
Abstract
The recent identification of endogenous neural stem cells and persistent neuronal production in the adult brain suggests a previously unrecognized capacity for self-repair after brain injury. Neurogenesis not only continues in discrete regions of the adult mammalian brain, but new evidence also suggests that neural progenitors form new neurons that integrate into existing circuitry after certain forms of brain injury in the adult. Experimental stroke in adult rodents and primates increases neurogenesis in the persistent forebrain subventricular and hippocampal dentate gyrus germinative zones. Of greater relevance for regenerative potential, ischemic insults stimulate endogenous neural progenitors to migrate to areas of damage and form neurons in otherwise dormant forebrain regions, such as the neostriatum and hippocampal pyramidal cell layer, of the mature brain. This review summarizes the current understanding of adult neurogenesis and its regulation in vivo, and describes evidence for strokeinduced neurogenesis and neuronal replacement in the adult. Current strategies used to modify endogenous neurogenesis after ischemic brain injury also will be discussed, as well as future research directions with potential for achieving regeneration after stroke and other brain insults.
Neurogenesis in the Adult Mammalian Forebrain
The dogma that neuronal production is confined to a discrete period of development has been overturned by a vast sum of evidence collected over the past 15 years that adds irrefutable weight to early reports of ongoing neurogenesis in certain regions of the adult mammalian brain (Allen, 1912; Kershman, 1938; Smart, 1961; Altman, 1969; Altman and Das, 1965; Kaplan and Hinds, 1977). Neural stem-like cells capable of self-renewal and multipotency in vitro persist in the adult rodent forebrain (Reynolds and Weiss, 1992; Richards et al, 1992; Ray et al, 1995). Technical advances such as the use of bromodeoxyuridine (BrdU) and retroviral reporter mitotic labeling (e.g., Corotto et al, 1994; Lois and Alvarez-Buylla, 1994; Gage et al, 1995; van Praag et al, 2002) have facilitated unambiguous identification of adult-generated cells in vivo as new neurons. Adult neurogenesis persists in every mammalian species examined to date, including human and nonhuman primates (Eriksson et al, 1998; Gould et al, 1998, 1999; Kornack and Rakic, 2001; Pencea et al, 2001a; Bedard and Parent, 2004), in two forebrain regions: the dentate gyrus and subventricular zone (SVZ) (Figure 1).
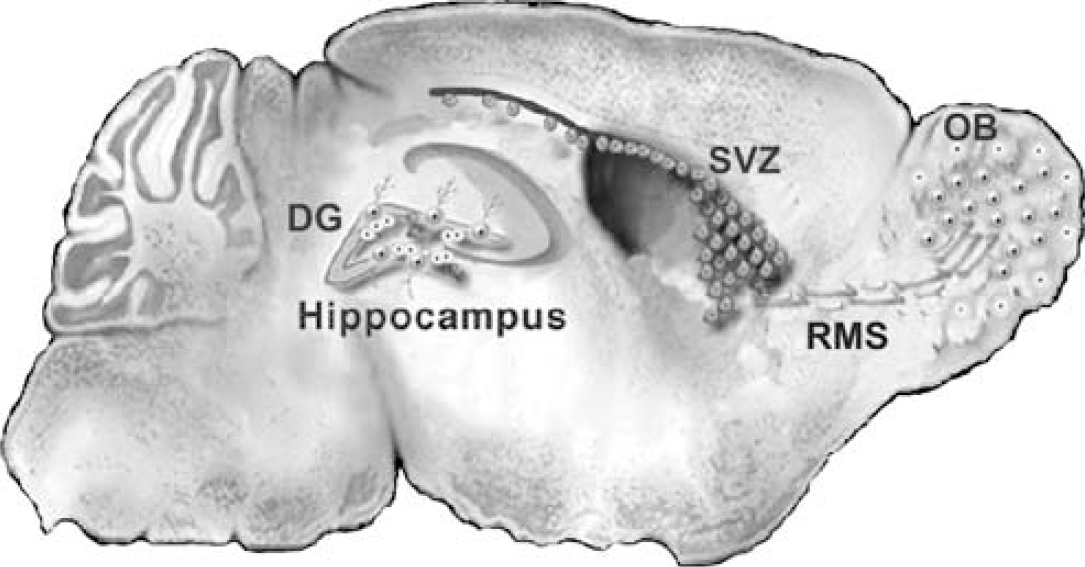
Schematic parasagittal view of the adult rodent brain. The two main regions of persistent neurogenesis in the forebrain are the hippocampal dentate gyrus (DG) and SVZ of the lateral ventricles. Neural progenitors under the DGC layer of the DG proliferate and generate DGCs throughout life. Progenitors in the striatal SVZ proliferate and give rise to neuroblasts that migrate tangentially in the RMS to their olfactory bulb (OB) target. Once they reach the OB, the neuroblasts disperse radially and differentiate into granule and periglomerular neurons.
Dentate Gyrus
Neuronal progenitors in the adult mammalian hippocampal dentate gyrus are located in the subgranular zone, at the border of the dentate granule cell (DGC) layer and hilus (Kaplan and Hinds, 1977; Cameron et al, 1993; Kuhn et al, 1996). The primary progenitor is a glial fibrillary acidic protein (GFAP)-expressing radial glia-like cell that gives rise to transit-amplifying progenitors which proliferate in clusters and generate neuroblasts (reviewed in Kempermann et al, 2004). The progeny disperse and migrate a short distance into the DGC layer where they differentiate into granule neurons (Cameron et al, 1993; Kuhn et al, 1996). Estimates suggest that approximately 9,000 DGCs are added to the young adult rodent dentate gyrus each day, and that newborn neurons comprise approximately 6% of the total population of DGCs (Cameron and McKay, 2001). Neurogenesis persists into senescence in the rodent, albeit at a lower rate in aged animals (Kuhn et al, 1996; see below).
Adult-born DGCs appear to functionally integrate into the circuitry of the dentate gyrus. They receive synaptic input (Kaplan and Hinds, 1977; Kaplan and Bell, 1983; Markakis and Gage, 1999; Carlén et al, 2002), send axonal projections to appropriate targets in hippocampal area CA3 (Stanfield and Trice, 1988; Hastings and Gould, 1999; Markakis and Gage, 1999), and acquire the electrophysiological properties of mature granule neurons (Wang et al, 2000; Song et al, 2002b; van Praag et al, 2002). Although the precise function of DGCs generated in adulthood is unknown, experimental depletion of new neurons disrupts the formation of some types of memory (Shors et al, 2001), suggesting that the new neurons play a role in hippocampal-dependent learning and memory.
Subventricular Zone
Neurogenesis also persists into adulthood in the mammalian forebrain SVZ. In adult rodents, SVZ neural stem cells give rise to neuroblasts that migrate to the olfactory bulb and differentiate into granule and periglomerular neurons (Altman, 1969; Corotto et al, 1993; Lois and Alvarez-Buylla, 1994). Similar to the DGC primary progenitor, the SVZ stem cell is a GFAP-immunoreactive astrocyte-like cell that probably arises from embryonic radial glia (Merkle et al, 2004). Neuroblasts generated in the SVZ migrate tangentially in chains surrounded by astrocytic ‘tubes' (Lois and Alvarez-Buylla, 1994; Lois et al, 1996; Thomas et al, 1996; Peretto et al, 1997) in the rostral migratory stream (RMS). On reaching the olfactory bulb subependymal region, they disperse radially to their final destinations in the granular and glomerular layers. Adult-born granular and periglomerular interneurons survive long term (Winner et al, 2002) and appear to integrate into bulb circuitry (Carlén et al, 2002; Carleton et al, 2003). As with adult-generated DGCs, the function of persistent olfactory bulb neurogenesis is largely unknown, but increasing evidence supports a role for the new neurons in olfactory memory or odor discrimination (Gheusi et al, 2000; Petreanu and Alvarez-Buylla, 2002; Rochefort et al, 2002).
Neurogenesis in Adult Human and Nonhuman Primates
Dentate granule cells continue to be generated throughout life in adult human and nonhuman primates. Several groups have shown neuronal birth in the dentate gyrus of Old and New World monkeys by showing that cells incorporating the S-phase markers tritiated thymidine or BrdU differentiate into neurons several weeks later (Gould et al, 1998, 1999; Kornack and Rakic, 1999). In the human dentate gyrus, new DGCs continue to be generated even as late as the seventh or eighth decades of life (Eriksson et al, 1998). Neuronal production also appears to continue in the adult SVZ-olfactory bulb pathway of primates (Kornack and Rakic, 2001; Pencea et al, 2001a). The adult primate SVZ underlies the ventral and lateral walls of the lateral ventricle. Subventricular zone neuroblasts migrate in the RMS as a thick chain surrounded by an astrocytic net (Pencea et al, 2001a). Interestingly, the adult human forebrain subependymal region appears to maintain neural stem-like cells, but chain migration of neuroblasts away from the lateral ventricles is not apparent (Sanai et al, 2004). Evidence suggests, however, that new neurons are added to the adult human olfactory bulb (Bedard and Parent, 2004) and their source therefore is uncertain.
Neurogenic Potential in Other Regions of the Mature Brain
Ongoing production of DGCs and olfactory bulb interneurons is widely accepted, but whether new neurons are added to other brain regions in adulthood remains a topic of debate. Reports of persistent neurogenesis in the rodent cortex date as far back as several decades ago (Altman, 1963; Kaplan, 1981), but more contemporary studies both in rodents and primates (Gould et al, 1999; Kornack and Rakic, 2001; Bernier et al, 2002; Koketsu et al, 2003; Dayer et al, 2005) thus far have failed to reach a consensus on the issue. Additional reports of new neurons in the adult striatum (Bedard et al, 2002; Dayer et al, 2005), substantia nigra (Zhao et al, 2003), and amygdala (Bernier et al, 2002) suggest that low-level neurogenesis in the adult mammalian brain might be a widespread phenomenon, but some of these studies are also in dispute (Cooper and Isacson, 2004; Frielingsdorf et al, 2004).
Regulation of Adult Neurogenesis
Many stimuli, exogenously applied agents, and endogenous factors or states appear to influence neurogenesis in the mature brain (Table 1). These interactions will first be described with regard to neurogenesis in the intact brain. Subsequent sections will discuss factors regulating neurogenesis in the ischemic brain.
In vivo modulators of adult neurogenesis in the intact and ischemic rodent forebrain
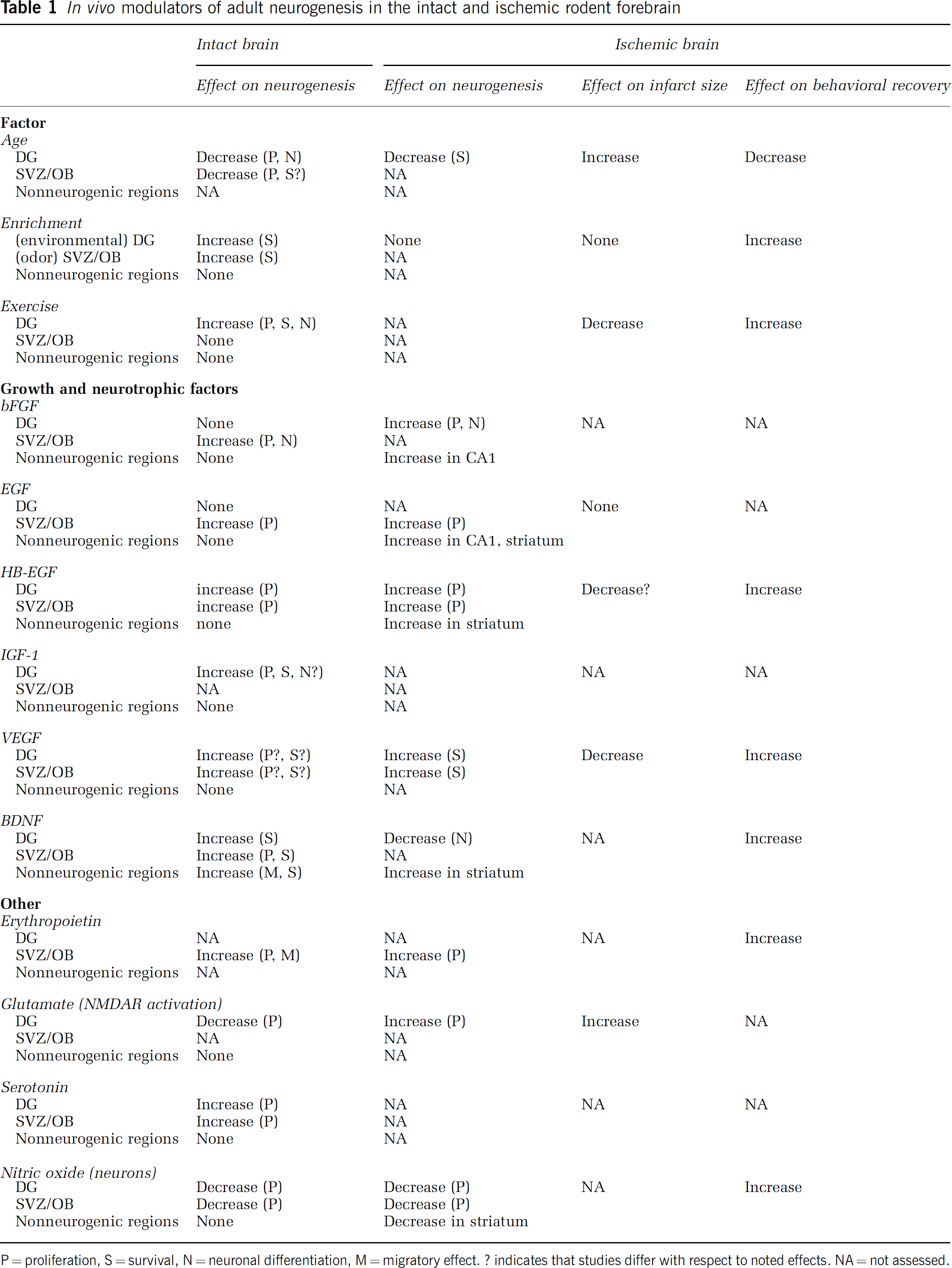
P = proliferation, S = survival, N = neuronal differentiation, M = migratory effect. ? indicates that studies differ with respect to noted effects. NA = not assessed.
Age
Although neurogenesis persists in the dentate gyrus and SVZ-olfactory bulb pathway throughout adulthood, the rate of neuronal production declines with age (Seki and Arai, 1995; Kuhn et al, 1996; Tropepe et al, 1997; Enwere et al, 2004). The decline is largely attributable to a reduction in proliferation of neuronal precursor cells, although evidence also implicates a decrease in neuronal fate determination (e.g., Kempermann et al, 1998) or slowed maturation of adult-born neurons in senescence (Rao et al, 2005). The age-dependent decline in neurogenesis appears to be mediated by age-related alterations in the cellular microenvironment rather than impaired responsiveness of progenitor cells to neurogenic stimuli. Tropepe et al, (1997) observed no difference between young adult and aged mice in the number or size of neurospheres derived from SVZ progenitors, a quantitative assay of mitotic activity. More recently, Enwere et al, (2004) depleted constitutively proliferating cells in the SVZ by infusing the antimitotic agent cytosine arabinoside (AraC) into the lateral ventricles of aged mice. They observed complete repopulation of SVZ progenitors, albeit at a slower rate than that observed in young adults, suggesting that the intrinsic proliferative capacity of SVZ progenitors is preserved throughout adulthood and that the decline in neurogenesis with age is likely due to changes in the local cellular environment. These groups further showed that aging is associated with a decline in SVZ expression of epidermal growth factor (EGF) receptor (Enwere et al, 2004). Young adult mice deficient in transforming growth factor-α (TGF-α), the predominant endogenous ligand for the EGF receptor, display senescent-like impairment in SVZ proliferation (Tropepe et al, 1997; Enwere et al, 2004) and olfactory neurogenesis and behavior (Enwere et al, 2004). Additional studies have shown that manipulating levels of growth factors and hormones in old animals profoundly influence neurogenesis. Aging is associated with elevated basal levels of corticosteroids (Sapolsky, 1992), and removal of corticosteroids by adrenalectomy significantly enhances DGC production in aged rats (Cameron and McKay, 1999; Montaron et al, 1999). Direct infusion of various growth factors, including heparin-binding EGF-like growth factor (HB-EGF), basic fibroblast growth factor (bFGF, also known as FGF-2), and insulin-like growth factor-1 (IGF-1), into the senescent brain reverses the age-related decline in neurogenesis in the dentate gyrus (Lichtenwalner et al, 2001; Jin et al, 2003a).
Environmental Enrichment and Exercise
Environmental enrichment increases the behavioral performance of adult rodents as well as the survival and neuronal differentiation of new DGCs generated in early adulthood (Kempermann et al, 1997; Nilsson et al, 1999), ‘middle age’ (Kempermann et al, 2002), and senescence (Kempermann et al, 1998). Exercise might be the predominant neurogenic component of environmental enrichment, given that physical activity (i.e., voluntary wheel running) alone enhances adult neurogenesis (van Praag et al, 1999). Environmental enrichment and exercise promote neurogenesis in the dentate gyrus, but appear to have no effect on neuronal addition to the olfactory bulb (Brown et al, 2003). Odor enrichment, conversely, has no effect on DGC production but enhances the survival of newly generated neurons in the olfactory bulb (Rochefort et al, 2002), and odor deprivation via unilateral naris closure reduces the number of surviving adult-born olfactory interneurons (Corotto et al, 1994). Growth factors might have a significant role in mediating these effects, since environmental enrichment increases hippocampal levels of glial cell line-derived neurotrophic factor (GDNF) and brain-derived neurotrophic factor (BDNF; Young et al, 1999), and blocking brain uptake of IGF-1 or vascular endothelial growth factor (VEGF) prevents the increase in DGC production induced by enrichment or exercise (Trejo et al, 2001; Fabel et al, 2003; Cao et al, 2004).
Growth and Neurotrophic Factors
Among the first growth factors shown to influence adult neurogenesis are bFGF and EGF. Neural stem cells isolated from the adult SVZ proliferate in vitro in response to the factors (e.g., Reynolds and Weiss, 1992; Richards et al, 1992; Palmer et al, 1995). Subcutaneous administration of bFGF stimulates mitotic activity in the neonatal and adult rodent SVZ (Wagner et al, 1999), and intracerebroventricular (ICV) infusion of bFGF or EGF increases proliferation in the adult SVZ (Craig et al, 1996; Kuhn et al, 1997). In addition, HB-EGF exerts a proliferative effect on neural progenitors in the adult dentate gyrus and SVZ (Jin et al, 2003a). In contrast to a similar mitogenic effect of the factors, bFGF treatment enhances neuronal addition to the adult olfactory bulb, whereas EGF predisposes SVZ precursor progeny to glial differentiation (Kuhn et al, 1997).
Brain-derived neurotrophic factor has been shown to increase adult SVZ neuronal production in EGFexpanded neurosphere and explant cultures, either via effects on differentiation or survival (Kirschenbaum and Goldman, 1995; Ahmed et al, 1995; Arsenijevic and Weiss, 1998). Brain-derived neurotrophic factor and bFGF also enable neurons to be generated and maintained in vitro from adult human SVZ (Pincus et al, 1998; Goldman, 1998). Recent in vivo studies of the effect of BDNF on the adult rat SVZ-olfactory bulb pathway have found that ICV infusion of the neurotrophin stimulates the production, migration, or survival of adult-generated olfactory bulb neurons (Zigova et al, 1998). Consistent with this finding, evidence from BDNF knockout mice suggests that it serves as a survival factor for postnatally generated olfactory bulb and dentate granule neurons (Linnarsson et al, 2000). Strikingly, ICV delivery of BDNF, either via osmotic minipump or adenoviral transduction, increases adult rat forebrain neurogenesis not only in the olfactory bulb but also in otherwise dormant regions including the neostriatum (Benraiss et al, 2001; Pencea et al, 2001b). Intracerebroventricular coinjection of adenoviral BDNF and the bone morphogenetic protein antagonist noggin significantly enhances neuronal addition to the adult rat striatum compared with BDNF alone, suggesting that endogenous progenitors are mobilized optimally when permissive and instructive cues are concurrently engaged (Chmielnicki et al, 2004).
The angiogenic factor VEGF has recently emerged as an important regulator of adult neurogenesis. Intracerebroventricular infusion of VEGF increases neurogenesis in the rodent dentate gyrus and SVZ, but it is not yet clear whether this effect results from increased cell proliferation (Jin et al, 2002), survival (Schänzer et al, 2004), or both (Cao et al, 2004). Neural stem cells in the SVZ (Schänzer et al, 2004) and immature, doublecortin-expressing neuroblasts in the dentate subgranular zone (Jin et al, 2002) express the kinase insert domain protein receptor (KDR, also known as VEGFR2 and Flk1). VEGF appears to modulate adult neurogenesis by signaling through this receptor, since administration of a dominant-negative mutant KDR reverses the VEGF-induced increase in neurogenesis (Cao et al, 2004).
Other growth factors and cytokines that modulate adult neurogenesis include TGF-α (Tropepe et al, 1997), IGF-1 (Åberg et al, 2000), and erythropoietin (Shingo et al, 2001). Although the stimulatory effect of many growth and neurotrophic factors on adult neurogenesis is well documented, the potential adverse consequences of in vivo treatment with these factors are less clear. Recently, one study has raised the possibility that EGF receptor expression may contribute to glioma formation by SVZ progenitors (Doetsch et al, 2002), and another report has shown that BDNF infusion into the hippocampus induces spontaneous limbic seizures in approximately 25% of treated rats (Scharfman et al, 2002). Clearly, the development of neurogenic therapies to treat ischemia and other brain insults will require a more thorough understanding of the range of side effects caused by growth factor administration.
Neurotransmitters and Other Factors
Additional mediators of neurogenesis in the adult mammalian forebrain include multiple hormones and neurotransmitters. Early studies of adult neurogenesis identified adrenal steroids as a negative regulator of mitotic activity in the dentate gyrus. Adrenalectomy and corticosteroid replacement studies have shown that circulating stress hormones suppress the rate of DGC production in adulthood (Gould et al, 1992; Cameron and Gould, 1994). In contrast, cell proliferation in the rat dentate gyrus (Tanapat et al, 1999) and prairie vole SVZ—RMS pathway (Smith et al, 2001) increases during estrus, and the effect is abolished by ovariectomy (Tanapat et al, 1999; Smith et al, 2001). Subventricular zone proliferation and olfactory neurogenesis is also enhanced during pregnancy in mice, and the upregulation is mediated by the hormone prolactin (Shingo et al, 2003).
Glutamatergic mechanisms also regulate neurogenesis in the adult rodent dentate gyrus. Activation of N-methyl-
The transmitter serotonin (5-hydroxytryptamine) also influences adult neurogenesis. Chronic treatment with antidepressant drugs that act as serotonin reuptake inhibitors increases cell proliferation in the rodent dentate gyrus (Malberg et al, 2000), and blockade of serotonin synthesis or administration of a serotonin neurotoxin decreases new cell production in the dentate gyrus and SVZ (Brezun and Daszuta, 1999). Moreover, antidepressantinduced DGC neurogenesis, as well as improved performance on measures of anxiety-related behavior, is abolished in serotonin 1A receptor null mice and in mice in which hippocampal progenitors are eliminated in adulthood by irradiation, suggesting that adult neurogenesis plays a role in the behavioral effects of antidepressant treatment (Santarelli et al, 2003).
Nitric oxide synthase (NOS) inhibitors increase SVZ proliferation and delay neuronal maturation of SVZ precursor progeny that migrate to the olfactory bulb (Cheng et al, 2003; Packer et al, 2003; Moreno-Lopez et al, 2004). The number of newly generated cells in the olfactory bulb and dentate gyrus also is enhanced significantly in null mutant mice lacking neuronal NOS activity (Packer et al, 2003). These studies implicate nitric oxide as a negative regulator of cell proliferation and a positive promoter of neuronal fate determination in neurogenic regions of the adult brain. Interestingly, treating cultured cortical neuroepithelial cells with NOS inhibitors abolishes the stimulatory effect of BDNF on neuronal differentiation (Cheng et al, 2003), suggesting that nitric oxide acts in tandem with other endogenous factors to mediate various phases of neurogenesis. Neuronal addition thus continues in adulthood under the regulation of a fine balance between abundant suppressors and stimulants of neurogenic activity.
The Neurogenic Niche
The local microenvironment in regions of ongoing neurogenesis in the adult mammalian brain provides what is referred to as a ‘stem cell niche’ which governs neuronal production and includes components such as astrocytes, microvasculature, and microglia in addition to progenitor cells (Wurmser et al, 2004). Astrocytes appear to influence all phases of adult neurogenesis, including neural precursor cell proliferation, migration, and differentiation. Astrocytes promote neuroblast proliferation and neuronal differentiation in cultured SVZ stem cells and hippocampal progenitors (Lim and Alvarez-Buylla, 1999; Song et al, 2002a). In addition, the RMS structural arrangement of neuroblast chain migration through astrocytic tubes suggests that astrocytes direct migration of neuroblasts from the SVZ to the olfactory bulb (Thomas et al, 1996; Peretto et al, 1997). Studies of neurogenesis in the adult rodent dentate gyrus and SVZ implicate radial glial cells or radial glia-like astrocytes as the neural stem cells from which neuroblasts derive (Doetsch et al, 1999; Seri et al, 2001; Merkle et al, 2004). Recently, Garcia et al, (2004) selectively ablated dividing cells in the adult brain that expressed the astrocytic protein GFAP. To accomplish the targeted ablation, they delivered the antiviral agent ganciclovir to transgenic mice expressing the ‘suicide gene’ herpes simplex virus thymidine kinase (HSV-TK) under the control of the GFAP promoter. Cessation of adult neurogenesis in the dentate gyrus and SVZ-olfactory bulb pathway in ganciclovir-treated GFAPHSV-TK mice showed that adult-generated neurons originate from GFAP-expressing progenitor cells (Garcia et al, 2004).
In addition to the critical role of astrocytes in adult neurogenesis, recent evidence shows a vascular presence in the stem cell niche. Neuronal precursors in the dentate gyrus and SVZ reside in close proximity to local microvasculature (Palmer et al, 2000; Capela and Temple, 2002), and their proliferative activity appears to be tightly linked with angiogenesis. Using cocultures of endothelial cells and neural progenitors, Shen et al, (2004) have shown that soluble factors released by endothelial cells potently stimulate neural stem cell expansion and neuronal production from embryonic ventricular zone and postnatal SVZ. In the adult, suppression of neurogenesis by irradiation may reflect disruption of local angiogenesis (Monje et al, 2002), and infusion of VEGF into the adult brain increases the production both of endothelial cells and DGCs (Jin et al, 2002). Steroid-induced, selective expansion of the microvasculature in the higher vocal center of adult canaries enhances neurogenesis in this region, potentially via elevated levels of local BDNF (Louissaint et al, 2002). Moreover, stem cells isolated from the adult rodent SVZ and grown in endothelial coculture display enhanced self-renewal and increased neurogenesis (Leventhal et al, 1999; Shen et al, 2004), further suggesting that the vascular endothelium is an important component of the neurogenic niche.
A third cell type, activated microglia, becomes an important member of the stem cell niche in the injured brain. Microglial activation occurs in response to numerous insults, including seizures, infection, and irradiation (Sloviter, 1999; Kempermann and Neumann, 2003). Recent studies suggest that reactive microglia disrupt neurogenesis in the adult dentate gyrus. Infusion of bacterial lipopolysaccharide (LPS) induces inflammation, stimulates reactive gliosis, and results in an 85% decrease in the survival of newborn DGCs (Ekdahl et al, 2003). The negative correlation between the number of activated microglia and the number of surviving new DGCs is reversed by treatment with minocycline, an inhibitor of microglial activation. Irradiation-induced inflammation also suppresses neurogenesis in the adult rodent dentate gyrus, and blocking inflammation with the drug indomethacin partially restores neurogenesis (Monje et al, 2003).
Influence of Brain Injury on Adult Neurogenesis
The persistence of neural stem cells in the adult mammalian forebrain raises the possibility for endogenous neuronal renewal in the setting of brain injury or degeneration. Several important questions must be addressed, however, in assessing prospects for endogenous repair. First, does a low level of self-repair exist in the adult mammalian brain? Second, will augmenting neurogenesis after brain insults improve recovery? Third, does the potential exist for neurogenesis to produce maladaptive consequences after brain injury? Finally, will findings in animal models of brain injury or degeneration be applicable to the development of therapies to augment endogenous brain repair in humans? We will discuss recent work aimed at answering some of these questions in the sections below, with a focus on experimental stroke.
Newly generated neurons in the adult must integrate and form long-distance connections for regeneration and functional restoration to occur after stroke or other brain insults. Is this possible? Using a form of ‘sterile injury,’ studies by Jeff Macklis and colleagues provide evidence that this is indeed the case. They produced selective apoptosis of neocortical layer VI pyramidal neurons via retrograde transport of a photolytically activated toxin in adult mice, and found that apoptotic cell death stimulates neurogenesis in the damaged neocortical layer (Magavi et al, 2000). More importantly, they used combined mitotic and retrograde tracer labeling to show that the newly generated cells exhibited pyramidal neuron morphology and a small percentage formed long-distance corticothalamic connections. The group recently showed that endogenous neuroblasts can also replace ablated corticospinal projection neurons in juvenile mice and that, remarkably, a portion of new neurons extends axons to the spinal cord (Chen et al, 2004). Although the presumed SVZ origin of the precursors is yet to be confirmed and the model is not associated with the tissue reaction typical of brain injury, the findings strongly support the contention that newly born neurons in the adult are able to integrate and form long-distance projections. These data raise optimism for the reparative potential of endogenous progenitors; however, the proportion of newborn neurons integrating over long distances is very low. Moreover, the functional consequences of neurogenesis after injury remain to be determined.
In terms of more realistic brain injury models, several consistent findings emerge across different models and for the two main regions of persistent neurogenesis. Nearly all forms of injury in the adult increase cell proliferation in the hippocampal dentate gyrus and forebrain SVZ (Figure 2). For example, damage induced by mechanical lesions, prolonged seizures, hypoglycemia, fluid percussion injury or stroke increases dentate gyrus cell proliferation by approximately 5- to 12-fold. As in the intact brain, many newly generated cells die, but the vast majority of those that survive differentiate into neurons, and typically less than 20% of adult-born cells become glia or remain undifferentiated (Gould and Tanapat, 1997; Parent et al, 1997; Liu et al, 1998; Jin et al, 2001; Rice et al, 2003; Suh et al, 2005). Proliferating cell numbers increase approximately 1.5- to 3-fold in the rodent striatal SVZ after various injuries, including cortical aspiration (Szele and Chesselet, 1996), cortical transection (Willis et al, 1976), fimbria fornix lesions (Weinstein et al, 1996), inflammatory demyelination (Calzà et al, 1998), seizures (Parent et al, 2002a), stroke (Zhang et al, 2001b; Arvidsson et al, 2002; Parent et al, 2002b), excitotoxic striatal lesions (Tattersfield et al, 2004), and fluid percussion injury (Chirumamilla et al, 2002).
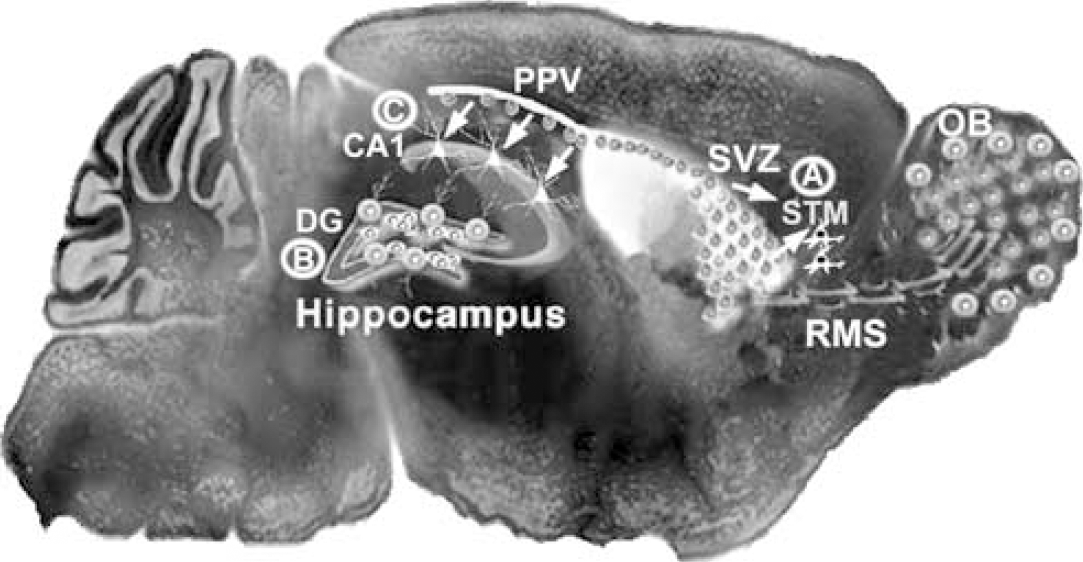
Modification of the schematic representation in Figure 1, showing regions of injury-induced neurogenesis. Many forms of injury increase cell proliferation and neurogenesis in the SVZ, and focal ischemic insults recruit SVZ progenitors (arrows) to form new neurons in the striatum (STM;
The phenotypic potential of adult rodent SVZ precursors after injury also has been explored more recently. Progenitors that migrate out of the SVZ or RMS toward sites of demyelinating or mechanical injury appear to differentiate into oligodendrocytes or astrocytes in some disease models (Holmin et al, 1997; Nait-Oumesmar et al, 1999). In contrast, transection lesions through the RMS induce ectopic neural precursor migration caudal to the lesion, perhaps by blocking the cells' ability to reach the olfactory bulb, and the progenitors migrating along the glial scar exhibit a neuronal phenotype and express GABA and calretinin (Alonso et al, 1999). Using the 6-hydroxydopamine lesion model of Parkinson's disease, Fallon et al, (2000) found that proliferating cell numbers increase in the striatal SVZ ipsilateral to injury. The neuroblasts migrate into the striatum and form neurons exclusively in lesioned animals that also receive striatal infusion of TGF-α. Recent attempts to replicate this finding, however, have failed to show a neuronal phenotype of the newborn striatal cells (Cooper and Isacson, 2004). Nonetheless, the large body of evidence now accumulated indicates that SVZ progenitors increase proliferation in response to forebrain injury. The progeny, moreover, are able to exit the normal migratory pathway, home to damaged regions, and differentiate into neurons or glia.
Another consistent finding is that similar injuries produce comparable effects on both the striatal SVZ and hippocampal dentate gyrus germinative zones. For example, damage caused by trauma, seizures or focal ischemia accelerates cell proliferation in both the dentate gyrus and SVZ-olfactory bulb pathway of adult rodents (Parent et al, 1997; Liu et al, 1998; Jin et al, 2001; Zhang et al, 2001b; Chirumamilla et al, 2002; Chen et al, 2004; Rice et al, 2003). Precursor progeny give rise mainly to neurons in both regions, although this finding is likely biased in part by the timing of BrdU administration relative to injury; proliferating astrocytes, microglial and endothelial cells also are found (Liu et al, 2001; Parent et al, 2002b). An important difference between the two regions, however, involves the probable consequences of increased neurogenesis. In some stroke models (described below), increased SVZ neurogenesis appears to be associated with cell replacement in injured striatum (Arvidsson et al, 2002; Parent et al, 2002b). No such evidence exists to support a similar cell replacement in the hippocampal dentate gyrus after stroke or other cerebral insults. Injury-induced dentate gyrus neurogenesis has not been shown to produce any newborn neurons other than DGCs, a principal neuronal population often preserved in injury models. Moreover, studies of seizure-induced neurogenesis in adult rat suggest that DGC neurogenesis is altered in the epileptic hippocampal formation (Parent et al, 1997; Scharfman et al, 2000). Newborn DGCs integrate in ectopic locations and are abnormally hyperexcitable, features that may contribute to the epileptic phenotype. An important challenge for studies of injury-induced neurogenesis therefore will be not only to show that newborn neurons integrate normally, but also that they preserve or improve network integrity and behavioral function.
stroke-induced Neurogenesis and Cell Replacement
stroke-induced Dentate Granule Cell Neurogenesis
Accumulating evidence suggests that ischemic injury dramatically increases neurogenesis in mitotically active regions of the adult rodent brain (Figure 2). Liu et al, (1998) first described a greater than 10-fold increase in cell proliferation in the dentate gyrus after transient global ischemia in the adult gerbil. Accelerated precursor proliferation is a transient phenomenon that occurs after a latent period, and the majority of surviving cells differentiates into DGCs (Liu et al, 1998). Additional studies of neurogenesis in the dentate gyrus in adult mice (Takagi et al, 1999; Tureyen et al, 2004) and rats (Jin et al, 2001; Kee et al, 2001; Yagita et al, 2001; Takasawa et al, 2002; Choi et al, 2003) confirm that transient focal or global ischemia potently stimulates proliferation, which peaks 7 to 10 days after ischemia and returns to baseline levels within several weeks. Many of the new cells die, but the percentage of surviving cells that adopt a neuronal fate is similar in the ischemic and uninjured dentate gyrus. Recently, Tanaka et al, (2004) injected an enhanced green fluorescent protein (EGFP) retroviral reporter into the dentate gyrus in adult gerbils 48 h before ischemia and then analyzed EGFP-labeled cells at 5, 10, and 30 days after ischemia. With increasing time after ischemia, EGFP-positive cells moved from the subgranular proliferative zone to the DGC layer, shifted from coexpression of immature to mature neuronal markers, and increased in measures of dendritic length (Tanaka et al, 2004), suggesting that newly generated DGCs in the ischemic brain follow a time course of neuronal maturation similar to that in normal animals (van Praag et al, 2002). Kawai et al, (2004) observed intracellular signaling in new DGCs in the ischemic rat dentate gyrus; ICV injection of NMDA 1 or 2 months after ischemia induced phosphorylation of ERK in BrdU-labeled DGCs, and the number of cells responsive to NMDA stimulation increased with time after ischemia. These studies collectively suggest that ischemia-induced neurogenesis in the adult dentate gyrus produces mature DGCs with the capacity for neuronal function. The relevance of strokeinduced DGC neurogenesis to repair after injury, however, remains uncertain.
stroke-induced Subventricular Zone Neurogenesis
Evidence from a number of investigators indicates that forebrain ischemic injury increases SVZ cell proliferation and neurogenesis. Using an embolic middle cerebral artery occlusion (MCAO) model in adult rats, Zhang et al, (2001b) have shown increased SVZ BrdU labeling and neurogenesis that peaks 7 days after ischemia. Subventricular zone neurogenesis also increases in the adult rat SVZ after transient MCAO (tMCAO (Jin et al, 2001)). The ischemia-induced neurogenesis is short-lived, and neither of these reports identifies persistent neurogenesis in periinfarct regions. More recent studies, however, provide important evidence that endogenous precursors can generate neurons after stroke in otherwise dormant regions of the adult rat forebrain. Using the adult rat tMCAO model, two groups have found that focal ischemia markedly increases forebrain SVZ neurogenesis ipsilateral to the infarct (Arvidsson et al, 2002; Parent et al, 2002b). The expanded neuroblast population also appears to be diverted to injured regions, as they extend from the SVZ to the neostriatum in migratory chains closely apposed to astrocytes (Figure 3). This migratory appearance is similar to the normal RMS chain migration of SVZ neuroblasts to the olfactory bulb (Lois et al, 1996; Peretto et al, 1997) and suggests that SVZ neuroblasts are redirected to migrate into periinfarct regions by cues induced by ischemic injury or related to astrocyte proliferation. Similar ectopic striatal migration of SVZ progenitors after focal ischemia also has been shown by labeling SVZ cells with the lipophilic carbocyanine tracer DiI (1, 1'-dioctadecyl-3,3,3',3'-tetramethylindocarbocyanine perchlorate) (Jin et al, 2003b).
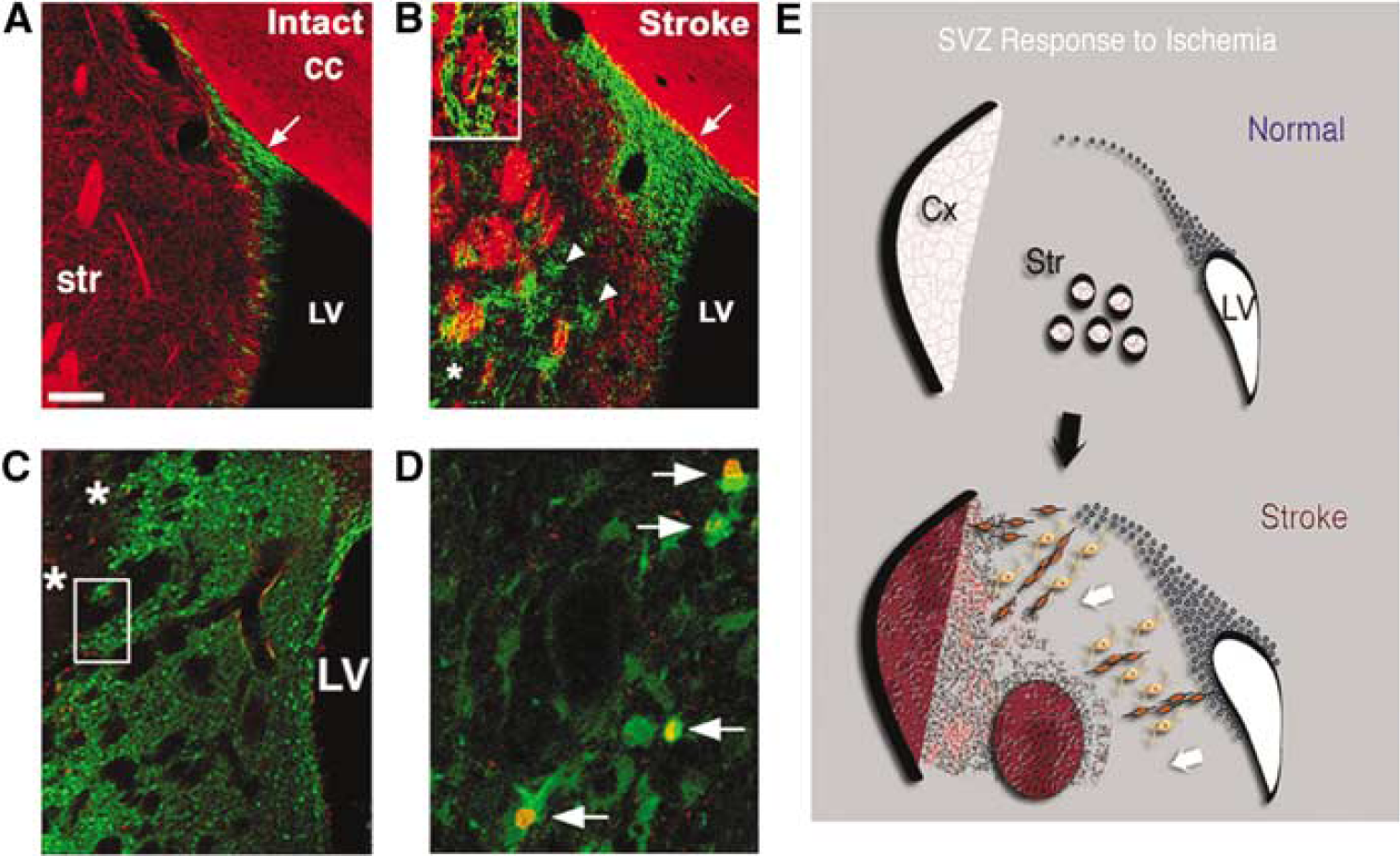
Confocal images of coronal sections through the SVZ and striatum of the adult rat brain after transient middle cerebral artery occlusion (tMCAO), showing uninjured (
An important issue is whether neural progenitors that migrate to injured regions after stroke differentiate into regionally appropriate types of neurons. After tMCAO, SVZ neuroblasts that migrate to the injured striatum express markers of medium spiny neostriatal projection neurons (Arvidsson et al, 2002; Parent et al, 2002b), suggesting that they have the potential to replace neurons lost after focal ischemia. Although some of the newly generated striatal neurons persist for up to 5 weeks after tMCAO, it seems that the majority fails to survive (Arvidsson et al, 2002; Parent et al, 2002b), and estimates suggest that the number of surviving adult-born striatal neurons comprises only ∼0.2% of the fraction of striatal neurons that die in response to the ischemic injury (Arvidsson et al, 2002). A smaller number of neuroblasts appear in the periinfarct neocortex after stroke (Arvidsson et al, 2002; Parent et al, 2002b; Jin et al, 2003b; Zhang et al, 2004). Whether these new cells arise from the SVZ or from intraparenchymal progenitors is uncertain. Moreover, progenitors do not appear to differentiate into neurons and persist in the periinfarct neocortex after stroke, although the possibility that ischemia induces mature cortical neurons to become mitotically active has been suggested (Jiang et al, 2001).
Even for putative striatal neuron replacement after focal ischemia, important questions remain to be addressed. For example, whether the new neurons generated after stroke integrate and contribute to functional recovery is unknown. To address the former question, we have begun to examine if newly generated neostriatal neurons extend long-distance projections using combined BrdU and FluoroGold (FG) retrograde tracer labeling. Injection of FG into the ipsilesional substantia nigra pars reticulata, a target of striatal projection neurons, 6 weeks after stroke labels medium spiny-like neurons adjacent to BrdU-labeled cells in the striatum (Figure 4); however, we have yet to find colocalization of BrdU within cells that retrogradely transported FG from this one potential target region. Nonetheless, accumulating data suggest that endogenous progenitors are capable of replacing some neurons damaged by focal ischemic injury in the adult, and raise the possibility that treatments to stimulate endogenous neurogenesis may improve stroke recovery.
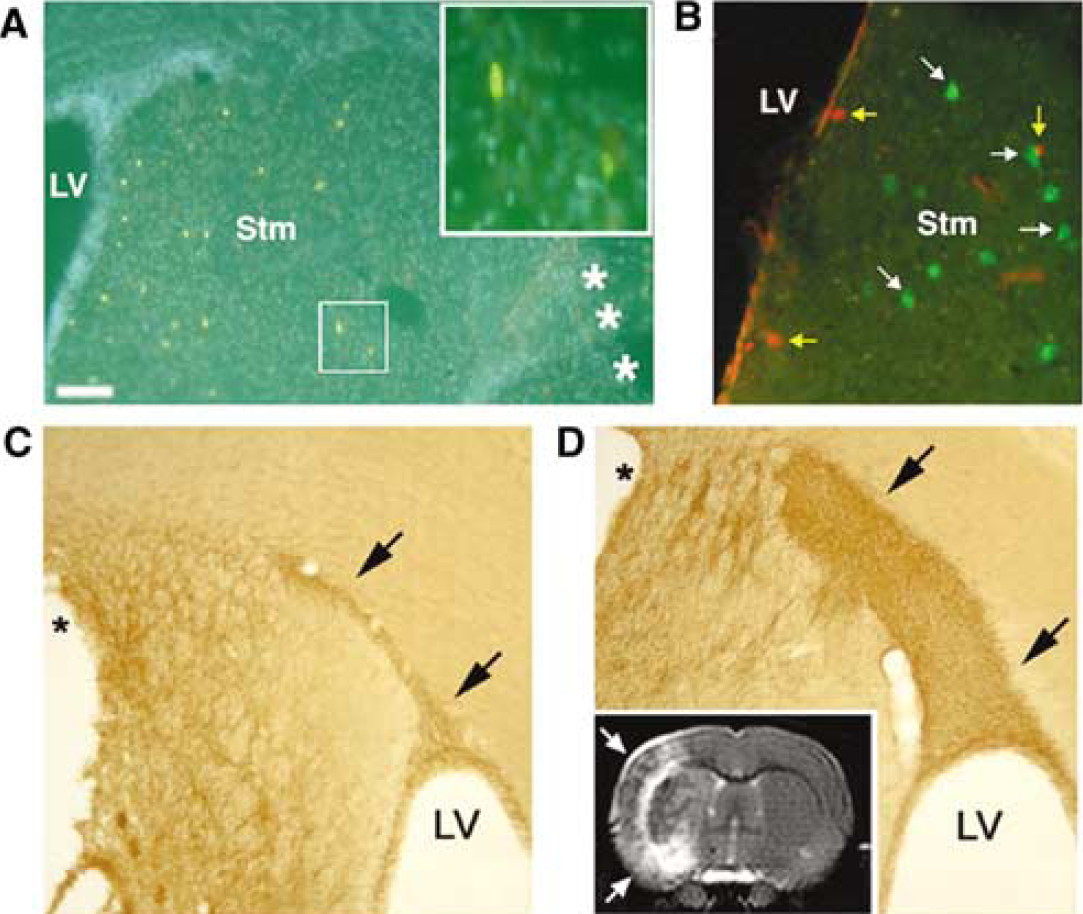
(
stroke-induced Neurogenesis in the Hippocampus Proper
Although hippocampal pyramidal neurons are generated exclusively during embryonic development in the rodent, recent work suggests that certain forms of ischemic brain injury reactivate this process in the adult. Nakatomi et al, (2002) have examined neurogenesis after inducing global ischemia by transient four-vessel occlusion and systemic hypotension in adult rats. This model produces a delayed apoptosis of hippocampal CA1 pyramidal cells, and the study provides evidence that a modest number of pyramidal cells regenerate within 4 weeks after stroke. Remarkably, infusion of bFGF and EGF intraventricularly in the first week after global ischemia leads to regeneration of nearly 40% of the damaged CA1 region at 28 days. DiI and retroviral reporter labeling shows that at least some of the newly generated neurons arise from the posterior periventricular region (Figure 2), migrate to the CA1 region after ischemia, and differentiate into electrophysiologically functional neurons with morphological characteristics of pyramidal cells (Nakatomi et al, 2002). Intriguingly, the growth factor-induced hippocampal neuronal replacement is associated with partial recovery of spatial learning deficits caused by global ischemia, although the contribution of the new neurons to the functional improvements remains to be proven.
Neurogenesis in the hippocampal pyramidal cell layer also has been reported in an adult gerbil model of transient global ischemia (Schmidt and Reymann, 2002). Other studies in the gerbil model have not found pyramidal neuron generation (Liu et al, 1998), suggesting that it is a rare event, at least without growth factor augmentation. Nevertheless, the presence of hippocampal pyramidal cell, as well as neostriatal, neurogenesis after stroke suggests that neuronal replacement by endogenous progenitors is possible in otherwise dormant brain regions. Moreover, pharmacological stimulation of neurogenesis after ischemic injury appears to promote neuronal renewal that may contribute to functional recovery.
Regulation of strokeinduced Neurogenesis
Many factors that regulate neurogenesis in the intact adult brain also play a role in the neurogenic response to ischemic injury (Table 1). In addition to mediating strokeinduced neurogenesis in the dentate gyrus and SVZ, a number of these factors facilitate mobilization of endogenous progenitors to areas of ischemic damage and also appear to influence stroke severity and functional outcome.
Age
Given that aging is a risk factor for ischemic injury and other neurological disorders, developing effective regenerative strategies may require a better understanding of how neural progenitors in the brain respond to insults during senescence. Cell proliferation in the adult dentate gyrus increases in response to transient forebrain ischemia both in young adult and aged rats (Yagita et al, 2001). In fact, the relative increase in mitotic activity after ischemia is actually greater in old rats compared with young adults. The survival of adult-born cells, however, decreases dramatically in the aged animals, such that the total number of newly generated cells 1 month after ischemia is approximately sevenfold lower in the older population (Yagita et al, 2001). In addition, migration of neuroblasts into the ischemic striatum also appears to be reduced in old rats compared with that in young adults (Jin et al, 2004a). Aged rats have larger ischemic infarcts (Davis et al, 1995; Kharlamov et al, 2000; Otsuka et al, 2000), higher mortality rates (Wang et al, 2003), and impaired functional recovery from focal cerebral ischemia compared with young adults (Badan et al, 2003). Deficits in ischemia-induced neurogenesis may contribute to impaired behavioral recovery. Raber et al, (2004) used irradiation to suppress neurogenesis in young adult gerbils and found that, in comparison with animals subjected to cerebral ischemia alone, irradiated animals that experienced ischemic injury display impaired performance on various behavioral tasks. Although confounding variables associated with irradiation complicate interpretation of these data, ongoing neurogenesis might be important for functional recovery from brain insults, and the age-dependent reduction in neurogenesis may impede recovery from injury. It remains to be seen whether treating old animals with growth factors or other neurogenic stimuli could facilitate functional recovery after stroke.
Neurogenesis in the hippocampal dentate gyrus and SVZ-olfactory bulb pathway peaks in the neonatal period. Unlike the decreased neurogenesis described for the aged rodent brain above, therefore, one may expect strokeinduced neurogenesis to be markedly increased in the neonate compared with young adult rodents. Unfortunately, this idea does not appear to hold true. Severe hypoxia—ischemia in neonatal rat or mouse actually depletes the SVZ progenitor pool (Brazel et al, 2004; Romanko et al, 2004). This depletion likely impairs the generation of neurons and glia from the postnatal SVZ. In contrast, Plane et al, (2004) showed that more moderate cerebral hypoxia—ischemia in neonatal mice markedly increases SVZ cell proliferation and neurogenesis (Figure 5). Chains of doublecortin-expressing neuroblasts extend from the SVZ toward the damaged striatum, as occurs after stroke in adult rodents, and many proliferating astrocytes appear in the periinfarct region. Unlike in the adult, however, the newly generated cells do not appear to generate mature neurons that survive in the lesioned striatum. Similar effects of moderate hypoxia—ischemia occur in the neonatal rat (Ong et al, in press). Differences in strokeinduced SVZ neurogenesis between neonatal and adult animals may reflect a more gliogenic environment in the immature brain, or the absence of a necessary survival factor(s) after hypoxic—ischemic injury in the neonate.
Environmental Enrichment and Exercise
Environmental enrichment improves functional outcome after tMCAO in adult rats. Although infarct size does not appear to be affected, enrichment significantly improves performance on several measures of motor behavior and cognitive function (e.g., Ohlsson and Johansson, 1995; Komitova et al, 2002; Dahlqvist et al, 2004; Gobbo and O'Mara, 2004). Postischemic enrichment is most effective soon after stroke (Biernaskie et al, 2004), and recovery of function is further enhanced by exposure to an enriched environment both before and after stroke (Ohlsson and Johansson, 1995). Postischemic enrichment has no apparent affect on dentate gyrus neurogenesis (Komitova et al, 2002), but an enriched environment after stroke, paired with daily rehabilitative training targeting fine digit and forelimb function, increases dendritic arborization of layer 5 pyramidal neurons within the contralateral forelimb motor cortex (Biernaskie and Corbett, 2001). Similar to the beneficial effect of environmental enrichment on stroke recovery, exercise improves functional outcome after ischemic brain injury. Daily treadmill training for several weeks before stroke (Wang et al, 2001; Ding et al, 2004) or during the first week after tMCAO (Yang et al, 2003) reduces infarct volume and neurological deficits. It is not clear whether the improvement conveyed by enrichment and exercise requires mobilization of endogenous progenitors for self-repair. Of note, however, is that ischemic rats exposed to exercise preconditioning have elevated cortical and striatal levels of nerve growth factor (NGF) and BDNF compared with nonexercised controls, in addition to increased microvessel density in the injured striatum (Ding et al, 2004). Several recent studies suggest a more direct role for growth factors, neurotrophins, and the vasculature in promoting endogenous neuronal renewal in response to stroke.
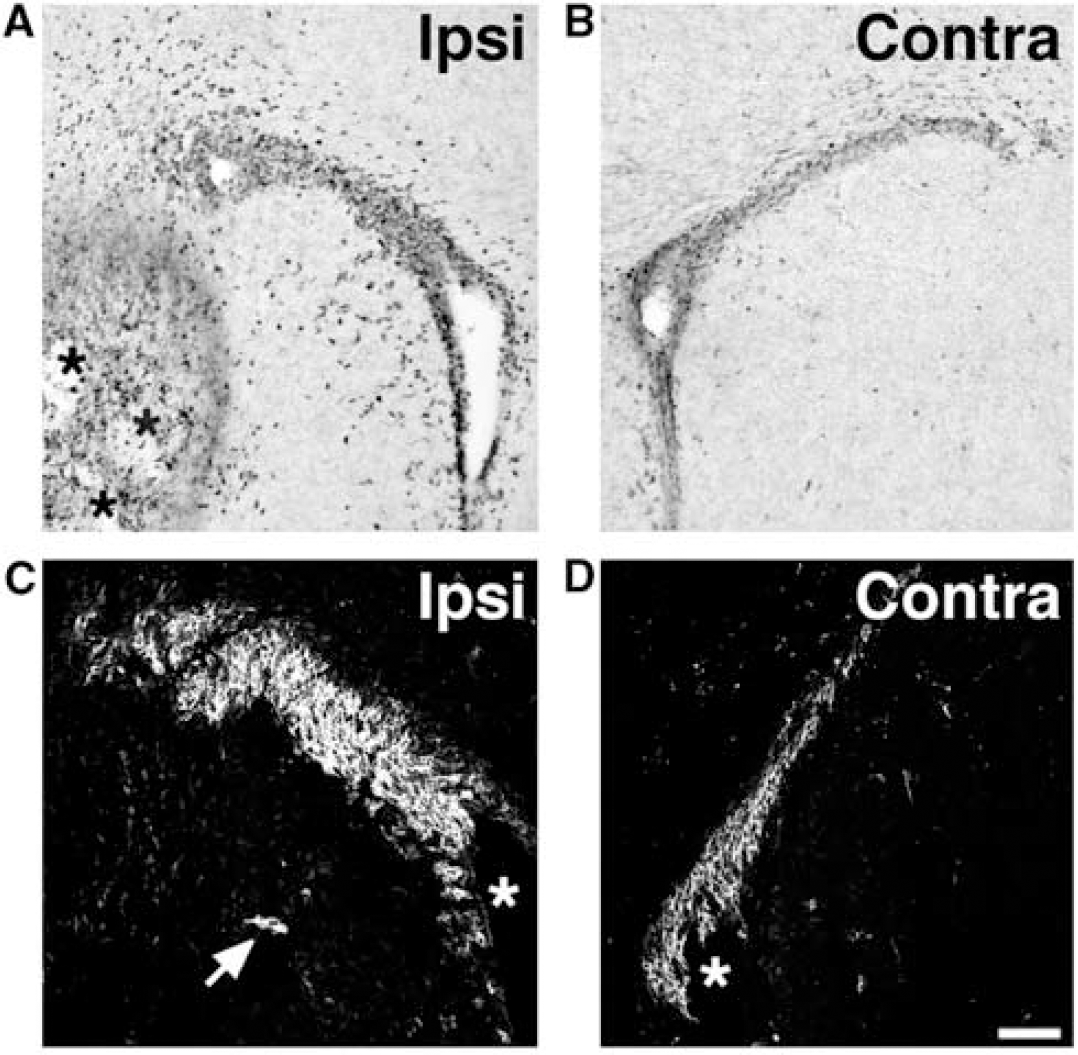
Subventricular zone cell proliferation and neurogenesis induced by focal hypoxic—ischemic insult in neonatal mice. Coronal sections through the striatal SVZ show BrdU (
Growth and Neurotrophic Factors
Growth or neurotrophic factors are candidates for mediating ischemia-induced neurogenesis and recruitment of endogenous progenitors to sites of damage. Forebrain ischemia increases the expression of several of these factors, including BDNF, bFGF, and GDNF (Takami et al, 1992; Kokaia et al, 1995; Lin et al, 1997; Kitagawa et al, 1999), and sustained upregulation of these factors may mediate strokeinduced neurogenesis and neuroblast migration. In the intact rodent brain, ICV delivery of BDNF induces neuroblast migration to nonneurogenic regions including the striatum, where the adult-generated cells differentiate into medium spiny neurons (Benraiss et al, 2001; Pencea et al, 2001b; Chmielnicki et al, 2004). Intrastriatal delivery of adenoviral BDNF 4 to 5 weeks before tMCAO in adult rats modestly increases survival of dorsolateral striatal neurons and improves functional outcome (Andsberg et al, 2002). Gustafsson et al, (2003) injected adenoviral BDNF into the substantia nigra 4 to 5 weeks before tMCAO and observed a dramatic increase in striatal BDNF levels and in the neurogenic response to stroke. The authors found that BDNF treatment approximately doubles the number of cells expressing doublecortin, a marker of migrating neuroblasts, and triples the number of newly generated neurons in the striatum ipsilateral to MCAO (Gustafsson et al, 2003).
Other candidates for mediating ischemia-induced neurogenesis and neuronal repair after stroke include GDNF, bFGF, EGF, nitric oxide donors, and VEGF. Intrastriatal GDNF treatment decreases death of striatal neurons (Hermann et al, 2001), and cortical application of the factor reduces infarct size after ischemic injury (Kitagawa et al, 1998; Tsai et al, 2000). Intracerebroventricular infusion of GDNF increases cell proliferation in the dentate gyrus and SVZ after stroke (Dempsey et al, 2003), suggesting a potential role for the factor in mobilizing endogenous progenitors to respond to injury. More recent evidence, however, suggests that the beneficial effect of GDNF on stroke recovery might be transient; experimental elevation of GDNF levels in the striatum has no apparent effect on survival of striatal neurons or functional outcome, assessed 5 to 8 weeks after stroke (Arvidsson et al, 2003).
To determine the role of endogenous FGF-2 (bFGF) on the neurogenic response to injury, Yoshimura et al, (2001) performed tMCAO on mice genetically deficient in FGF-2 and in wild-type controls. Significantly fewer adult-born DGCs are apparent in FGF-2−/- mice compared with wild-type littermates after ischemia. FGF-2 appears to specifically promote cell proliferation and neuronal differentiation in the ischemic dentate gyrus, as shown by BrdU labeling and analysis of colocalization of newly generated cells with the neuronal marker NeuN. Moreover, viral delivery of the FGF-2 gene to knockout animals on the day before ischemia increases neuronal proliferation to levels observed in wild-type littermates (Yoshimura et al, 2001). In addition to a role for FGF-2 in mediating the neurogenic response to injury in the adult dentate gyrus, the factor promotes neuronal replacement in typically nonneurogenic regions damaged by stroke. As discussed above, Nakatomi et al, (2002) found that ICV infusion of FGF-2 and EGF induces at least a fourfold increase in the number of newly generated CA1 pyramidal neurons present 1 month after ischemia. Recent studies further support a role for EGF in mediating strokeinduced neurogenesis and neuronal repair.
Intraventricular infusion of EGF beginning 2 days or several weeks after tMCAO in adult mice produces a dramatic increase in the number of newly generated cells in the ischemic striatum (Teramoto et al, 2003). Early administration of EGF appears to be important, since the proliferative response to late EGF treatment is less pronounced. Epidermal growth factor infusion early after ischemia also appears to enhance the migration of neuroblasts from the SVZ to the damaged striatum. At 13 weeks after ischemia, 65% of newly generated neurons express parvalbumin, suggesting that EGF facilitates replacement of interneurons in the ischemic striatum (Teramoto et al, 2003). Delivery of HB-EGF to ischemia-lesioned rats by adenoviral vector or ICV infusion also increases proliferation in the adult dentate gyrus (Jin et al, 2004b) and SVZ (Sugiura et al, 2005), reduces infarct volume (Jin et al, 2004b; also see Sugiura et al, 2005), elevates the number of new neurons in the ischemic striatum, improves functional outcome (Jin et al, 2004b; Sugiura et al, 2005), and promotes angiogenesis in the periinfarct striatum (Sugiura et al, 2005).
In adult rats subjected to middle cerebral artery embolic strokes, intravenous infusion of VEGF 48 h after stroke promotes microvascular formation in the ischemic border zone and improves functional neurological recovery compared with saline-treated rats (Zhang et al, 2000). In addition, ICV VEGF infusion for 3 days after tMCAO reduces infarct volume by ∼35% and improves neurological function compared with aCSF-infused ischemic rats (Sun et al, 2003). Vascular endothelial growth factor does not appear to enhance strokeinduced neurogenesis in the dentate gyrus or SVZ, but survival of adult-born cells is greatly increased in both neurogenic zones after VEGF treatment. Angiogenesis, measured as increased immunolabeling with endothelial cell markers, is also significantly upregulated in the ischemic striatum, but not in the dentate gyrus (Sun et al, 2003).
Neovascularization may provide a mechanism by which growth and neurotrophic factors are delivered to the area of ischemic damage, and might be important for the survival of new neurons generated in response to stroke. Mabuchi et al, (2005) evaluated whether neuronal proximity to microvasculature influences neuronal integrity after focal ischemia. In the ischemic striatum in adolescent baboons, uninjured neurons reside significantly closer to neighboring microvessels than do injured neurons, identified by dUTP incorporation. This is in agreement with the observation that newly generated cells in the postischemic monkey dentate gyrus are frequently in close proximity to proliferating blood vessels (Yamashima et al, 2004). In human stroke patients, increased blood vessel density in the penumbra correlates with longer survival after stroke (Krupinski et al, 1994).
Neurotransmitters and Other Factors
As discussed above, existing evidence suggests that glutamatergic mechanisms play a role in adult neurogenesis. Neurogenesis in the intact dentate gyrus decreases in response to activation of NMDA receptors, whereas administration of the NMDA receptor antagonist MK-801 increases DGC production (Cameron et al, 1995). In the ischemic brain, systemic and local hippocampal injections of MK-801 or the 2-(aminomethyl)phenylacetic acid/kainate (AMPA/KA) receptor antagonist 2,3-dihydroxy-6-nitro-7-sulfamoylbenzo(F)-quinoxaline (NBQX) prevents CA1 neuronal death and abolishes the strokeinduced increase in cell proliferation in the dentate gyrus after transient global ischemia in adult gerbils (Bernabeu and Sharp, 2000). Blocking NMDA receptor activation via MK-801 also prevents the ischemia-induced increase in proliferation in the adult rat dentate gyrus after tMCAO, whereas NBQX treatment has no effect in this model (Arvidsson et al, 2001). Glutamatergic mechanisms mediating strokeinduced neurogenesis in the adult dentate gyrus thus appear to be unique in that NMDA receptors modulate the effect of focal ischemia, whereas both NMDA and AMPA receptors are involved in the neurogenic response to global ischemia. Alternatively, species-specific differences might be more important than the type of ischemic insult. Also of interest is the finding that NMDA receptor blockade enhances neurogenesis in the intact dentate gyrus (Cameron et al, 1995; Bernabeu and Sharp, 2000), but suppresses it in the ischemic brain (Bernabeu and Sharp, 2000; Arvidsson et al, 2001). It seems likely that other glutamate-mediated changes in the ischemic brain, such as increased synthesis of specific growth factors, may override the normal action of glutamatergic mechanisms on adult neurogenesis (Arvidsson et al, 2001).
Additional studies support a role for nitric oxide in strokeinduced neurogenesis. Intravenous delivery of a nitric oxide donor to ischemic rats increases cell proliferation in the dentate gyrus and SVZ and improves performance on the rotarod task (Zhang et al, 2001a). Members of the nitric oxide synthase (NOS) family may differentially influence the response of endogenous progenitor cells to ischemia and other brain insults. Whereas strokeinduced proliferative activity in the dentate gyrus is abolished in null mutant mice lacking inducible or endothelial NOS (Zhu et al, 2003; Chen et al, 2005), neuronal proliferation in the dentate gyrus, SVZ, and striatum ipsilateral to ischemic injury increases in mice deficient in neuronal NOS (Sun et al, 2005). These findings suggest that neuronal NOS negatively regulates strokeinduced neurogenesis, but that inducible and endothelial NOS serve as positive modulators of neurogenesis in the ischemic brain. Interestingly, mutant mice lacking endothelial NOS display poorer functional outcome after stroke than do wild-type littermates, as well as reduced angiogenesis in the ischemic border and an apparently selective decrease in BDNF expression (Chen et al, 2005).
The cytokine erythropoietin (EPO) is also a potential modulator of ischemia-induced neurogenesis. Infusion of EPO directly into the brains of adult mice increases both cell proliferation in the SVZ—RMS pathway and the number of new interneurons added to the olfactory bulb (Shingo et al, 2001). EPO receptors are expressed in the adult SVZ and the cytokine is upregulated in response to mild hypoxia (Shingo et al, 2001), making it a good candidate for mediating injury-induced surges in neurogenesis. Wang et al, (2004) treated adult rats systemically with EPO 24 h after focal ischemia induction by MCA embolus. They found that EPO increases SVZ neurogenesis, periinfarct angiogenesis and stroke recovery. These changes occur in association with increased brain VEGF and BDNF levels. Thus, EPO may exert its effects on strokeinduced angiogenesis and neurogenesis indirectly through other factors.
Retinoic acid (RA) is a vitamin A derivative that has pleiotropic effects on neural development. Recently, we found that retinoid signaling augments SVZ neurogenesis and is required for SVZ neuroblast migration to the olfactory bulb in neonatal mice (Wang et al, in press). To determine whether RA influences SVZ neurogenesis after stroke, adult rats were administered intraperitoneal RA every other day for several weeks after tMCAO, and were killed on day 35 after stroke. As shown in Figures 4C and 4D, RA treatment markedly increased the numbers of doublecortin-expressing cells in the ipsilesional SVZ. Studies are ongoing to determine whether this augmentation of neurogenesis leads to increased numbers of striatal neurons and improved functional outcome after focal ischemia.
Relevance to Stroke in Humans
Cell proliferation in the dentate gyrus and SVZ of young adult macaque or cynomolgus monkeys increases in response to global or focal ischemia, respectively (Tonchev et al, 2003). The presence of strokeinduced neurogenesis in these primate models suggests that mobilization of endogenous progenitors is a reasonable therapeutic avenue to pursue in human stroke patients. Many parameters, however, are more amenable to treatment in experimental animals than in patients. For example, at least one study in rodents suggests that early intervention to augment neurogenesis after ischemia is critical (e.g., Teramoto et al, 2003), and some stroke patients might not receive treatment within the temporal window of efficacy. Most experimental studies are conducted in young adult animals, but stroke is most prevalent among the aged population; repairing the aged ischemic brain might be significantly more challenging given age-related declines in neurogenesis (reviewed in Bernal and Peterson, 2004).
Regenerative Strategies for Stroke
Recent findings in rodent experimental stroke models suggesting striatal and hippocampal pyramidal cell replacement by endogenous progenitors raise optimism for regenerative therapies. Early work using primate models of stroke seems to support this optimism. Importantly, neuronal replacement strategies that rely on endogenous progenitors avoid many of the potential technical and ethical limitations associated with fetal or stem cell transplantation. Many issues, however, remain to be addressed to allow for the adequate assessment of the feasibility of this therapeutic approach. Foremost is the fact that the potential for harnessing endogenous neural progenitors in the human brain for repair after stroke or other injuries remains unknown. Goldman and colleagues (Roy et al, 2000a, b ) have devised methods to purify neural progenitors from adult human brain; however, they estimate that the total numbers of endogenous forebrain neural progenitors are very small, such that their manipulation alone might not be satisfactory for repair. Additional cautions arise from rodent data, suggesting that adult neurogenesis is maladaptive after certain forms of injury (Parent et al, 1997; Scharfman et al, 2000) and that, unlike in the neostriatum, the adult rodent cortex appears to be hostile to neuronal replacement (Arvidsson et al, 2002; Parent et al, 2002b). Nonetheless, the important findings that newly generated neurons in the adult neocortex can replace dying neurons and form long-distance connections (Magavi et al, 2000; Chen et al, 2004) are proof of principal that endogenous repair is possible.
Future steps involve determining whether newly generated neurons functionally integrate after stroke and other forms of brain injury, and whether this neuronal replacement leads to functional recovery. Developing methods to expand the endogenous progenitor pool, direct migration and differentiation, and promote survival of the progeny is a priority, and these tasks will benefit from ongoing advances in stem cell biology. The application of findings from rodent models to primate models of stroke and other brain injuries, and to human stem cells in culture, also are necessary steps for developing regenerative therapies. Knowledge of the role of neural stem cells in brain development and repair after injury is advancing rapidly indeed, and raises excitement for the prospect of stem cell regenerative therapies for stroke and other brain disorders.