
Abstract
The right hemisphere is predominantly involved in tasks associated with spatial attention. However, left hemispheric dominance for spatial attention can be found in healthy individuals, and both spatial attention and language can be lateralized to the same hemisphere. Little is known about the underlying regional distribution of neural activation in these ‘atypical’ individuals. Previously a large number of healthy subjects were screened for hemispheric dominance of visuospatial attention and language, using functional Doppler ultrasonography. From this group, subjects were chosen who were ‘atypical’ for hemispheric dominance of visuospatial attention and language, and their pattern of brain activation was studied with functional magnetic resonance imaging during a task probing spatial attention. Right-handed subjects with the ‘typical’ pattern of brain organization served as control subjects. It was found that subjects with an inverted lateralization of language and spatial attention (language right, attention left) recruited left-hemispheric areas in the attention task, homotopic to those recruited by control subjects in the right hemisphere. Subjects with lateralization of both language and attention to the right hemisphere activated an attentional network in the right hemisphere that was comparable to control subjects. The present findings suggest that not the hemispheric side, but the intrahemispheric pattern of activation is the distinct feature for the neural processes underlying language and attention.
Introduction
Functional imaging has increased awareness for the variability in the relation between brain function and brain structure. This variability is most striking for lateralized brain functions. Neuroimaging and neuropsychological research indicate that the left hemisphere is specialized for language and the right for control of spatial attention in most individuals (Ojemann, 1991; Mesulam, 1999; Bookheimer, 2002). However, this is not an invariable principle. There are occasional patients with aphasias after right-hemispheric lesions, or with neglect after left-hemispheric lesions (Dronkers and Knight, 1989). The same hemisphere can even be dominant in control of both language and spatial attention (Fischer et al, 1991; Trojano et al, 1994; Alexander and Annett, 1996; Osmon et al, 1998; Floel et al, 2001; Floel and Knecht, 2002).
Recently, a number of studies have addressed the anatomical correlates of language processing in subjects with atypical right-hemisphere language dominance (Hubrich-Ungureanu et al, 2002; Staudt et al, 2002; Knecht et al, 2003; Jansen et al, 2004b). They showed that right-hemisphere recruitment for language occurs in brain areas homotopic to the left-hemisphere regions typically involved in language processing. Detailed studies on the lateralization of the neural substrate that underlies attentional processing in atypical cases are missing so far.
In a previous study on healthy subjects, we investigated whether functional transcranial Doppler sonography (fTCD) and functional magnetic resonance imaging (fMRI) determine hemispheric dominance of brain activation related to visuospatial attention concordantly (Jansen et al, 2004a). In this study, we identified three subjects who were left-hemisphere lateralized for spatial attention and right-hemisphere dominant for language and two subjects who were right-hemisphere dominant for both language and spatial attention.
This unique sample of subjects gave us the possibility to investigate two important questions: (i) the neuroanatomic correlates of visuospatial attention processing in the left hemisphere, and (ii) the influence of atypical right-hemispheric language lateralization on right-hemispheric visuospatial attention. In the present study, we therefore analyzed the data for topographical information about brain activation, which were not included in the previous study.
We hypothesized that atypical left-hemispheric dominance for attention would likewise be a mirror of right-hemispheric attentional dominance, similar to what has been shown for language (Knecht et al, 2003). For subjects with both language and attention lateralized to one hemisphere, we hypothesized that the intrahemispheric distribution of activation would not differ from subjects with the typical pattern. Seven subjects from the previous study, with the typical pattern of handedness (right), language (left), and attention (right) served as control group.
Materials and methods
Subjects
Twelve healthy volunteers (5 men), aged 24 to 30 years, were included in the study. Based on their language and attentional dominance, subjects were selected from a cohort of 15 subjects previously assessed for attentional and language dominance with both fMRI and fTCD to examine the intermethod-correlation of both techniques (Jansen et al, 2004a). Subjects in the study by Jansen and colleagues had been selected from a cohort of 326 healthy volunteers studied for language dominance by fTCD (Knecht et al, 2000a, b). A much higher percentage of the subjects chosen for the Jansen et al study, compared with the general population, had atypical right-hemispheric representation for language to increase the possibility to include subjects with an atypical left-hemispheric representation for spatial attention. In the present study, we chose five subjects with atypical hemispheric dominance for language and/or attention from the previous study (Jansen et al, 2004a) for in-depth examination of the intrahemispheric topography of atypical hemispheric lateralization. All subjects had completed the equivalent of a high school degree (‘Gymnasium'). None had a serious history of medical, neurologic, or psychiatric illnesses or any brain pathology or abnormal brain morphology on T1-weighted MR images. All subjects gave their written informed consent before participation, according to the declaration of Helsinki. The subjects' characteristics are presented in Table 1.
Subjects' characteristics: age, sex, handedness, and language lateralization
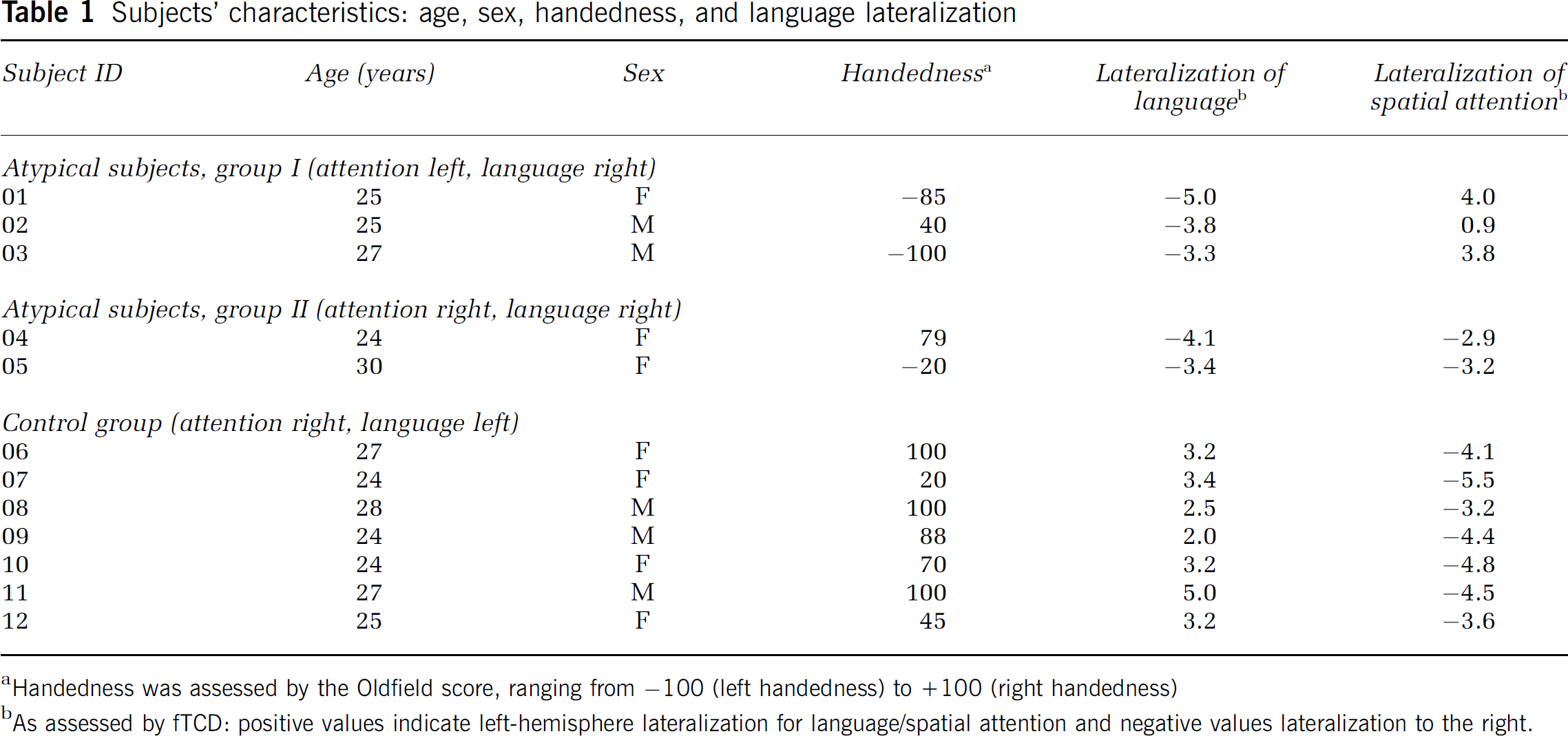
Handedness was assessed by the Oldfield score, ranging from −100 (left handedness) to +100 (right handedness)
As assessed by fTCD: positive values indicate left-hemisphere lateralization for language/spatial attention and negative values lateralization to the right.
Handedness was assessed by the Edinburgh handedness inventory (Oldfield, 1971). Hemispheric dominance for language and spatial attention was originally assessed by fTCD (see below), and confirmed by fMRI (Knecht et al, 2003; Jansen et al, 2004a).
Functional Transcranial Doppler Ultrasonography
Experimental Design: Word generation (WG). Language lateralization was assessed with a standard phonemic verbal fluency task, which is based on the Controlled Word Association Test (Lezak, 1995). Subjects were presented with a letter on a computer screen 5 secs after a cueing tone. They silently generated as many words as possible starting with the displayed letter. Task compliance was ensured by having subjects report the words after a second auditory signal, 15 secs after presentation of the letter. A third auditory signal, 5 secs later, marked the beginning of a control condition that lasted for 35 secs, and during which subjects were instructed to imagine a night sky (control condition). The following trials were presented after the same schema. One epoch lasted 60 secs (25 secs WG, 35 secs control task). Twenty epochs were recorded in total. Letters were presented in random order. ‘Q', ‘X', and ‘Y’ were excluded because few words in German start with these letters. Each letter was only displayed once.
Line bisection. To assess spatial attention, the Landmark Task, a variant of line bisection, which is frequently used in the assessment of visuospatial neglect (Harvey et al, 1995), was administered. Five seconds (secs) after a cueing tone, a horizontal line was presented for 10 secs on a computer screen (visual angle 9.3°). The horizontal line was bisected by a vertical line either in the exact middle or deviating to the right or the left by 0.1° or 0.2°. During the 10-sec period, subjects had to silently decide if the horizontal line was bisected in the middle, slightly to the left, or slightly to the right. Task compliance was ensured by having subjects report the decision verbally (middle, left, or right) after a second auditory signal at the end of the presentation of the bisected line. A third auditory signal 5 secs later marked the beginning of the control condition. The next bisected line was then presented in the same way. One epoch lasted 40 secs (20 secs for Landmark Task, 20 secs for control condition). Twenty epochs were recorded in total. Lines were bisected in the exact middle four times, and four times each displaced to the left or to the right by 0.1° or 0.2°, respectively. The sequence of conditions was randomized.
Functional transcranial Doppler sonography data acquisition: Using a commercially available 2 MHz transcranial Doppler ultrasonography device (Multidop T, DWL Sipplingen, Germany), the blood flow velocities within both middle cerebral arteries (MCAs) were measured continuously. Details of the technique, especially the identification of the MCAs, are described elsewhere (Ringelstein et al, 1990). The spectral envelope curves of the Doppler signal were stored for offline analysis.
Functional transcranial Doppler sonography data analysis: Analysis was performed using the software package AVERAGE (Deppe et al, 1997). After automatic artefact rejection, like probe displacement, data were integrated over the corresponding cardiac cycles, segmented into epochs that related to the first cueing tone and averaged. The mean velocity in the 15-sec precueing interval (Vpre.mean) was taken as a baseline value. The relative CBFV changes (dV) were calculated by the formula
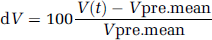
where V(t) is the CBFV over time and Vpre.mean is the mean velocity in the precueing interval.
Examples of dV change related to visuospatial attention are shown in Figure 3. Differences in the velocity between both MCAs were statistically analyzed using the Wilcoxon test for each sample point.
Calculation of LIfTCD
The LIfTCD was computed by
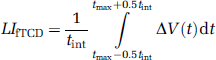
where
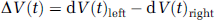
is the difference between the relative CBFV changes in the left and right MCA. The time point tmax represents the latency of the absolute maximum of ΔV(t) within the activation interval (10 to 20 secs). As integration interval, a time period of tint = 2 secs was chosen. A positive value of the LIfTCD indicates left-hemispheric processing for spatial attention; a negative value stands for right-hemispheric dominance. The magnitude represents the degree of lateralization. The accuracy of the LIfTCD of each individual is determined by its stochastically estimated confidence interval.
According to their hemispheric dominance for language and spatial attention, subjects were divided into three groups:
Group I (subjects 01–03): Right hemispheric dominance for language, left hemispheric dominance for spatial attention.
Group II (subjects 04–05): Right hemispheric dominance for both language and spatial attention.
Group III (subjects 06–12): Left hemispheric dominance for language, right hemispheric dominance for spatial attention.
Group I thus comprised ‘atypical’ subjects with dissociated lateralization of language and attention, and Group II ‘atypical’ subjects with associated lateralization of language and attention. Group III (subjects with the standard or typical pattern of hemispheric lateralization) served as a control.
A missing dissociation of lateralization of attention and language is often associated with cognitive deficits. Therefore, subjects who lacked dissociation between lateralization for language and for visuospatial attention were tested post hoc with the Leistungspruefsystem (LPS) for adults <50 years (Sturm et al, 1993) to assess for verbal and nonverbal intelligence. To screen for attentional deficits, digit span was assessed using the subtest of the Wechsler Memory Scale (WMS) (Wechsler and Stone, 1974), and visual span using the Corsi-tapping test (Lezak, 1995). Speed of processing, mental flexibility, and attention were examined with the Trail Making Test (Spreen and Strauss, 1991).
Magnetic Resonance Imaging Experimental Design
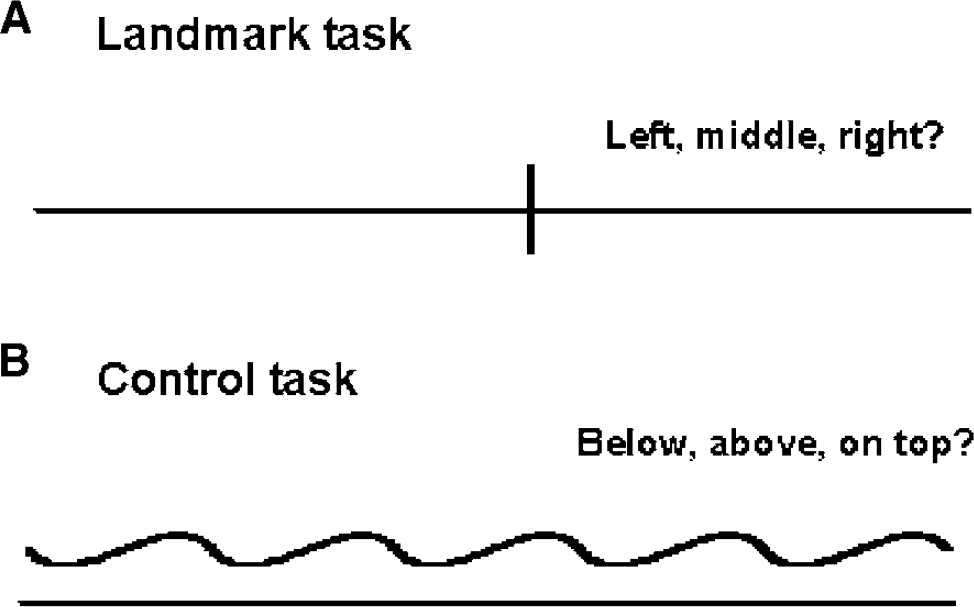
For the purpose of functional imaging studies, hemispheric lateralization for attention may be assessed in a number of ways (for a review see Cabeza and Nyberg, 2000). In the present study, we chose to examine lateralization for spatial attention with the Landmark task. Subjects had to decide whether prebisected horizontal lines are divided in the exact center, or slightly to the right or left of the center (Figure 1). This task was chosen, although it is not without criticism (Halligan et al, 1989; Ferber and Karnath, 2001), because it corresponds to deficits frequently encountered in patients experiencing visuospatial neglect after stroke (Harvey et al, 1995; Heilman, 1997; Marshall, 1998; Mesulam, 1999).
(
The control task used in the present study was devised to incorporate all aspects of the experimental task except the bisection judgment itself (Fink et al, 2000, 2002). It controls for the visual input, the neural responses resulting from the hand used for the responses that indicate subjects' performance, and the decision component involved in the task per se. In this way, we depicted the neural substrates that are specific to the visuo-spatial judgments associated with the experimental task.
Five seconds after a color change (duration 0.5 secs), subjects were presented with a horizontal line for 5 secs on a computer screen (visual angle 9.3°). The horizontal line was either bisected by a vertical line in the exact middle or slightly deviating to the left or the right (Landmark task) or presented together with a waveform-shaped line (control task). During the 5-sec period, subjects silently decided where the horizontal line was bisected (Landmark task), or where the waveform-shaped line was presented (control task), respectively. Immediately after the line presentation, subjects were instructed to report their decision by pressing corresponding buttons on a response pad with both hands. After 2 secs, another color change marked the beginning of the next epoch. Six Landmark tasks were followed by six control tasks. This sequence was repeated seven times.
Magnetic Resonance Imaging Data Acquisition
All MRI data were acquired on a neuro-optimized GE 1.5T whole body scanner equipped with a standard circular polarized head coil. Functional images were acquired using a T*2-weighted gradient echo EPI sequence (TE = 40 ms, TR = 2 secs, flip angle 90°, slice thickness 7 mm, 1 mm gap, matrix 64 × 64, FOV 200 mm, in-plane resolution 3.125 × 3.125 mm). Sixteen axial slices orientated parallel to the AC-PC line covering the whole head were taken. Additional high-resolution T1-weighted anatomic images were acquired (TE = 8 ms, TR = 24 ms, flip angle 30°, matrix 256 × 256, FOV 250 mm, 124 sagittal slices, slice thickness 1.5 mm).
Magnetic Resonance Imaging Data Analysis
All calculations and image manipulations were performed on an Enterprise 450 (SUN Microsystems Computers) using MATLAB (www.mathworks.com) and SPM99 (www.fil.ion.ucl.ac.uk/spm). Statistical parametric map (SPM) was used for realignment, normalization, smoothing, and statistical analysis to create statistical parametric maps of significant relative regional BOLD response changes (Friston et al, 1995a, b ).
The first five images of each session, during which the MR signal reaches a steady state, were discarded. The remaining scans of each individual were realigned to the first image (corresponding to the sixth acquired image of the time series) to correct for movement artefacts. Sinc interpolation was used in the transformation. A mean functional image volume was constructed for each subject from the realigned images and coregistered with the anatomic image. The functional images were normalized to a template brain image created by the Montreal Neurological Institute (MNI) (Evans et al, 1994). The coregistered anatomic image was used to determine the parameters for the spatial normalization process (Friston et al, 1995a). The resulting voxel size in standard stereotactic coordinates was 2 × 2 × 2 mm3. The normalized images were smoothed using an isotropic Gaussian kernel (FWHM: 6 mm for single subject analysis, 10 mm for group analysis).
After preprocessing, statistical analysis was performed on individual data. The experimental conditions (judgment of prebisected lines/control task) were modelled using a boxcar function convolved with a hemodynamic response function (hrf) in the context of the general linear model used by SPM99. The time series was filtered with the hrf as low-pass filter. Specific effects were examined by applying appropriate linear contrasts to the parameter estimates for the experimental conditions, resulting in a t-statistic for every voxel. These t-statistics constitute an SPM and can be interpreted by referring to the probabilistic behavior of Gaussian random fields.
To obtain activation maps across subjects, the functional data within the control group were combined in a ‘fixedeffect’ analysis as the most sensitive approach involved to identify areas in attentional processing (Friston et al, 1999). For the ‘atypical’ subjects, we did not perform a group analysis. On the one hand, the group sizes were too small; on the other hand, one could not expect a priori that the ‘atypical’ subjects show the same kind of ‘atypicality', that is, activate the same brain regions among each other. Their respective individual activation pattern was compared with the typically activated brain region of the control group.
The anatomical localizations of activated brain regions were determined by reference to a standard stereotactic atlas (Talairach and Tournoux, 1988), using the T2T applet (http://neurologie.uni-muenster.de/ger/dienstleistungen/index.html) (Deppe et al, 2003).
Results
Behavioral Testing
Subjects with associated lateralization for language and attention (subjects 4 and 5), often associated with cognitive deficits, showed normal or above normal scores on standard psychological testing (Table 2). Notably, performance on nonverbal tasks was not below overall performance.
Neuropsychologic examination of the two subjects (subjects 4 and 5) with lateralization of both language and spatial attention to the right hemisphere
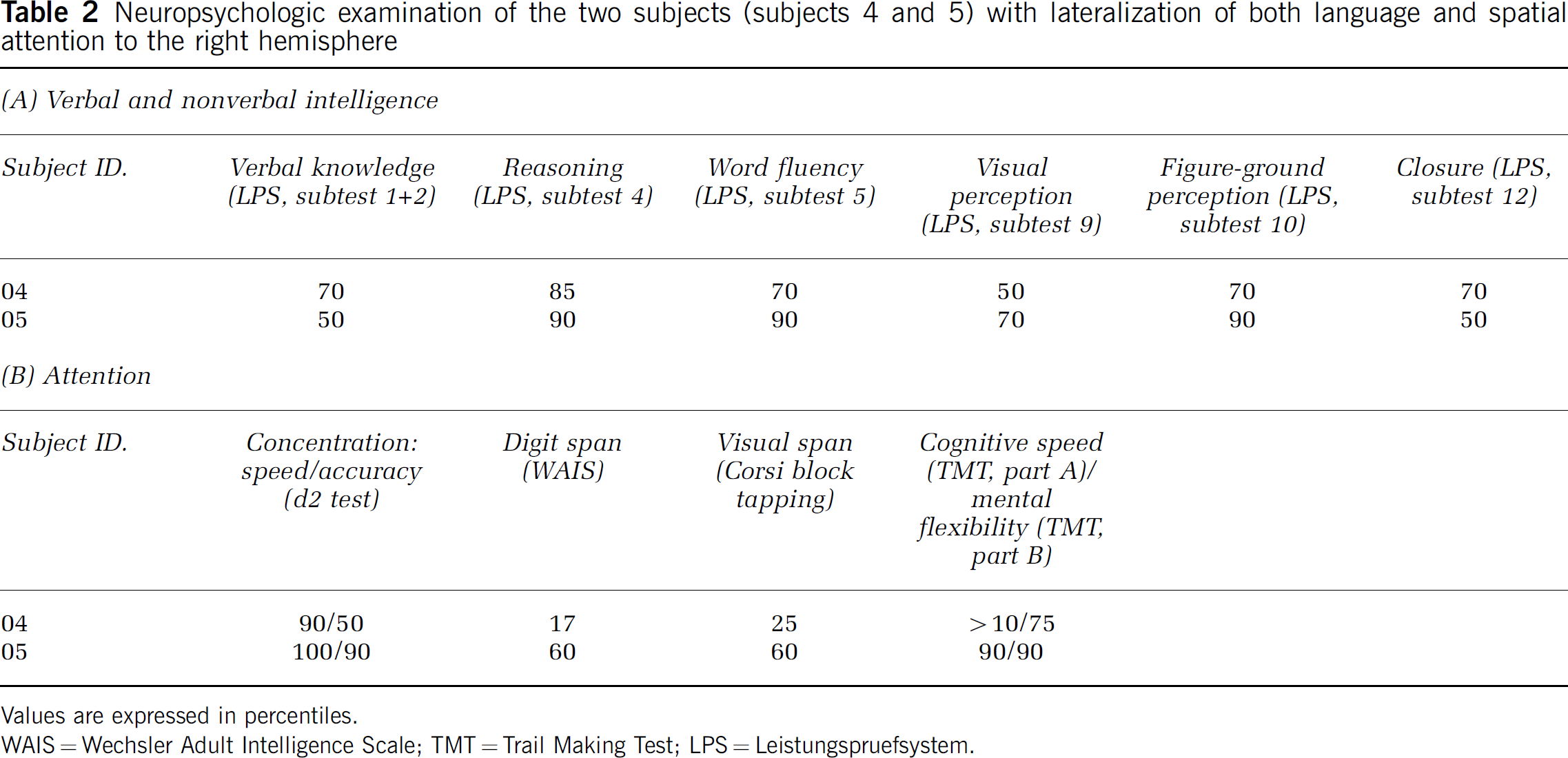
Values are expressed in percentiles.
WAIS = Wechsler Adult Intelligence Scale; TMT = Trail Making Test; LPS = Leistungspruefsystem.
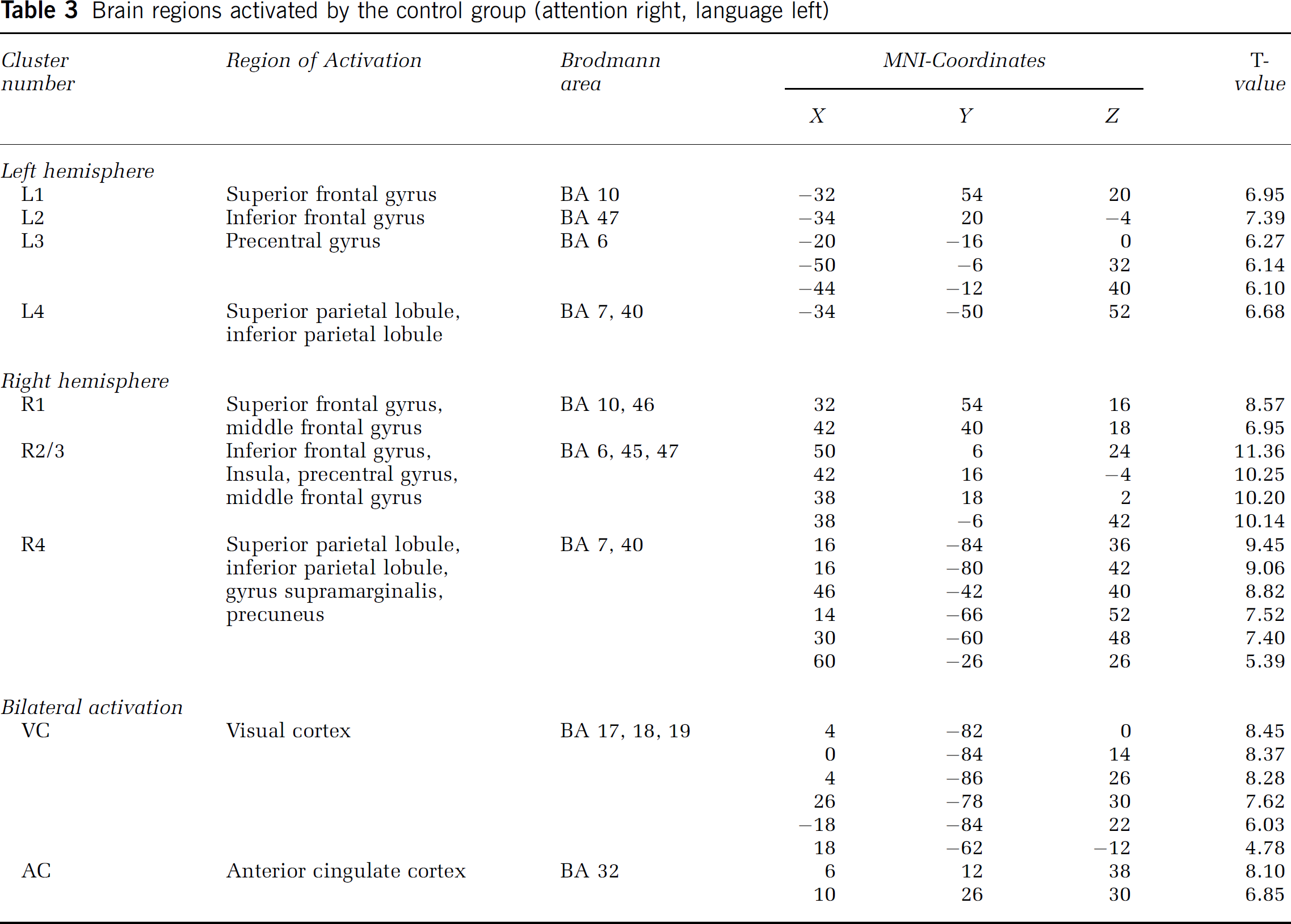
Brain Regions Activated by the Control Group (Right-Hemisphere Dominance for Spatial Attention, Left-Hemisphere Dominance for Language)
Figure 2 and Table 3 summarizes for the control group the regions with increased neural activity associated with the Landmark task (P<0.01, corrected for multiple comparisons). Only clusters with a size of at least 100 activated voxels are displayed. Additionally, to illustrate the differences between the individual activation and the group activation, the activated pattern of a representative subject is provided in Figure 3A.
Brain regions activated by the control group (attention right, language left)
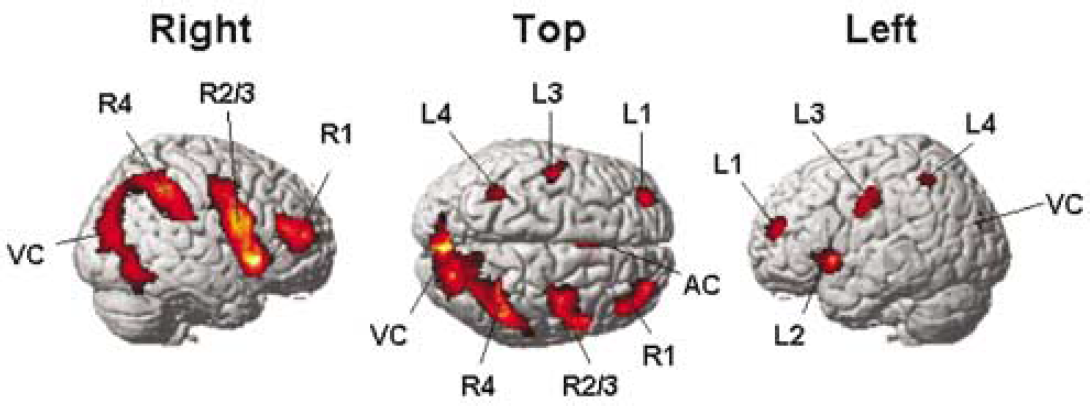
Activation in the control group. Relative increases in neural activity associated with the landmark task relative to the control task (P<0.01 corrected for multiple comparisons).
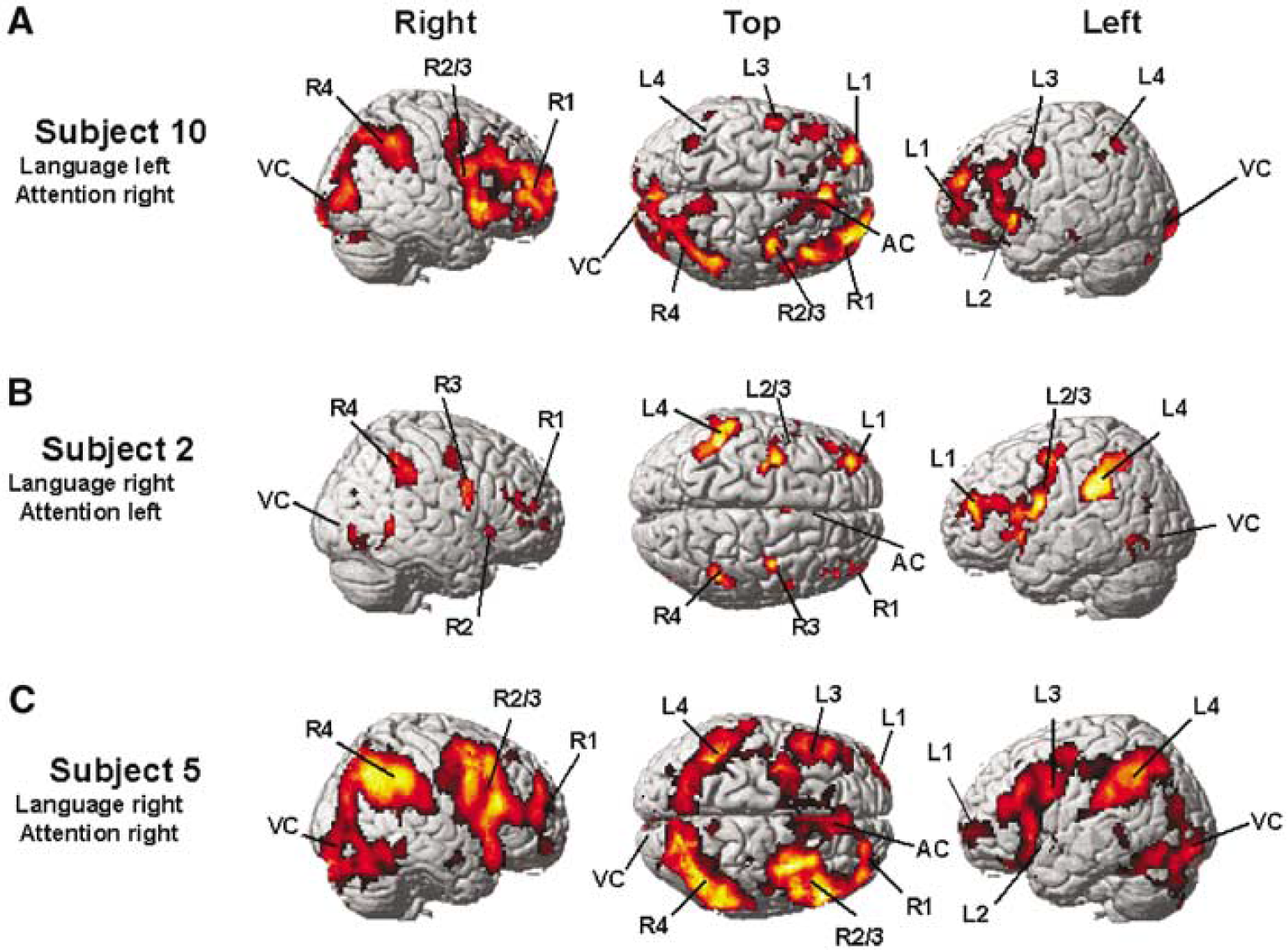
Landmark activation pattern of representative subjects (P<0.001, uncorrected for multiple comparisons) with (
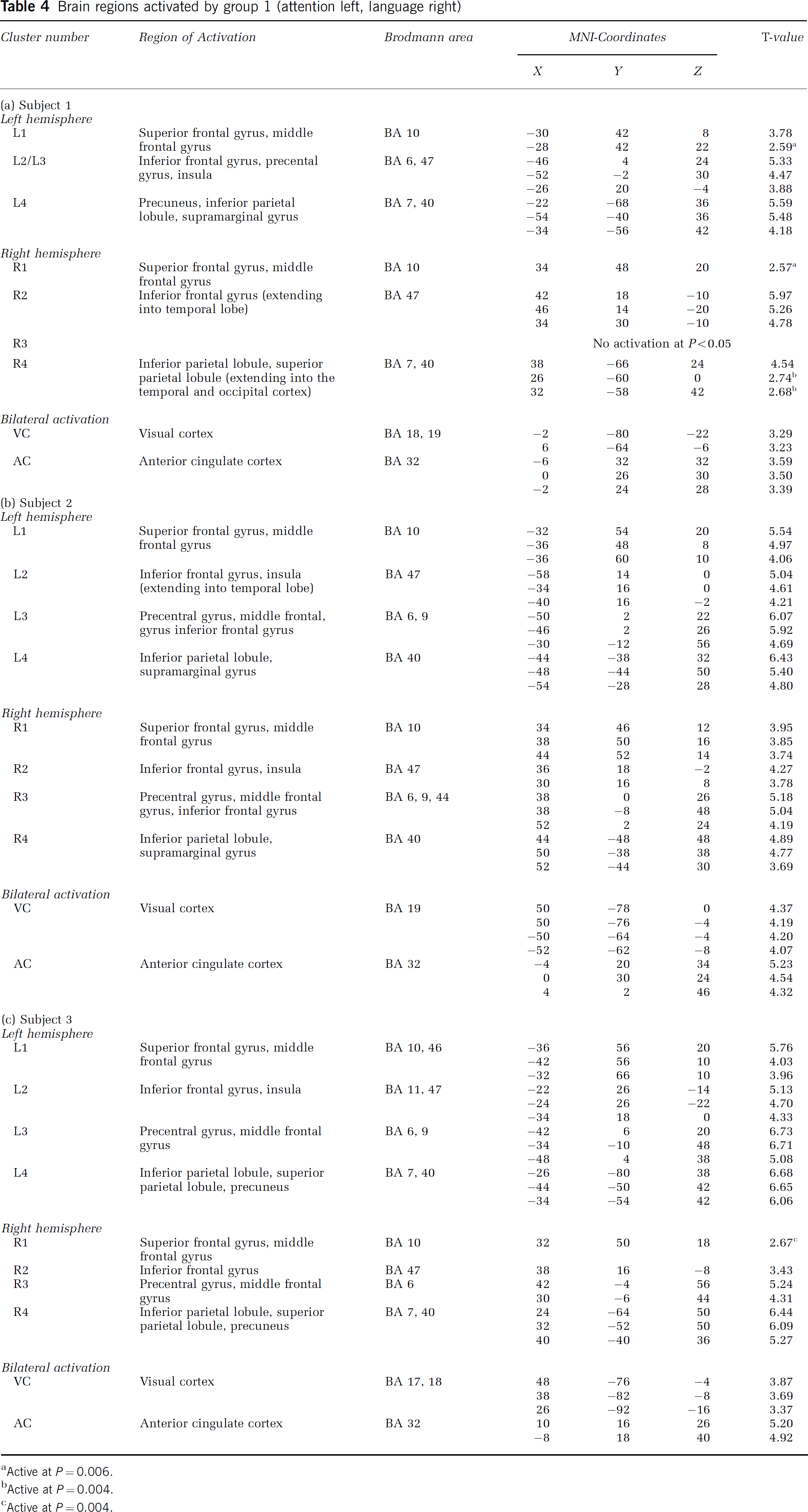
Brain regions activated by group 1 (attention left, language right)
Active at P = 0.006.
Active at P = 0.004.
Active at P = 0.004.
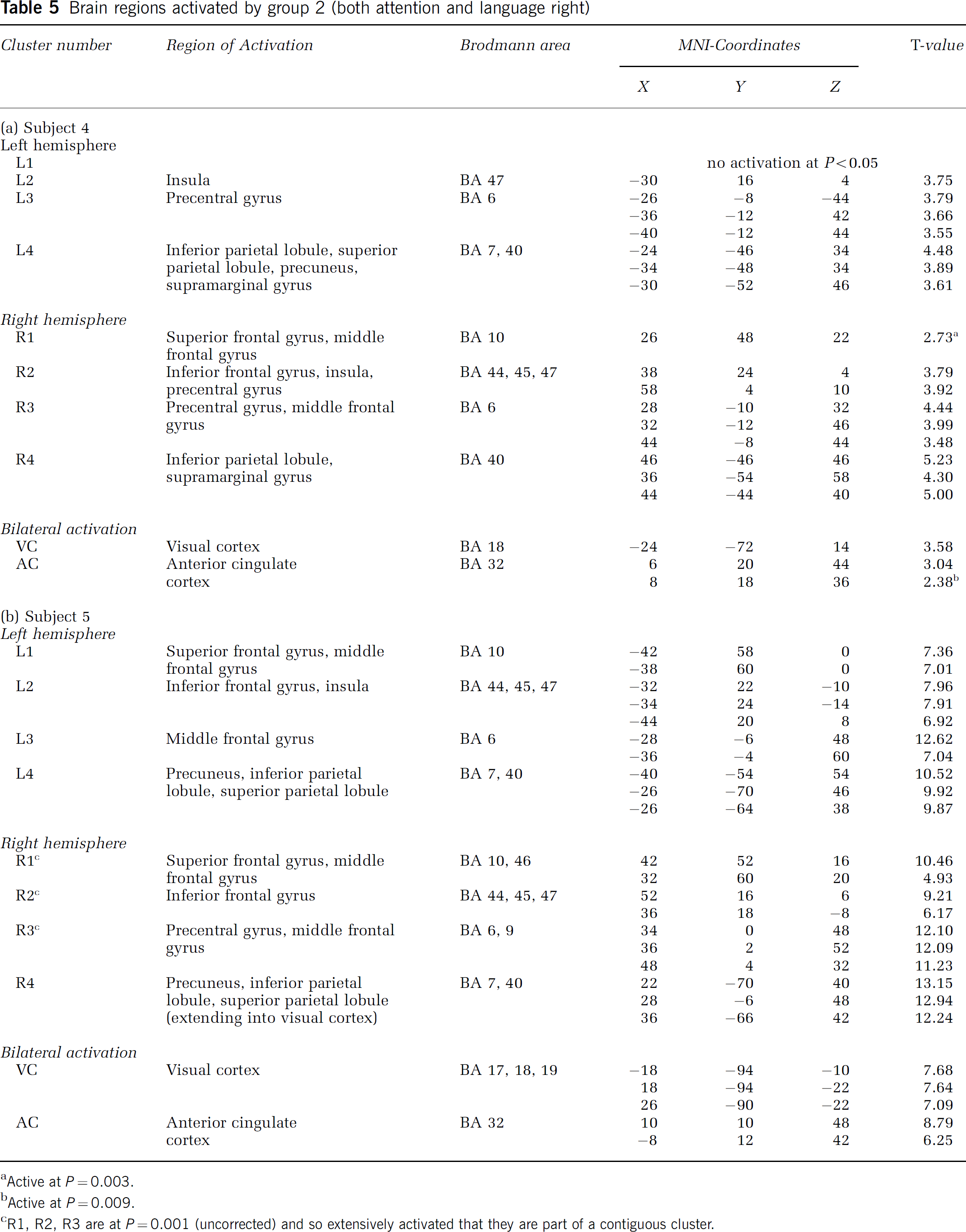
Brain regions activated by group 2 (both attention and language right)
Active at P = 0.003.
Active at P = 0.009.
R1, R2, R3 are at P = 0.001 (uncorrected) and so extensively activated that they are part of a contiguous cluster.
Increases in neural activity were observed in the ventrolateral prefrontal cortex (R1 and L1, resp.), the lateral orbito-frontal cortex (R2 and L2, resp.), the dorsolateral frontal cortex (R3 and L3, resp.), and the lateral parietal cortex (R4 and L4, resp.). These activations were observed in the right as well as in the left hemispheres, although the extent of activation was higher in the right hemisphere. Additionally, there was bilateral activation in the anterior cingulate cortex (AC) and the visual cortex (VC).
Brain Regions Activated by Atypical Subjects from Group I (Left-Hemisphere Dominance for Spatial Attention, Right-Hemisphere Dominance for Language)
Table 4 summarizes the brain regions that were activated by the subjects 1–3 during the Landmark task (P<0.001, uncorrected). Figure 3B provides the activation pattern of a representative subject. All three subjects activated brain regions homotopic to those activated by the control group.
Brain Regions Activated by Atypical Subjects from Group II (Right-Hemisphere Dominance for Both Spatial Attention and Language)
Table 5 summarizes the brain regions that were activated by the subjects 4 and 5 during the visuospatial task (P<0.001 uncorrected). Figure 3C provides the activation pattern of a representative subject. Both subjects activated the same brain regions as the control group. There was no influence of the atypical right-hemisphere language lateralization on the
Discussion
To the best of our knowledge, this is the first study that examines the functional neuroanatomic correlates of atypical left-hemispheric attentional processing, and the influence of atypical right-hemisphere language dominance on the functional organization of right-hemispheric visuospatial attention.
Our data showed that
in subjects with atypical left-hemisphere dominance for spatial attention, recruitment of brain areas occurs in regions homotopic to the regions typically activated by subjects with ‘normal’ right-hemisphere dominance;
for subjects with both language and spatial attention lateralized to one hemisphere, the intrahemispheric activation pattern during a visual-spatial attention task does not differ from subjects with the typical pattern of hemispheric dominance.
Intrahemispheric Pattern of Activation for Spatial Attention Tasks
The pattern of activation in our control group (of ‘typical’ subjects) is in line with findings of a previous study by Fink and colleagues who used a similar paradigm (Landmark task) in healthy volunteers (Fink et al, 2000, 2001). During the Landmark task, these authors found activation predominantly in the right superior and inferior parietal cortex, as well as prefrontal cortex and anterior cingulate. The intrahemispheric pattern of activation in their study was also similar to the activations found in our subjects with left-hemispheric attentional dominance.
The present study concurs with findings on atypical language lateralization (Knecht et al, 2003) by showing that distinct intrahemispheric brain regions are activated in spatial attentional tasks, regardless of hemispheric dominance and regardless of language lateralization. Taken together, the two studies strongly support the notion that the intrahemispheric pattern and not the interhemispheric side is the distinct feature for the neural processes underlying language and spatial attention. In the following sections, we will discuss the implications of these findings from a theoretical and clinical perspective.
Theories on Cerebral Lateralization
A major phenomenon of human brain function is its ubiquitous lateral cerebral specialization (Geschwind and Galaburda, 1985; Gazzaniga, 2000). The key features of this specialization have traditionally been the left hemisphere's superiority for language and speech and the right hemisphere's superiority for tasks such as attentional monitoring. This division was thought to result from the fact that the left hemisphere has marked limitations in perceptual functions, and the right hemisphere prominent limitations in its cognitive functions (Gazzaniga, 2000). Other researchers have suggested that lateralization of language and control of spatial attention dissociate because the cerebral hemispheres differ fundamentally in their computational modes, with the left being biased for symbolic and the right for analogue processes (Corballis, 1997). Relative hemispheric specialization for global and local processing has also been proposed to account for lateralization of cognitive functions (Robertson and Lamb, 1991; Fink et al, 1997; Barrett et al, 1998).
A growing body of lesion studies (Fischer et al, 1991; Trojano et al, 1994; Alexander and Annett, 1996; Osmon et al, 1998) and activation studies (Floel et al, 2001, 2003; Floel and Knecht, 2002; Hubrich-Ungureanu et al, 2002; Staudt et al, 2002; Knecht et al, 2003) inform us, however, that other patterns of hemispheric spezialization may occur. For language, a number of studies have addressed the funtional topography in subjects with atypical right-hemisphere language dominance (Hubrich-Ungureanu et al, 2002; Staudt et al, 2002; Knecht et al, 2003; Jansen et al, 2004b). The underlying neuroanatomic correlates for attention are largely unknown, however. In the present study, we have addressed this issue, and found that also for spatial attention, atypical hemispheric dominance shows a mirror-reverse pattern of brain activation, compared with the ‘standard’ pattern.
These findings suggest that atypically lateralized brain functions in general are localized to homotopic sites within the respective hemisphere. Intrahemispheric pattern rather than hemispheric side seems to be a hard-wired pattern of brain organization, at least in healthy individuals (see examples of cross-modal plasticity in chronic deafferentation, review in Burton, 2003).
Neuropsychologic Implications
Our data further suggest that the brain may organize in a way that language and spatial attention can exist within the same hemisphere without penalty to brain function. The subjects in our study with lateralization of both language and attention to the right hemisphere displayed normal intellectual, linguistic, and spatial performance. Although we did not perform detailed morphometric MRI analysis, we inspected the MRIs for evidence of structural damage. No such changes were detected. Therefore, brain lesions early in life, which might have been involved in the development of atypical lateralization for language and attention, are highly unlikely in our case.
On the basis of studies in patients with early-onset seizures (Saykin et al, 1989; Strauss et al, 1990; Helmstaedter et al, 1994), it has been hypothesized that the limited computational capacities would not allow one hemisphere to be dominant for both language and spatial attention without detrimental effects to linguistic and spatial abilities (‘crowding hypothesis') (Saykin et al, 1989; Strauss et al, 1990; Carlsson et al, 1994; Helmstaedter et al, 1994; Mariotti et al, 1998; Staudt et al, 2002). However, in contrast to previous studies on the crowding phenomenon (Saykin et al, 1989; Strauss et al, 1990; Carlsson et al, 1994; Helmstaedter et al, 1994; Mariotti et al, 1998; Staudt et al, 2002), no negative behavioral consequences could be ascertained in our subjects. Most importantly, we did not find major discrepancies between verbal and nonverbal performance. We believe that the decisive difference to the previous studies, which all examined populations with left-hemispheric damage or seizures, is that our subjects do not have any evidence of (left-hemispheric) damage.
Clinical Relevance
Our findings show that all patterns of cerebral lateralization may occur in healthy subjects, and that in certain individuals, different functions may lateralize independently from each other. Clinicians need to be aware of the possibility of combined linguistic and visuospatial impairments of variable degrees after left- or right-hemispheric damage.
Our results are also of direct clinical relevance in patients where fMRI is used to determine cortical ‘attention areas' before brain surgery (e.g., tumor or epilepsy surgery). Perhaps the most difficult task in the interpretation of fMRI data is the differentiation between ‘artificial’ and ‘real’ activation. From our results, it can be concluded that in patients with atypical hemispheric dominance of a cognitive function, activation should be expected in homotopic brain regions. Activation outside these expected locations should be interpreted with caution.
Conclusion
In the present study, we showed that not hemispheric side, but the intrahemispheric pattern, is the distinct feature of localization for attention and language (Gazzaniga, 2000). Differences in hemispheric side for lateralized brain functions need not be accompanied by impaired function, as proposed by lesion studies (Saykin et al, 1989; Strauss et al, 1990; Staudt et al, 2002). Rather, all patterns of cerebral lateralization may be implemented in the healthy human brain. The present results add to the evidence that atypical lateralization as such does not reflect disorganization of the neural basis of language. It is still unknown, however, why some people develop an atypical pattern of hemispheric lateralization, while the majority of healthy individuals adheres to the standard pattern. Genetic, hormonal, and environmental factors may affect embryonic development and the side of cerebral lateralization, an issue for further investigation.
Footnotes
Acknowledgements
We are grateful to Jochen Hempelmann for his help in data analysis.