
Abstract
Neonatal hypoxia-ischemia (HI) is a major contributor to many perinatal neurologic disorders and, thus, the search for therapies and effective treatments for the associated brain damage has become increasingly important. The tetracycline derivative, doxycycline (DOXY), has been reported to be neuroprotective in adult animal models of cerebral ischemia. To investigate the putative neuroprotective effects of DOXY in an animal model of neonatal HI, a time-course study was run such that pups received either DOXY (10 mg/kg) or VEH immediately before hypoxia, 1, 2, or 3 hours after HI (n=6). At 7 days after injury, the pups were euthanized, and the brains were removed and processed for immunohistochemical and Western blot analyses using antibodies against specific markers for neurons, apoptotic markers, microglia, oligodendrocytes, and astrocytes. Results showed that in vulnerable brain regions including the hippocampal formation, thalamus, striatum, cerebral cortex and white matter tracts, DOXY significantly decreased caspase-3 immunoreactivity (a marker of apoptosis), promoted neuronal survival, inhibited microglial activation and reduced reactive astrocytosis compared with VEH-treated HI pups. These effects were found to occur in a time-dependent manner. Taken together, these results strongly suggest that doxycycline has potential as a pharmacological treatment for mild HI in neonates.
Introduction
Hypoxia-ischemia (HI) is a common form of perinatal brain damage and is a major contributor to the neurologic entities of cerebral palsy, epilepsy, learning disabilities and mental retardation (Vannucci, 2000). Hypoxia-ischemia launches a cascade of biochemical alterations that are initiated during the course of the insult and proceed well into the recovery period after resuscitation (Vannucci, 1993; Vannucci and Perlman, 1997), leading to both immediate and delayed brain cell death. Because the window for effective clinical intervention might be particularly narrow in the fetus and the newborn because of the often slow and subtle presentation of the onset of the insult, recent focus has been on defining and blocking the mechanisms underlying delayed or secondary brain injury (Nakajima et al, 2000; du Plessis and Volpe, 2002). Key players in delayed brain injury, such as apoptosis and the inflammatory response, have been reported to play a significant role in the evolution and propagation of the brain injury associated with HI (Johnson et al, 2001; Arvin et al, 2002; Cheng et al, 1998; Northington et al, 2001; Nakajima et al, 2000). Previously, inflammation has been shown to be involved in the pathogenesis of traumatic brain injury, adult cerebral ischemia and neurodegenerative disorders including Alzheimer's and Parkinson's diseases (Yrjanheikki et al, 1999; Merrill, 1992; McGeer and McGeer, 1995). Because microglia are the innate inflammatory and resident tissue macrophages of the central nervous system (CNS) (McGeer and McGeer, 1995; Magnus et al, 2002; Hanisch, 2002; Streit, 2002; Liu and Hong, 2003) and caspase-3 is the main death effector involved in apoptosis (Han et al, 2002), the changes in these variables with hypoxia and ischemia in neonatal rats were investigated in the present study.
Doxycycline (DOXY) is a lipophillic, second-generation tetracycline derivative that has antiinflammatory actions independent of its antimicrobial actions (Tikka et al, 2001; Yrjanheikki et al, 1998) (Figure 1A). It crosses the blood–brain, blood–cerebral spinal fluid and placental barriers, and thus has excellent brain and body tissue penetration. Doxycycline and another tetracycline, minocycline (MINO, Figure 1B), were previously reported to be neuroprotective in global and focal brain ischemia in adult animals as well as in cell culture models of cell death (Yrjanheikki et al, 1998, 1999; Tikka et al, 2001). These drugs significantly protected hippocampal neurons from global and focal ischemia (Yrjanheikki et al, 1998) and delayed mortality associated with other types of brain injury such as Huntington's disease (Chen et al, 2000). Minocycline has well-documented mechanisms of action; it is known to inhibit the induction of interleukin 1-β converting enzyme (ICE), prevent inducible nitric oxide synthase (iNOS) protein expression, inhibit caspase production and prevent the activation of microglia (Chen et al, 2000; Yrjanheikki et al, 1998, 1999; Tikka et al, 2001). Also, MINO has recently been shown to be neuroprotective in the same animal model of HI as used in our study, even when administered hours after the HI insult (Arvin et al, 2002). Minocycline, though, has a significant side-effect profile. It has potential for vestibular toxicity (Klein and Cunha, 1995) and, through its propensity to bind calcium, markedly affects bone and teeth development. It is therefore not approved clinically for use in neonates. In contrast, DOXY has fewer adverse effects and is approved for use in neonates. Thus, the aim of this study was to investigate if DOXY, through inhibition of brain microglial cell activation and reduction in caspase-3 activation, would protect the neonatal brain from the effects of acute HI and provide therapeutic efficacy from HI-induced brain injury.
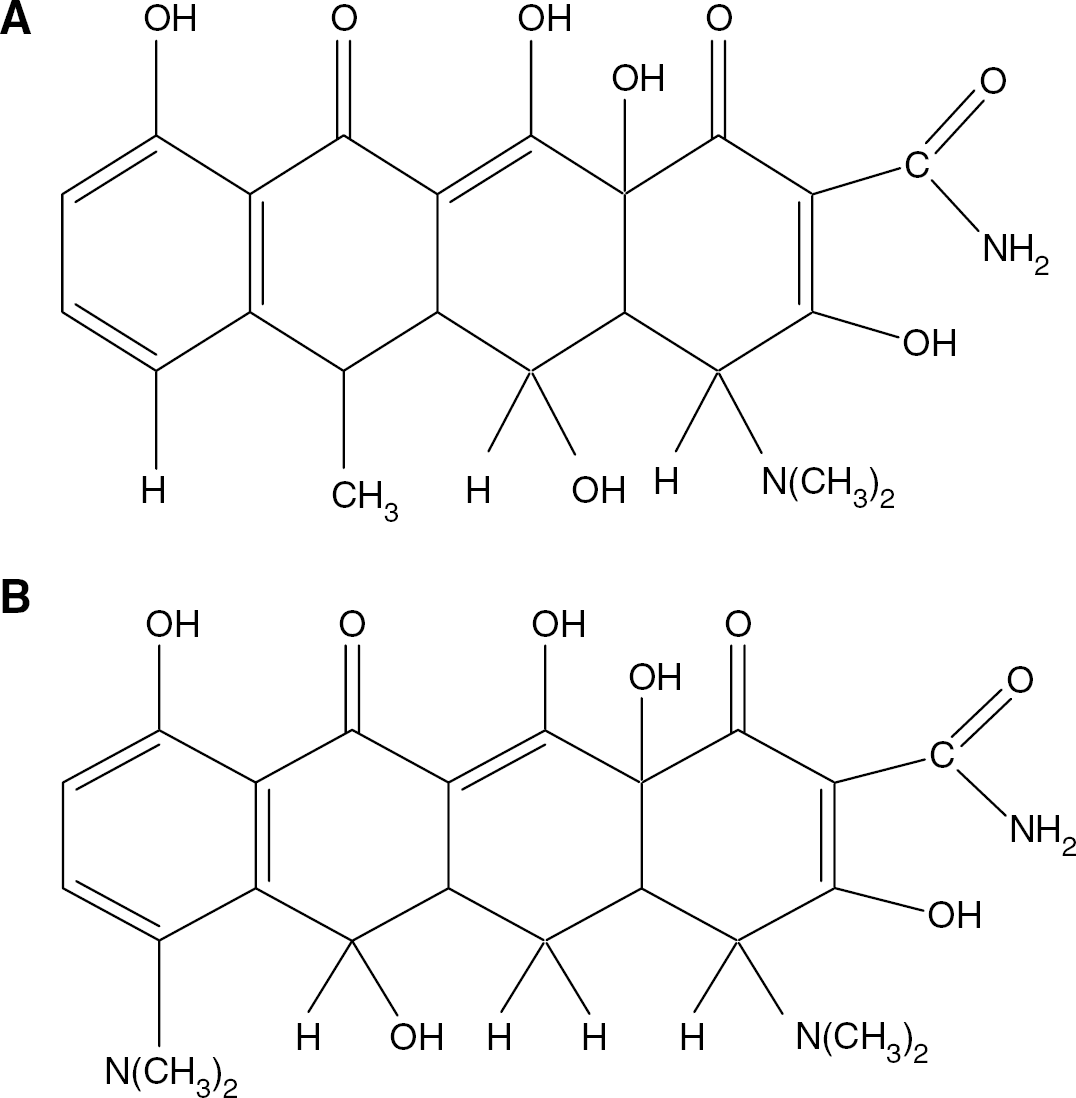
Structures of MINO and DOXY. Doxycycline (
Materials and methods
Animals and Surgical Procedures:
All experiments were performed in accordance with the guidelines set forth by the Canadian Council on Animal Care and were approved by the Health Sciences Animal Policy and Welfare Committee of University of Alberta. On postnatal day 7 (P7), Sprague-Dawley rat pups with weights between 12 and 17 g were anesthetized with a mixture of halothane (5% induction, 1.5% maintenance) and oxygen balanced with nitrogen. We used the well-characterized modified-Levine procedure in which unilateral common carotid artery (CCA) ligation is followed by exposure to hypoxia (Levine, 1960; Vannucci et al, 1999; Rice et al, 1981; Nakajima et al, 2000). The modified Levine's procedure was chosen for these investigations because the P7 rat brain is histologically similar to that of the 32- to 36-week gestation human fetus and because this model produces a reproducible pattern of brain injury that is very similar to the brain damage observed in the human infant (Vannucci et al, 1999; Cheng et al, 1998). To induce HI, on the anterior ventral surface, a small lateral incision was made at the base of the neck and the right CCA was exposed, isolated and permanently ligated using an electrocauterizer. SHAM-operated controls underwent the same surgical procedure, with the exception of the cauterization of the CCA. The wound edges were then opposed and sutured with a 4–0 surgical thread and the pups were returned to their dams. The entire surgical procedure lasted approximately 10 mins. After a 2-hour recovery period, the pups, in groups of 10, were placed in a humidified hypoxia chamber for 2.5 hours at 8% oxygen and balanced nitrogen where normothermia was maintained using a heating pad in the chamber set at 37.5°C. Those pups serving as ischemia-only controls were always excluded from the hypoxia chamber and, likewise, those pups serving as hypoxia-only controls never received a CCA ligation. After the hypoxic period, the pups were returned to their dam until euthanasia.
Drug Administration and Tissue Preparation
Based on previous dose–response studies performed in our laboratory (data not shown) and previous literature reports on the use of DOXY in adult cerebral ischemia (Yrjanheikki et al, 1998), DOXY (SIGMA, 10 mg/kg) or saline vehicle (VEH) was administered intraperitoneally in a volume of 100 μL as a one-time dose immediately before HI, 1, 2, or 3 hours after HI (n=6). This dose of DOXY is below, but approaches, the lowest end of dosing commonly used for antimicrobial effects in children. Injections were performed using a 3/10 cc diabetic syringe with a 29-gauge needle (Kendal). Pups were euthanized by decapitation 7 days after the HI insult and their brains removed and flash frozen in isopentane on dry ice and stored at −80°C until analysis. For immunohistochemical analyses, serial coronal sections (20 μm) throughout the brain were obtained using a cryostat.
Immunohistochemistry
Immunostaining was performed using antibodies recognizing neurons (mouse anti-neuronal nuclei (NeuN), Chemicon, 1:1000), an apoptotic effector (rabbit anti-human cleaved caspase-3, Cell Signaling Technology, 1:100), a microglial marker (mouse anti-rat ED-1, Serotec, 1:100), an oligodendrocyte marker (mouse anti-CNPase, Chemicon, 1:250) and an astrocyte marker (rabbit anti-cow glial fibrillary acidic protein (GFAP), Dako, 1:1000). For each animal, immediately adjacent sections were used for each of the primary antibodies. Briefly, fresh frozen sections were postfixed in buffered formalin and then defatted and dehydrated in a graded series of ethanol washes. Sections were then incubated in a humidifying chamber with 1% hydrogen peroxide to quench endogenous peroxidase enzyme activity, and were subsequently blocked with universal blocking serum (Dako) or normal horse serum mixed with Triton X-100 (0.2% for GFAP, CNPase, NeuN and caspase-3, and 0.5% for ED-1) to reduce nonspecific background staining. Sections were allowed to incubate with the primary antibodies for 1 hour (GFAP, NeuN) or overnight (caspase-3, CNPase and ED-1). After incubation with the species appropriate secondary IgG antibody (all Dako, 1:200) for 30 mins, the sections were carefully washed and incubated for 30 mins with an avidin–biotin complex (ABC 1:100, Vector Laboratories), washed three times and immunoreactivity visualized with 3,3′ diaminobenzedine tetrahydrochloride (DAB, Sigma). In addition to the standard negative control (no primary antibody applied during the course of the immunohistochemistry procedure), a cleaved caspase-3-blocking peptide (Cell Signaling Technology) was also used to further evaluate the specificity of the cleaved caspase-3 antibody reactivity. Specifically, 10 μL of cleaved caspase-3 antibody and 10 μL of blocking peptide in 480 μL Tris buffer was incubated at 4°C for 2 hours before allowing the solution to react to the sections.
The numbers of immunopositive cells in a minimum of five fields (depending on the size of the brain region) in each of the hippocampus, thalamus and cerebral cortex were determined using brightfield microscopy and an unbiased stereological procedure by an investigator blind to the treatment groups. The values obtained in each field were averaged and the means were used in statistical analyses.
Because the values obtained were ordinal and normally distributed, and given that there were no significant differences in the group variances, parametric statistical analyses (analysis of variance (ANOVA)) were applied to the data to evaluate main treatment effects. Where significant main effects were identified, Newman–Keul multiple comparison tests were run. A probability of P<0.05 was chosen to represent statistical significance. Statistical analyses revealed no significant differences in the brains of the obligate control animals at any time point. Therefore, these animals were pooled into a single control group.
Fluorescent immunostaining was performed to corroborate the immunoreactivity seen using the DAB chromagen. The above procedure was used until the application of the secondary antibody. On representative sections, after the excess primary antibody was washed off, an appropriate secondary IgG antibody conjugated with fluorescence was applied for 30 mins. For the cleaved caspase-3 primary antibody, a donkey anti-rabbit Cy2 conjugate or donkey anti-rabbit Texas Red was chosen. For the NeuN primary, a donkey anti-mouse conjugated to FITC was used, and for the ED-1 primary antibody, an anti-mouse IgG conjugated to Texas Red was applied (1:100 for all). The sections were then rinsed and coverslipped using an aqueous mounting medium to prevent autofluorescence.
Western Blotting
To investigate whether cleaved caspase-3 expression was elevated before 1-week post HI, Western blotting was performed using a separate group of pups. These pups received hypoxia and ischemia as mentioned above and received DOXY or VEH 1 hour pre-HI (n=6), and were euthanized 1, 2, and 4 hours after-injury. Brains were removed and their hemispheres were dissected into the following regions—striatum, hippocampus and frontal-parietal cortex—and were stored at −80°C. At the time of analysis, the regions were homogenized in 100 μL of ddH2O and 30 μL aliquots were taken and diluted with an equal volume of 2 × RIPA buffer (2% Igepal CA-630, 1% sodium deoxycholate, 0.2% SDS in PBS) that contained protease inhibitors (complete mini, Roche Biochemical). In all, 50 μg of protein was then loaded onto a 12% SDS-PAGE gel. After electrophoresis, the gel was wet-transferred onto a PVDF membrane at 4°C. The blots were subsequently blocked and incubated with the primary antibody (caspase-3 or cleaved caspase-3 specific, 1:1000) overnight at 4°C. Target proteins were detected by electrochemiluminescence (ECL). Membranes were reprobed with anti-β-actin antibody (42 kDa) to verify the consistency of protein loading between lanes.
Results
No observable alterations in weight gain, suckling or reflexive functions were observed in any pup during the 1-week observational period. Histopathologic evaluation of the brain sections revealed that the neuronal cell loss observed in the VEH-treated HI animals was diffuse, patchy and present throughout most of the brain, although more severe damage was observed in the hemisphere ipsilateral to the ligated carotid artery. Specific areas of the developing brain found to be especially susceptible to this combination of hypoxia and ischemia, as evidenced by significant patterns of neuronal loss and gliosis, were identified in the cerebral cortex, striatum, hippocampus, thalamus, and subcortical and periventricular white matter tracts. Figure 2 illustrates the characteristic pattern of neuronal loss observed in CA1 hippocampus of the hemisphere ipsilateral to carotid ligation. In VEH-treated pups, the average loss was consistently observed to be 30% of total cells in this region. Treatment with DOXY at all time points significantly reduced the neuronal loss observed in CA1 to 10% of total cells in this region.
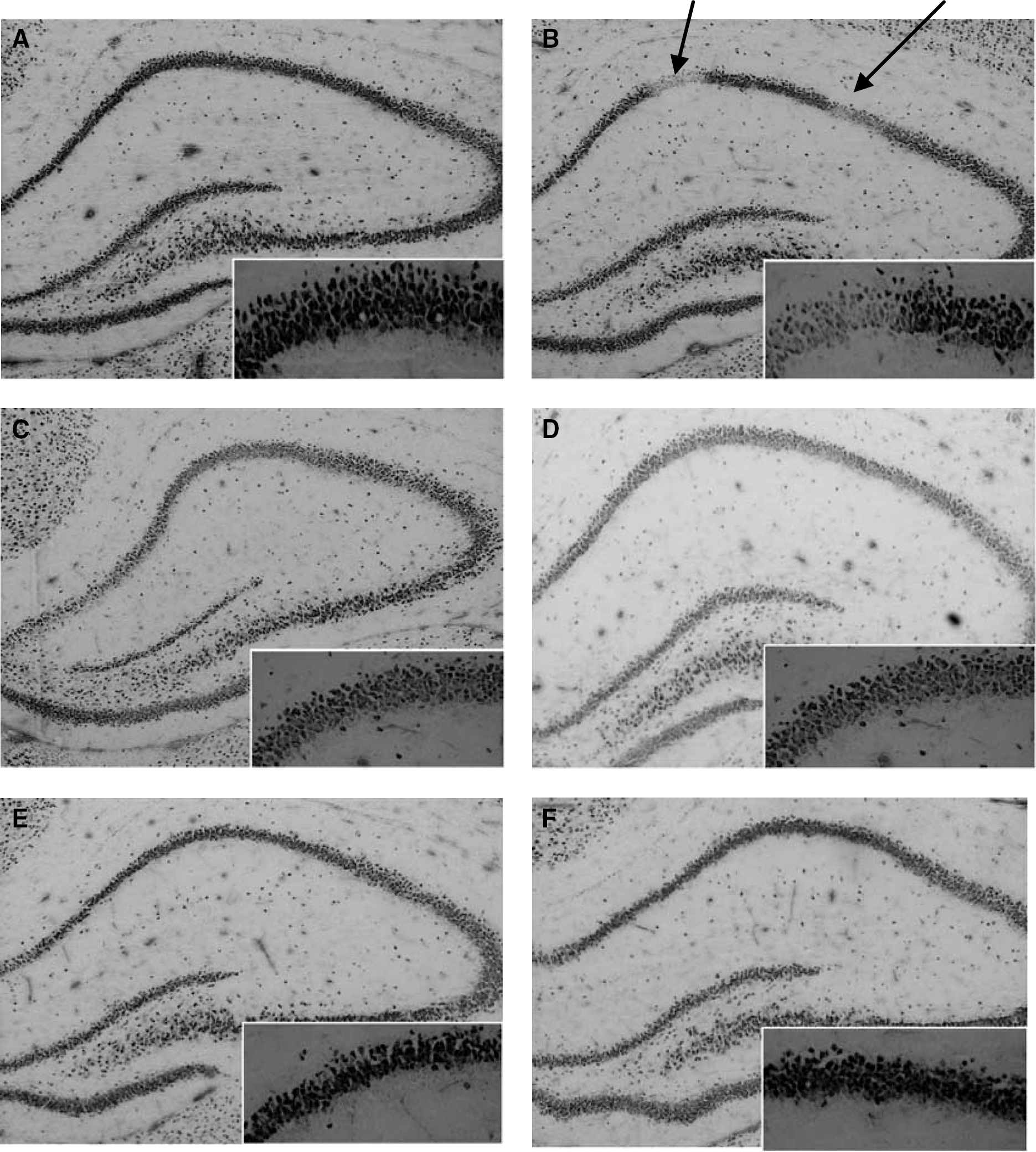
NeuN immunoreactivity. Doxycycline attenuates neuronal loss in the hemisphere ipsilateral to carotid ligation in hypoxic-ischemic rat pups. Representative photomicrographs illustrating patchy neuronal cell loss in the hippocampus of vehicle-treated HI rat pups (
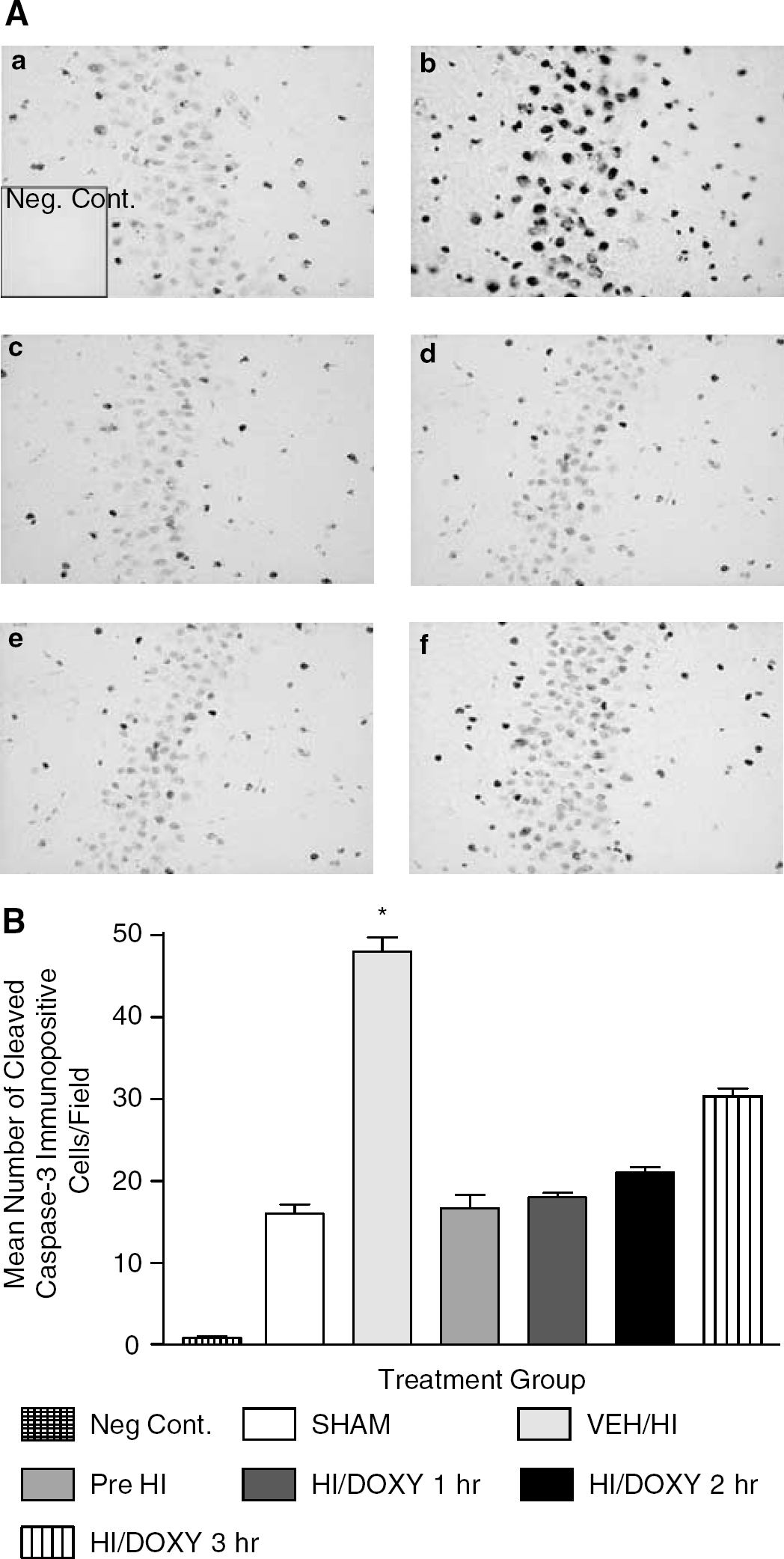
Cleaved caspase-3 immunoreactivity was seen as dense, dark and coarse granular deposits within cells. Figure 3 illustrates the specificity of the cleaved caspase-3 immunostaining performed and shows the difference in staining patterns in sections exposed to the cleaved caspase-3 antibody and a specific blocking peptide, as well as the difference caspase-3 immunoreactivity in the ipsilateral and contralateral cerebral cortex. It was found that cleaved caspase-3 immunoreactivity was increased in the ipsilateral but not the contralateral hemisphere of the ischemic brain. Using antibodies specific for neurons (NeuN) and cleaved caspase-3, and appropriate secondary antibodies conjugated to fluorochromes, double staining was performed to pinpoint the cellular location of the activated caspase-3. Activated (cleaved) caspase-3 was observed colocalized to neurons (Figures 4A–C). Large numbers of cleaved caspase-3 positive nonneuronal cells were also seen in white matter, and these were identified as oligodendrocytes (Figure 4D). Figure 5 displays representative photomicrographs and quantitative analyses of caspase-3 immunoreactivity in the hippocampus of brain sections obtained from SHAM, VEH-treated and DOXY-treated HI rat pups. There was a basal level of cleaved caspase-3 immunoreactivity observed in the SHAM animals. The number of cells positive for cleaved caspase-3 was significantly increased (P<0.05) in VEH-treated HI animals (48±3.0) compared with the SHAM (16±2.0) and all DOXY-treated animals (Pre DOXY+ HI=16.7±2.8; DOXY 1 hour after HI=18±1; DOXY 2 hours after HI=21±1.3; DOXY 3 hours after HI=30±2.5). Further, the numbers of cleaved caspase-3 immunopositive cells were increased in the animals that received DOXY 3 hours after HI compared with animals that received DOXY pre-HI, 1 and 2 hours after HI, although this was not significant. Similar results were observed in the thalamus and cerebral cortex (data not shown). The basal levels of caspase-3 activation were quantified in the SHAM-operated animals and it was found that DOXY did not affect the basal caspase-3 activation. Fluorescent staining performed further illustrated the decrease in the caspase-3 immunoreactivity with the administration of DOXY (data not shown). The Western blotting performed was consistent with the immunochemical results. It was found that DOXY decreased cleaved caspase-3 protein expression (Figure 6B) and increased caspase-3 protein expression as compared with vehicle-treated HI pups (Figure 6A). It was observed that 1 hour after HI in the striatum of a P7 rat, there was a significant upregulation of cleaved caspase-3 protein expression and that the administration of DOXY significantly attenuates this upregulation.
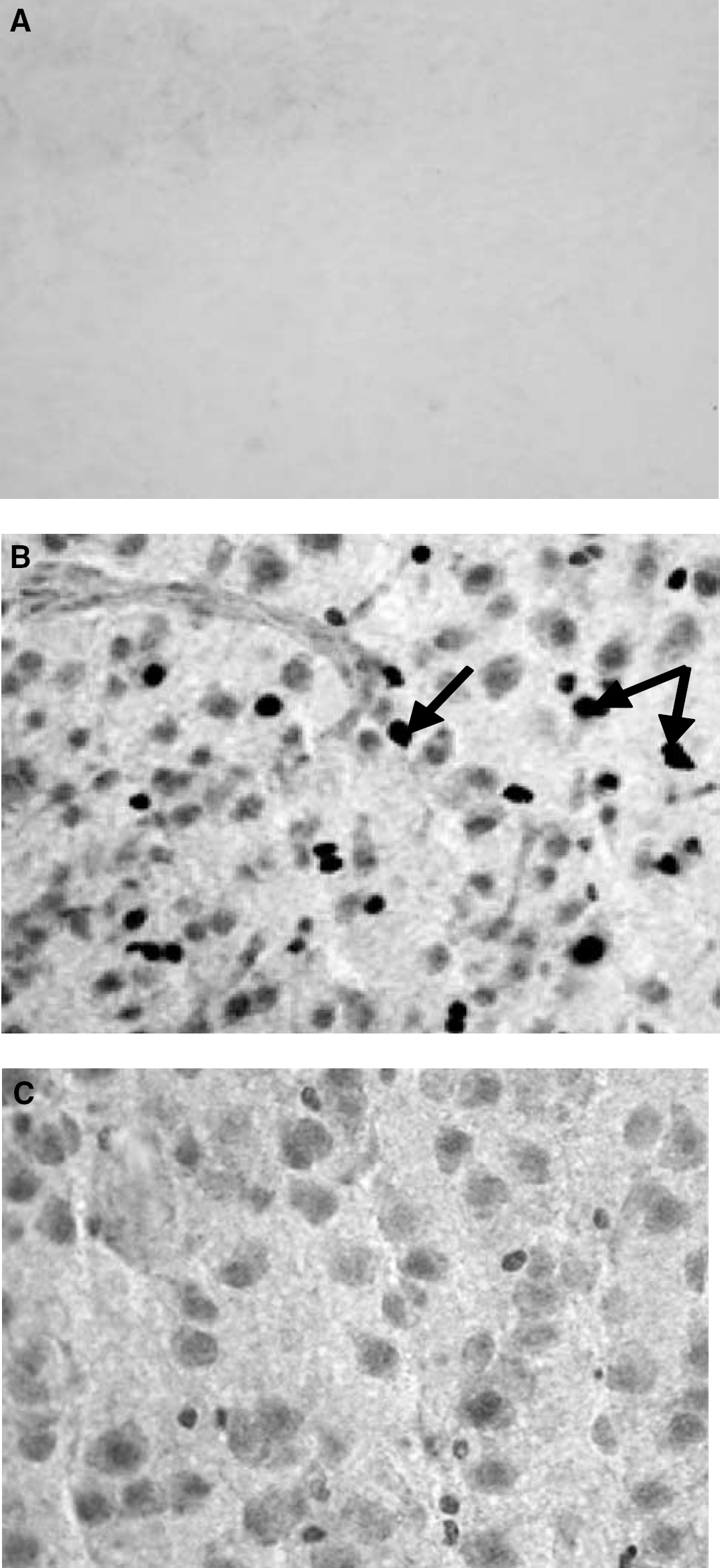
Cleaved caspase-3 immunoreactivity. Cleaved caspase-3 immunoreactivity is different in the ipsilateral and contralateral hemispheres of the neonatal brain subjected to HI. Representative photomicrographs of cleaved caspase-3 immunopositive cells in the cerebral cortex of postnatal day (P) 10 rat brains shown 3 days after common carotid ligation and exposure to hypoxia for 2.5 hours at P7. The right cerebral cortex (
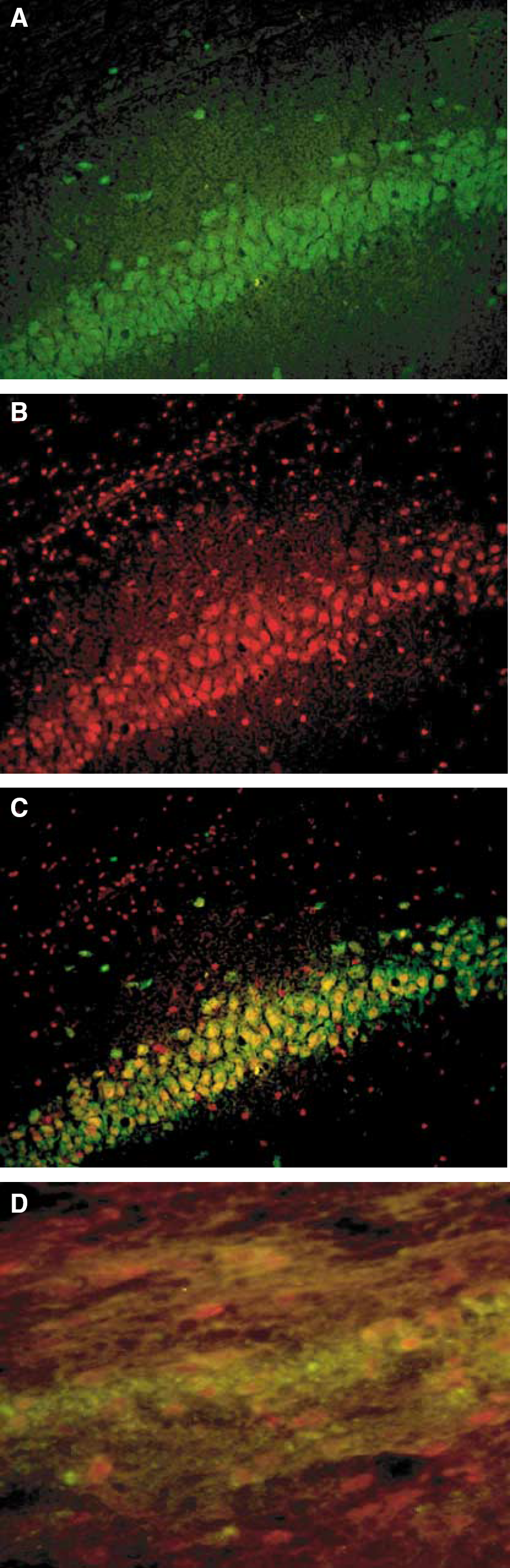
Neurons and oligodendrocytes are cleaved caspase-3 positive. Double immunostaining for (
(
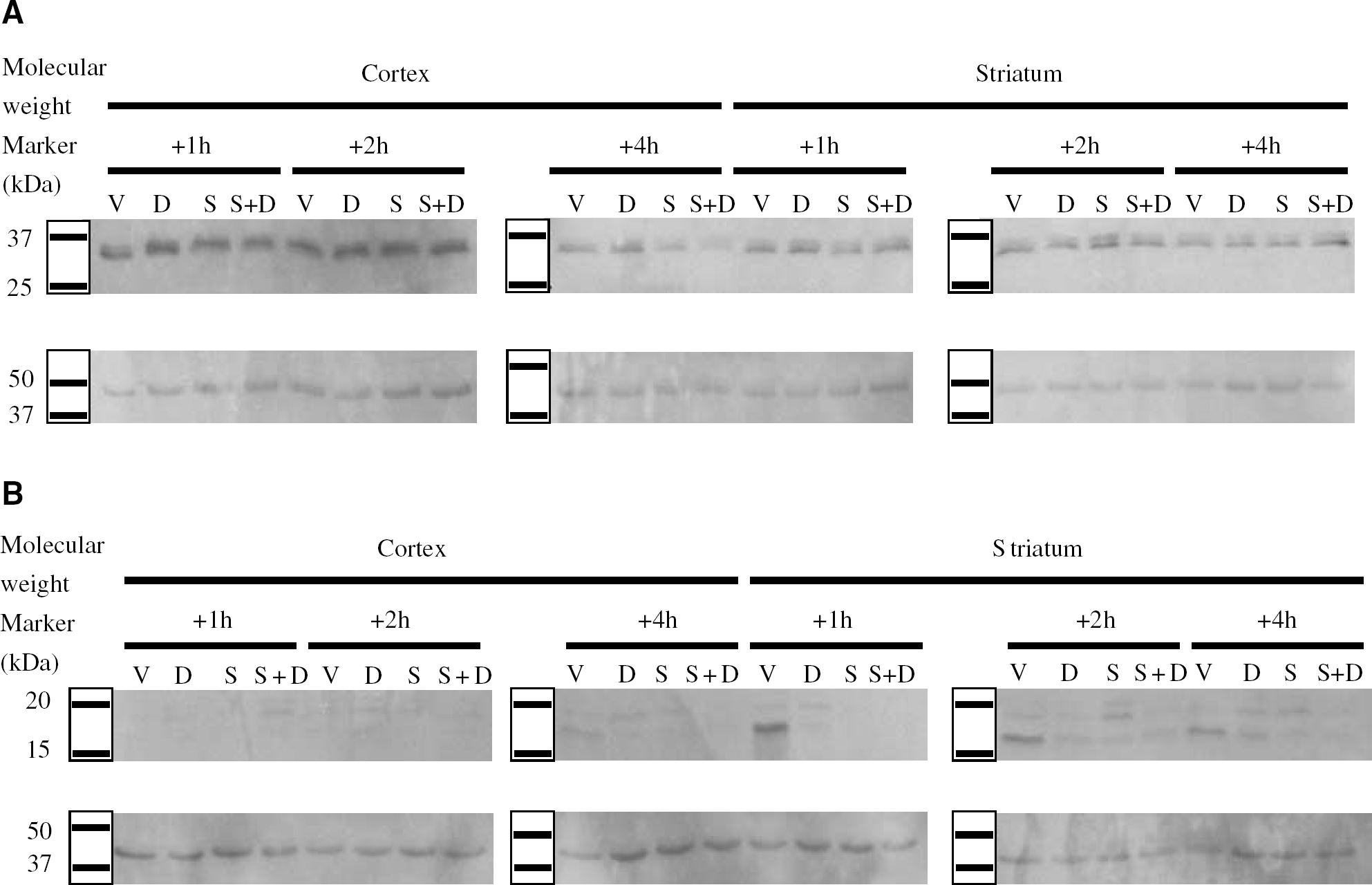
Doxycycline decreases cleaved caspase-3 protein expression and increases caspase-3 protein expression after HI. Representative Western blots depicting decreased cleaved caspase-3 protein expression and increased caspase-3 protein expression in the striatum and fronto-parietal cortex of HI pups in response to treatment with DOXY. Caspase-3 (
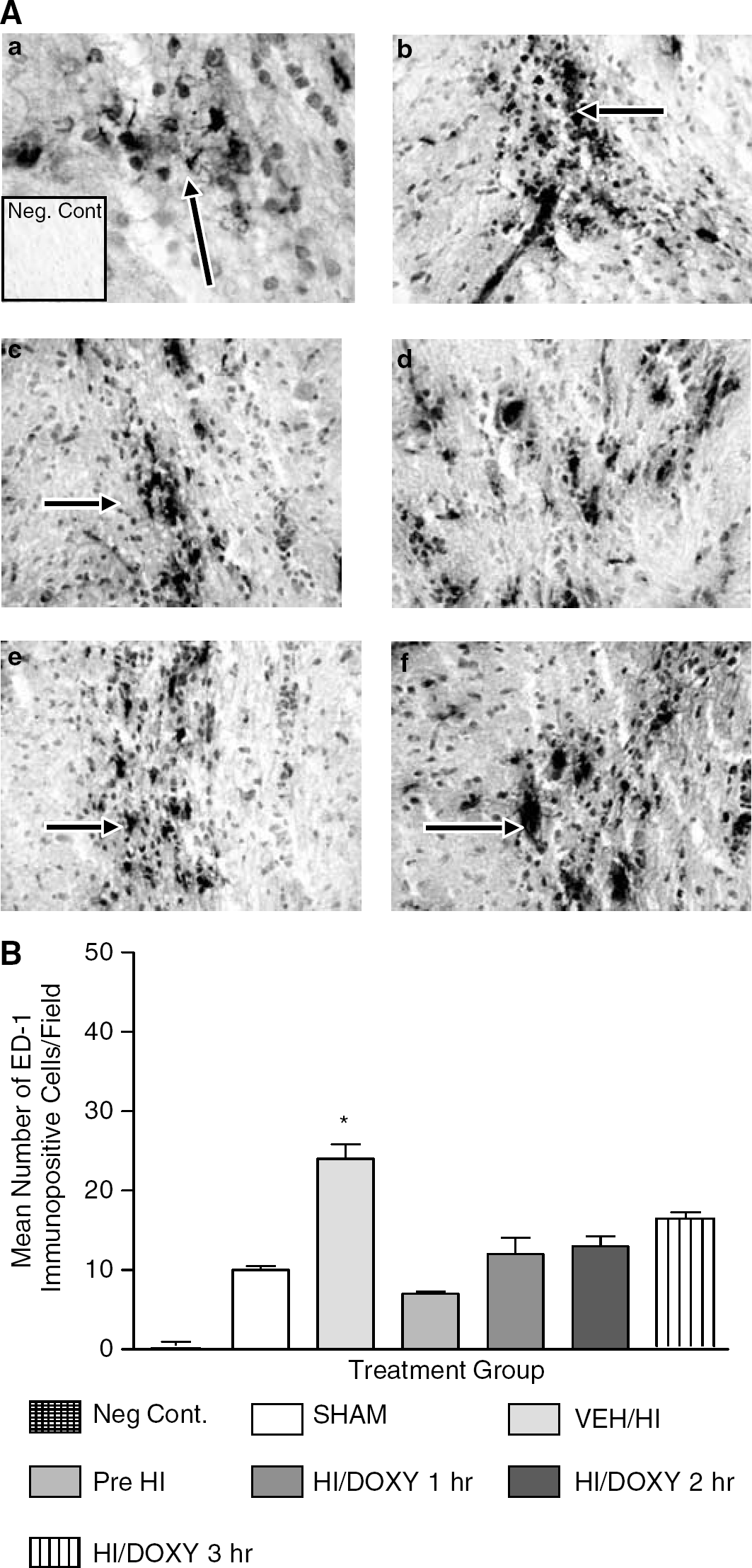
We also evaluated the ability of DOXY to inhibit microglial activation. Figure 7A displays representative photomicrographs of the hippocampus in brain sections obtained from SHAM, VEH-treated and DOXY-treated HI rat pups. In the SHAM animals, few activated microglia were observed. ED-1-positive microglial cells in this group maintained the ramified morphology of resting microglia (Figure 7A plate(a)). In contrast, in sections obtained from the vehicle-treated HI pups, the microglia displayed dramatic morphological changes from resting ramified cells to activated spheroid microglia and became more numerous in response to injury (Figure 7A plate(b)). Figure 7B displays quantitative analyses of ED-1 immunopositive cells. ED-1 immunoreactivity was significantly increased (P<0.05) in VEH-treated HI (24±2.6) compared with SHAM (10±0.70) and DOXY-treated animals (Pre-DOXY+HI=7±0.4; DOXY 1 hour after HI=12±3.0; DOXY 2 hours after HI=13±1.80; DOXY 3 hours after HI=16±2.4). It was found that the inhibition of microglial activation occurred in a time-dependent manner. Further, as shown in Figure 7, the numbers of ED-1 immunopositive cells were increased in animals that received DOXY 3 hours after-HI compared with animals that received DOXY pre-HI, 1 and 2 hours after HI, although this was found not to be significant. Again, similar results were observed in the thalamus and cerebral cortex (data not shown). Fluorescent staining performed further illustrated the reduction in activated microglia with DOXY administration (data not shown).
(
Reactive astrocytes were observed in all animals in the study. Glial fibrillary acidic protein was heavily expressed in mature astrocyte processes and its expression increased after injury with concomitant changes in the morphology of astrocytes with the administration of DOXY. The basal level of reactive astrocytes present in a SHAM animal is shown in Figure 8A. These astrocytes were very delicate, with small cell bodies and thin projections. In contrast, the astrocytes observed in the vehicle-treated HI pups (Figure 8B) were hypertrophic, with large, swollen cell bodies and thick projections. With administration of DOXY, astrocytes were less hypertrophic, less intensely GFAP immunoreactive, and exhibited thinner projections, similar to those observed in the SHAM animal. Consistent with our results from caspase-3 and ED-1 immunoreactivity, the GFAP immunoreactivity and astrocyte morphology changed in a time-dependent fashion with administration of DOXY after the HI insult.

Glial fibrillary acidic protein (GFAP) Immunoreactivity. Doxycycline changed the morphology of astrocytes in neonatal rats subjected to HI. GFAP positive astrocytes were present in all experimental and control animals. The SHAM animals (
Discussion
In this study, we assessed whether DOXY would protect the neonatal rat brain from HI-induced brain damage. We showed that a single dose of DOXY administered immediately before, 1, 2, or 3 hours after HI to neonatal rats significantly decreased apoptotic signaling (caspase-3 immunoreactivity/protein expression), microglial activation (ED-1 immunoreactivity) and promoted neuronal cell survival. Previous studies have shown that a significant proportion of neuronal cell death in brain development and pathology has features of apoptosis, with prominent caspase-3 activation (Roth and D'Sa, 2001; Silverstein et al, 1997; Pulera et al, 1998; Gill et al, 2002; Deckwerth et al, 2001; McDonald et al, 1997; Hu et al, 2000; Wang et al, 2001; Morrison et al, 1998; Northington et al, 2001; Cheng et al, 1998; Nakajima et al, 2000). Cleaved caspase-3 immunoreactivity was found in both hemispheres of the neonatal brain subjected to HI. It was found that the ipsilateral hemispheres of the HI brain had more caspase-3 immunoreactivity and more neuronal loss as compared with the contralateral side, indicating the consistency in the chosen model and the expected pattern of brain damage with this model (Cheng et al, 1998; Vannucci et al, 1999). Regarding our results, DOXY's mechanism of action was associated with a significant decrease in the activation of caspase-3 and, thus, a reduction in the amount of apoptosis that occurred during HI injury. Colocalization of cleaved caspase-3 with fluorescent NeuN and CNPase indicated that both neurons and oligodendrocytes were dying via apoptosis and, thus, DOXY was attenuating the production of this final effector of apoptotic cell death. This finding supports the conclusion that DOXY decreased cleaved caspase-3 protein expression while caspase-3 protein expression was upregulated as compared with the VEH-treated pups. The decrease in apoptotic cell death was observed in various vulnerable regions throughout the neonatal brain, including the CA1 region of the hippocampus as well as the dentate gyrus, cortex, and striatum (data not shown). The decrease occurred in a DOXY-administration, time-dependent manner, and strongly suggests that DOXY may promote neuronal survival by interfering with pathologic apoptosis and preventing the cleavage of the pro-form of caspase-3 to the active form.
Our results are consistent with the previous notion that apoptosis might be the most prominent cell death mechanism in the neonatal brain as compared with adult animals (Hu et al, 2000; Gill et al, 2002; Pulera et al, 1998). It is thought that the involvement of caspase-3 in the pathogenesis of cell death after HI declines during neuronal maturation and that caspase-3 plays a major role in the death of immature neurons after acute HI (Wang et al, 2001; Hu et al, 2000). Other studies have shown that inhibition of caspases protects neurons against HI-induced injury and protects the neonatal rat brain from HI injury (Han et al, 2002; Cheng et al, 1998). It is therefore postulated that inhibition of caspase-3 is an important aspect of neuroprotection associated with neonatal HI. Thus, the ability of DOXY to promote cell survival via the decrease in caspase-3 expression provides a mechanism of action for DOXY and illustrates its potential as a therapy for HI brain injury.
Our data also show that DOXY significantly reduced ED-1 immunoreactivity, a marker for activated microglia/macrophages in HI pups, suggesting that DOXY is a potent inhibitor of microglia. As cells of macrophage potential, microglia need appropriate stimulation to enter a stepwise transformation to develop the features and functions of full-blown macrophages (Hanisch, 2002). Blocking the activation of microglia may inhibit phagocytosis, inhibit release of cytotoxic molecules, preclude entry into certain cell death pathways, thus preventing the subsequent demise of neurons that occurs after the activation of these cells.
Glial fibrillary acidic protein immunohistochemistry was undertaken in these investigations because damage to the CNS is usually accompanied by hypertrophy and hyperplasia of astrocytes; therefore, increased GFAP immunoreactivity was taken as a reliable marker of CNS insult (Todd and Butterworth, 1998). In addition to its role as an inhibitor of apoptosis and microglial activation, DOXY had modulatory effects on astrocytes. Doxycycline did not reduce the number of reactive astrocytes, but produced an altered morphology of the astrocytes present. Our results are also consistent with previous data that suggest that the mechanism of action associated with the activation of microglia and astroglia are independent of each other and mediated through different pathways (Yrjanheikki et al, 1998), in that the administration of DOXY did not affect the number of astrocytes present after HI but did reduce the number of microglia present after HI. It also furthers the notion that activated microglia, but not necessarily astrogliosis, contributes to ischemic neuronal cell death and provides more evidence that astrocytes are fulfilling neuronal support roles in HI (Yrjanheikki et al, 1998).
Taken together, the findings from this study indicate that systemic administration of DOXY protects the neonatal brain from HI even when administered 3 hours after the insult. In fact, DOXY may prove to be the most attractive pharmacologic therapy because of its ability to protect the brain against pathological apoptotic neuronal cell death and neuroinflammation secondary to brain injury. Furthermore, DOXY has the least toxic side effects of the tetracyclines and is especially clinically relevant as it is already approved for the use in neonates as an antibiotic. Thus, the results gained from this study indicate that DOXY may have the potential to play an important role in the therapeutic strategy to prevent neuronal cell death due to perinatal hypoxic-ischemia.
Footnotes
Acknowledgements
We are extremely grateful to the Canadian Institutes for Health Research (CIHR), the Stollery Children's Hospital Foundation and the Davey Endowment for Brain Research, for the generous funding provided for this project. A very special thank you to Richard Strel, Veronique Tanay and Sarah Cooper for their guidance, technical assistance and expert advice.