expression of MMP2 and MMP9 in cerebral ECs differ in response to stroke, rht-PA and atorvastatin.
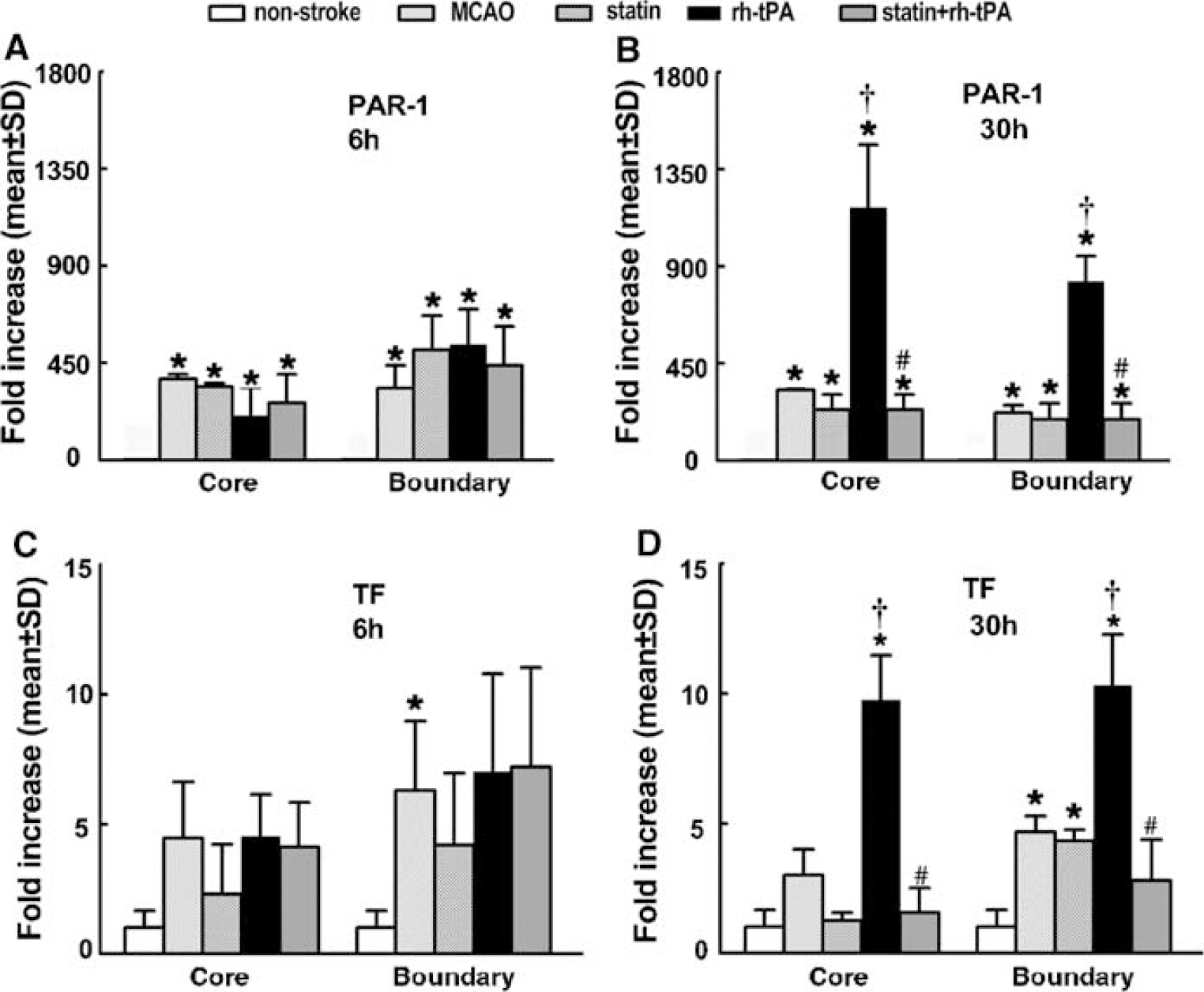
Atorvastatin downregulates rht-PA aggravated PAR-1 and TF RT-PCR analysis of cerebral ECs revealed that PAR-1 (
ANOVA results for MMP2, MMP9, TF, PAR-1 30 h after ischemia
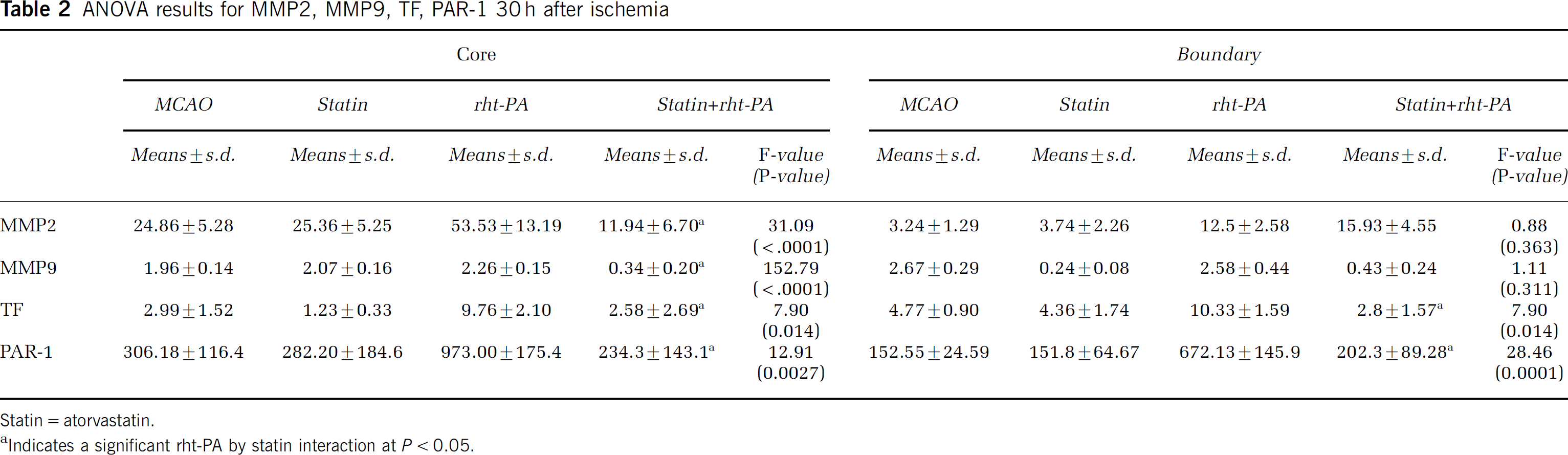
Statin = atorvastatin.
Indicates a significant rht-PA by statin interaction at P < 0.05.
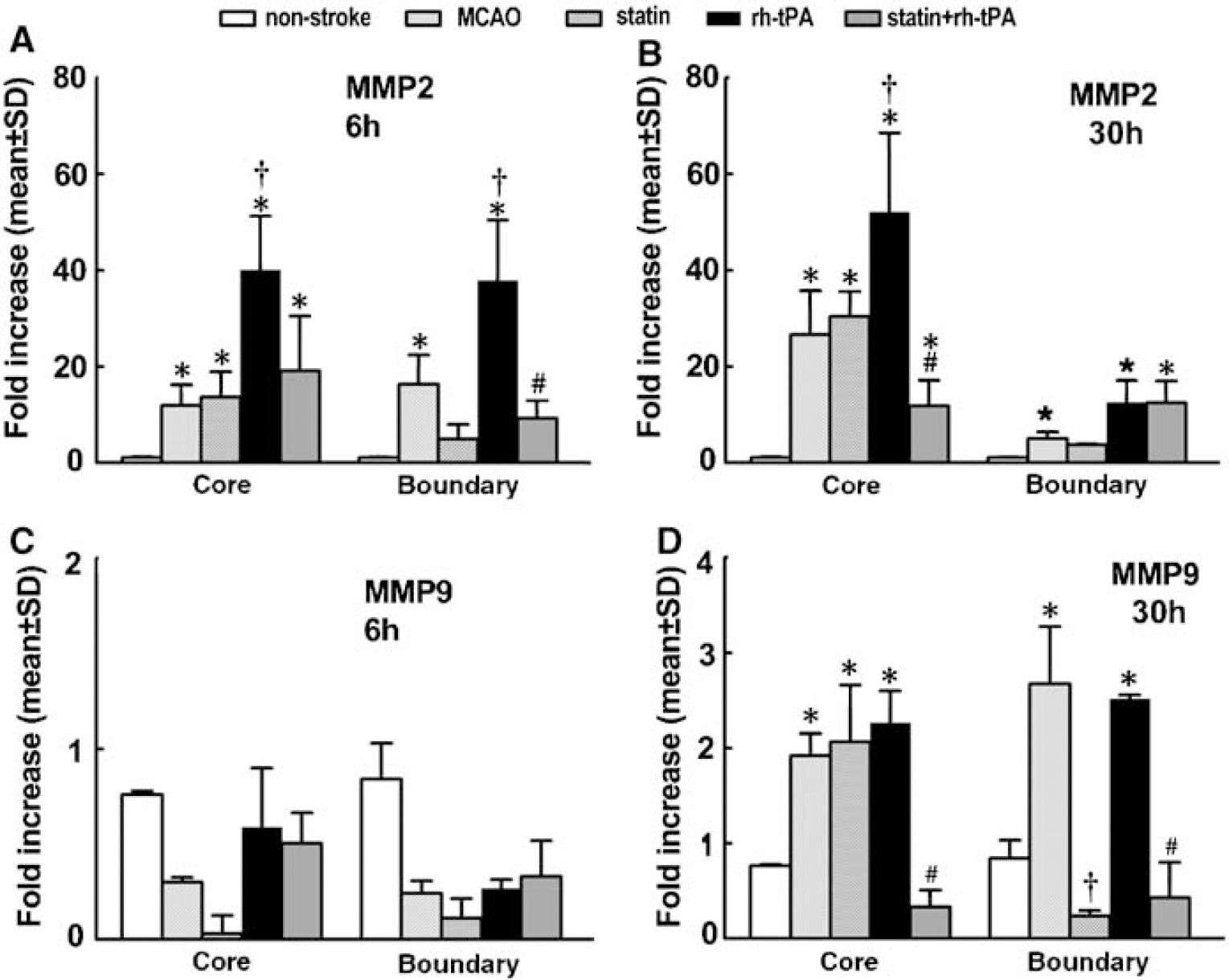
Atorvastatin downregulates rht-PA-aggravated MMP2 and MMP9. RT-PCR analysis of cerebral ECs revealed that MMP2 (
Discussion
The present study shows that cerebral ECs captured by LCM exhibited less GFAP mRNA in coronal sections with double-immunofluorescent staining for vWF and GFAP than ECs in single-immunofluorescent staining for vWF, suggesting that double-immunofluorescent staining increases the cellular specificity for LCM to capture an individual cell type from a heterogeneous cell population. In addition, we show that atorvastatin downregulates exogenous t-PA aggravated genes mediating thrombosis and BBB leakage in cerebral ECs, which could contribute to the beneficial effects of atorvastatin on thrombolysis for treatment of acute stroke.
The effects of statins on gene expression in single cerebral ECs from animals have not been investigated, although many studies have been performed in cultured ECs (Weis et al, 2002; Kalinowski et al, 2002; Miura et al, 2004). The present study shows that atorvastatin significantly blocked exogenous tPA-upregulated PAR-1 and TF in cerebral ECs, which are key modulators of thrombus formation. Plasmin, which is generated from the cleavage of plasminogen by rht-PA, activates PAR-1 (Majumdar et al, 2004). Indeed, our data show that treatment with rht-PA elevated PAR-1 levels 500- and 1000-fold in the ischemic boundary and core regions 30 h after stroke, respectively, above the normal group, while stroke alone increased PAR-1 levels up to 200-fold compared with levels in the nonstroke rats. Protease-activated receptor 1 elevated by rht-PA was coincident with increases in TF levels, suggesting that thrombin stimulates the induction of TF. Importantly, exogenous tPA-upregulated PAR-1 and TF in cerebral ECs was completely inhibited by atorvastatin, when atorvastatin was administered in combination with rht-PA 4 h after stroke. We previously showed that treatment with atorvastatin in combination with rht-PA 4 h after stroke significantly reduced infarct volume and increased cerebral microvessel patency, whereas the monotherapy of atorvastatin or rht-PA 4 h did not have neuroprotective effects (Zhang et al, 2004). The present data indicate that downregulation of PAR-1 and TF genes by atorvastatin could contribute to the effects of atorvastatin on enhancement of thrombolysis with rht-PA. Consistent with our findings, administration of rosuvastatin in combination with rht-PA significantly reverses the detrimental effects of rht-PA and reduces infarct volume (Kilic et al, 2005). The recent Interaction of Atorvastatin and Clopidogrel Study shows that administration of atorvastatin significantly reduced platelet expression of PAR-1 in patients undergoing coronary stenting (Serebruany et al, 2004). Our data provide direct evidence that, in addition to its effects on platelets in circulation, atorvastatin regulates procoagulation genes in cerebral ECs.
Matrix metalloproteinases mediate BBB permeability (Fujimura et al, 1999; Aoki et al, 2002). Thrombolysis with tPA exacerbates the disruption of the BBB after stroke (Kaur et al, 2004). Our data show that the temporal and spatial profiles of MMP2 and MMP9 expression in cerebral ECs differed in response to stroke and rht-PA, with high levels of MMP2 and MMP9 in the ischemic core and boundary regions, respectively. However, atorvastatin significantly downregulated MMP2 and MMP9 expression aggravated by exogenous tPA within both regions. These data are consistent with previous findings that the tPA mediates MMP expression in cerebral ECs via the low-density lipoprotein receptor-related protein and that treatment of diabetic retinopathy in rats with high-dose simvastatin significantly reduced vascular leakage (Miyahara et al, 2004, Wang et al, 2003). Although previous studies have shown that MMP2 and MMP9 mediate BBB leakage after stroke and tPA upregulates these gene expressions (Fujimura et al, 1999; Aoki et al, 2002), the present study shows that atorvastatin downregulates tPA-upregulated cerebral endothelial genes that mediate BBB leakage, which provides a molecular basis for the underlying beneficial effects of statins on thrombolysis for treatment of acute stroke (Zhang et al, 2004).
In contrast to the effects of atorvastatin in combination with rht-PA on reduction of gene expression, monotherapy of atorvastatin 4 h after stroke did not significantly reduce stroke-upregulated genes examined in the present study, except MMP9, which is consistent with our previous findings that administration of atorvastatin 4 h after stroke did not reduce infarct volume (Zhang et al, 2004). Reduction in cerebral vascular patency in the occluded MCA territory could result in a decreased atorvastatin access to cerebral microvessels, which may contribute to failure of monotherapy of atorvastatin to regulate endothelial gene expression in the ischemic region. Consistent with this notion, the effect of monotherapy of atorvastatin on reduction of MMP9 was only detected at the ischemic boundary.
Laser capture microdissection in combination with real-time RT-PCR has been applied for the quantification of mRNA levels with high sensitivity and reliable specificity in different tissues (Luo et al, 1999; Suarez-Quian et al, 1999; Wittliff and Erlander, 2002; Jacquet et al, 2005). Using double immunofluorescent staining with antibodies against vWF and GFAP, we showed reduction of GFAP mRNA levels in ECs and complete elimination of genes for endothelial markers in astrocytes, indicating that this protocol permits capture of a relatively pure population of ECs and astrocytes within brain tissue. In addition, our data show that GFAP mRNA was still present in ECs isolated from the vWF-positive and GFAP-negative vessels, suggesting the presence of astrocytic vascular end-foot processes which are not GFAP immunoreactive, as shown by Simard et al (2003). Real-time RT-PCR analysis revealed that mRNA levels in tissue samples acquired by this protocol were comparable to that in whole-brain tissue samples, indicating that the double-immunofluorescent staining protocol does not impair PCR efficiency, which is important for the ΔΔCt calculation, because the efficiency between the target and reference must be approximately equal. Collectively, our data show that the double-immunofluorescent staining protocol for LCM is sensitive for quantification of gene profiles in a relatively pure cell population from the brain tissue.