
Abstract
Endothelin-1 (ET-1) is synthesized by endothelial cells and astrocytes in stroke and in brains of Alzheimer's disease patients. Our transgenic mice with ET-1 overexpression in the endothelial cells (TET-1) showed more severe blood–brain barrier (BBB) breakdown, neuronal apoptosis, and glial reactivity after 2-hour transient middle cerebral artery occlusion (tMCAO) with 22-hour reperfusion and more severe cognitive deficits after 30 minutes tMCAO with 5 months reperfusion. However, the role of astrocytic ET-1 in contributing to poststroke cognitive deficits after tMCAO is largely unknown. Therefore, GET-1 mice were challenged with tMCAO to determine its effect on neurologic and cognitive deficit. The GET-1 mice transiently displayed a sensorimotor deficit after reperfusion that recovered shortly, then more severe deficit in spatial learning and memory was observed at 3 months after ischemia compared with that of the controls. Upregulation of TNF-
INTRODUCTION
Dementia is closely associated with ischemic/reperfusion (I/R) injury and Alzheimer's disease (AD). More than 25% of patients have stroke developed dementia within 3 months after acute ischemic attack. 1 The AD threshold was found to be significantly lowered by cerebrovascular diseases associated with subcortical infarction. 2 Recently, there is a growing evidence that I/R exacerbates amyloid pathology leading to sporadic AD.3,4 In experimental animal models, it has been shown that vascular stress contributes significantly to cognitive deficit.5–7
Endothelin-1 (ET-1), a potent vasoconstrictor, is implicated in the pathogenesis of stroke. The level of plasma ET-1 is markedly increased by four-fold in patients with acute ischemic stroke 8 and the extent of increase is well correlated with the severity of neurologic outcomes. 9 In the brain, intrastriatal administration of ET-1 peptide induced substantial brain lesion around the injection site. 10 In addition, its receptors, ETA and ETB subtypes are differentially activated in neurons and in glial cells in response to ischemic brain injury. 11 Endothelial ET-1 exerts an important role in neurovascular reactions by increasing blood–brain barrier (BBB) permeability and vasoconstriction after cerebral ischemia.12–14 In our previous report, transgenic mice overexpressing endothelial ET-1 also showed more severe brain damage after transient middle cerebral artery occlusion (tMCAO),15,16 suggesting the contribution of endothelial ET-1 in the neurovascular instability leading to BBB breakdown and ischemic brain injury after stroke. Animal models with ET-1 injection to induce cerebral ischemia have been used to investigate β-amyloid (Aβ) toxicity under ischemic stroke. Intracerebroventricular 7 or hippocampal 17 administration of Aβ to those mice was found to exacerbate the AD-like cognitive deficit. The striatal injection of ET-1 causes significant elevation of amyloid precursor protein fragments including Aβ deposition, 18 suggesting that upregulation of ET-1 during ischemic stroke may responsible for the Aβ production and accumulation, which is neurotoxic to the brain cells and results in impairment of the neurovascular unit, such as BBB breakdown. In addition, overexpression of ET-1 in the endothelial cells exacerbates anxiety-like behavior and spatial reference learning and memory impairment after a short-term ischemia with 7-day reperfusion 16 suggesting the importance of endothelial ET-1 in Aβ and dementia.
However, the level of ET-1 mRNA is also induced in the astrocytes in various neurodegenerative disorders, such as stroke and AD in addition to endothelial cells.19,20 Several lines of evidence support the notion that astrocytic ET-1 may be involved in dementia associated with I/R and AD. Similar to ET-1, Aβ is significantly elevated in the plasma of patients having ischemic stroke. 21 Moreover, circulating Aβ can be actively up taken by receptor for advanced glycation end product across the BBB, which leads to cerebrovascular amyloid accumulation and ET-1-mediated vasoconstriction. 22 Autopsy examination of human AD brain reveals that there is an intense ET-1-like immunoreactivity in the astrocytes surrounding the senile plaques. 20 More importantly, elevation in Aβ peptide is detected and colocalized in the reactive astroglia surrounding capillaries 23 and within the neuropil in the rat hippocampus after lesion 24 and in the CA1 and CA4 regions of the dorsal hippocampus after global ischemia. 25 Recently, the link between ET-1, Aβ, and cognitive deficit has been emerged.18,26 These findings suggest that astrocytic ET-1 may have a critical role in the pathogenesis of I/R and AD.
Previously, we have showed that transgenic mice overexpressing ET-1 in the astrocytes (GET-1) exhibit increased neurologic infarct associated with cerebral edema and BBB breakdown after tMCAO.
27
However, the impact of astrocytic ET-1 induction on cognitive function has not been reported. In the current study, the long-term effect of astrocytic ET-1 induction in the development of poststroke cognitive deficit in association with astrocyte-derived amyloid production was investigated. The cognitive function and the neurologic deficits associated with I/R injury and AD were evaluated in the GET-1 mice. Also, the response in amyloid secretion by ET-1 overexpressing astrocytic cells under hypoxia/ischemia (H/I) condition was also examined by
MATERIALS AND METHODS
Animals
All mice were bred and maintained under strictly controlled environmental conditions (i.e., 20°C, a light/dark cycle of 12-hour day/night and free access to sterilized diet and water). All the male GET-1 mice and their nontransgenic control (NTg) (8 to 12 weeks old) were maintained in the same genetic background. GET-1 mice appeared normal and healthy, and displayed no obvious physiologic, gross anatomic, or behavioral abnormalities to those of NTg mice.
27
A total of 71 male homozygous GET-1 and NTg mice, weighing 20 to 25 g, were used (four groups (NTg Sham,
Transient Focal Cerebral Ischemia
Intraluminal occlusion of the right transient middle cerebral artery (tMCAO) was used as a model of transient focal cerebral ischemia in this study. Detailed surgical procedures were described in our previous publication, 27 except that the durations of the occlusion and the reperfusion were amended to 30 minutes and a longer term (i.e., up to 5 months after ischemia), respectively. In brief, after the experimental mice were gas anesthetized (induction: 2% halothane in 70% N2O/30% O2; maintenance: 1% halothane in 70% N2O/30% O2) with a stabilized body temperature (i.e., rectal temperature, 37°C), a nylon monofilament (Johnson & Johnson, Brussels, Belgium) coated with impression material (3 mol/L Dental Products, St Paul, MN, USA) was inserted into the right internal carotid artery to block the origin of right middle cerebral artery. Laser Dropper flowmetry (Perimed, Jarfalla, Sweden) using an optic fiber (with probe 418–2) placed directly on the skull (2 mm posterior and 6 mm lateral to bregma) was used for continuous measurement of the regional cerebral blood flow (rCBF) throughout the surgical procedure to ensure successful induction of ischemia and reperfusion. A decrease of 70% or more in rCBF was considered as successful induction of ischemia. The experimenter who performed the tMCAO was blinded to the genotypes.
Assessment of Neurologic Deficits, Cerebral Edema, and Infarct Size
To assess the neurologic and sensorimotor deficits associated with I/R injury, a 4-point scoring paradigm 28 was used in a double-blinded manner at 1, 3, and 7 day(s) after cerebral ischemia. The criteria were briefly summarized as follows: 0, no observable deficit; 1, failure to extend forepaw in opposite side (wrist drop); 2, circling in walking toward the contralateral side; 3, loss of walking ability and righting reflex. Animals that died during reperfusion period would be excluded for data analysis.
For
For the measurement of brain shrinkage, brains were sliced into coronal sections with 2 mm thickness and stained with 2% 2,3,5-triphenyltetrazolium chloride (TTC). The posterior surface of each brain slice was photographed and analyzed by the image analysis program, the brain integrity was determined by calculating the percentage of the area in the ipsilateral hemisphere relative to that of the corresponding contralateral hemisphere. 27
Cognitive Function Assessment
Forty-seven age-matched male mice were grouped into four cohorts based on the genotype and treatment as follows: (1) NTg sham; (2) GET-1 sham; (3) NTg tMCAO; and (4) GET-1 tMCAO. All mice were habituated for 7 days in the experimental room before the behavioral assessment. The spatial reference learning and memory of the experimental mice were assessed 3 months after tMCAO by Morris water maze (MWM) according to a well-established protocol published elsewhere 29 with slightly modification. Briefly, the water maze procedures were divided into three sessions: (1) the visible platform test (2 days, Days 1 and 2), in which the pool was filled with water, and the platform was submerged 1 cm below the water surface, a flag was placed on the platform to increase its visibility. The visible platform test composed of two consecutive training blocks of eight trials per day to ensure normal sensorimotor ability of the mice following surgical procedures. The time required for locating the visible platform was averaged per block and was expressed as escape latency. Mice failed to complete this task (i.e., escape latency longer than 2 s.e.m. of the corresponding sham-operated group) were eliminated for subsequent test; (2) the hidden platform test (6 days, Days 3 to 8), which composed of six consecutive training blocks of four trials per day, was conducted to aim for tracing the ability of spatial learning of the mice. The time required for locating the hidden platform was averaged per block and was expressed as escape latency; (3) the probe trial test (3 days, on Days 5, 7, and 9), which composed of three intercalated 60-seccond free swimming without platform, was conducted to assess the ability of the spatial retention of the mice during different phases of learning. Time spent on the target quadrant (i.e., where the platform previously located) was recorded by an automated video tracking system (EthoVision professional version, Noldus Information Technology, Wageningen, The Netherlands) and was expressed as the percentage of spatial bias. In all, 25% of spatial bias was considered as no memory retention, in which mice swam randomly in the pool. The experimenter was blinded to the genotypes.
Tissue Collection and Processing for Histologic and Biochemical Analyses
The brains from the experimental animals were harvested from deeply anesthetized animals with sodium pentobarbital followed by cervical dislocation. The ipsilateral and contralateral brains were separated along the midline of the hemispheres. For histologic analysis, separated brains were fixed in 4% paraformaldehyde for overnight, followed by serial dehydration in graded ethanol and chloroform before paraffin embedding. For Western blot analysis, ipsilateral hippocampus was isolated and snap-frozen in liquid nitrogen and stored at −80°C until further protein extraction.
Immunocytochemical Analysis
Rehydrated sections were incubated with appropriate blocking serum for 1 hour at room temperature and primary antibodies against, TNF-α (1:500; Santa Cruz, CA, USA) and cleaved caspase-3 (1:500; Cell Signaling, Beverly, MA, USA), which were performed in the ipsilateral sides of hippocampal regions, and GFAP (1:5,000; Z0334; Dako, Glostrup, Denmark) or ET-1 (1:1,500; T-4050; Peninsula Laboratories, Inc., Belmont, CA, USA), which were performed in the ipsilateral sides of penumbra regions, for overnight at 4°C. The immunocytochemical procedures were described previously. 27 The immunofluorescence signal was obtained by conjugating with FITC-conjugated secondary antibody (1:100; T2769, Molecular Probes, Eugene, OR, USA) for 1 hour followed by DAPI (1:1,000) or Thioflavin-S (Thio-S) (1% in 50% ethanol (w/v)) counterstaining. For quantitative analysis of amyloid burden, four TS-stained sections, with 140 μm apart, were examined in each animal. The number of Thio-S-positive pial vessels, intra-cerebrovessels, and extracellular plaques were counted. Photomicrographs were taken with conventional fluorescent microscopy (Axioplan; Carl Zeiss Inc., Petaluma, CA, USA) or laser scanning confocal microscopy (LSM 510 Meta; Carl Zeiss Inc.). All analyses were made in a double-blinded manner.
Counting for 3,3′-Diaminobenzidine Stained Cleaved Caspase-3 Neuronal Cells in Hippocampus
The DAB (3,3′-diaminobenzidine)-stained cleaved caspase-3 neuronal counts were performed blinded to the genotypes with the aid of Neurolucida 8.0 software (MBF Bioscience Inc., Williston, VT, USA). At least three individual animals of brain sections were used, and over 100 cells were counted per section. Cleaved caspase-3 stained immunocytochemistry sections in the hippocampal regions were viewed at high magnification (X40) under light microscope, and cells with cleaved caspase-3 staining at three regions of interest in the hippocampus were counted. Over 300 cells were counted for each genotype.
Western Blot Analysis
Protein extraction and gel electrophoresis were performed as described previously. 27 The membranes were probed with primary antibodies against GFAP (1:10,000; Z0334; Dako), heat-shock protein 70 (HSP70) (1:2,000; SPA-810; Stressgen, Ann Arbor, MI, USA), peroxiredoxin 6 (Prx6) (1:2,000; ab59543; Abcam, Cambridge, UK), β-actin (1:10,000; A1978). Immunoreactive bands were quantified individually by estimating the number of pixels using an image analysis system (Image J, NIH, USA). The band intensity of the target protein was first normalized with the band intensity of the β-actin and expressed as a ratio of the two bands. The results were expressed as the percentage of the expression level of the target protein of the GET-1 group relative to that of the NTg group by the calculation as follows: (ratio of the individual GET-1 mice÷averaged value of the ratio of the individual NTg mouse) × 100%.
Cell Culture
The astrocyte-like ET-1-over-expressing stable cell line (C6-ET1) was generated by cotransfection plasmid that drives the overexpression of active ET-1 peptide (pGET-1) with plasmid containing puromycin-resistance gene (pPUR) into a D1 TNC1 astrocytic cell line (ATCC, CRL-2005). The puromycin-resistance clones were screened by PCR and were selected by puromycin to ensure a stable integration of pGET-1. Stable clone was characterized and was selected based on the level of ET-1 secretion by ELISA (Amersha, Piscataway, NJ, USA). Mock-transfected clone (C6-Mock) was used as a control in which pMock (i.e., pGET-1 without ET-1 cDNA and SV40 polyA signal) and pPUR were cotransfected. All cells were cultured in Dulbecco's modified Eagle's medium-F12 (Gibco, IMDME; GIBCO, Grand Island, NY, USA) containing 10% fetal bovine serum and incubated in a humidified chamber supplied with 5% CO2 at 37°C for propagation until seedling for experiments.
Determination of the Effect of Astrocytic Endothelin-1-Mediated β-Amyloid Peptides Secretion after In Vitro Hypoxia/Ischemia Challenge
To study the cellular effects of astrocytic ET-1 overexpression on the secretion of the two species of Aβ peptides, Aβ1–40 and Aβ1–42, the C6-ET1 and C6-Mock astrocytes were challenged under H/I or normoxia (N) condition. The H/I condition (i.e., no glucose and no oxygen) was created by culturing cells in a serum- and glucose-free culture medium (i.e., pre-purged with gas mixture without O2) and was maintained in a temperature-controlled anaerobic chamber. The N condition (i.e., normal glucose and normal oxygen) was achieved by culturing cells in the same culture medium except that it was supplemented with 10% serum and 5.5 mM glucose and maintained in an ordinary chamber with standard O2/CO2 level. 30 C6-ET1 and C6-Mock astrocytes were subjected to N or H/I condition for 24 hours with different drugs or vehicle treatment. Culture medium were collected for the measurement of Aβ1–40 and Aβ1–42 peptides being released into the medium by ELISA method (27720, 27721, Immuno-Biological Laboratories, Fujioka-Shi, Gunma, Japan) according to the manufacturer's instructions. Drugs including BQ-123 (selective endothelin A receptor (ETA-R) antagonist) and BQ-788 (selective endothelin B receptor (ETB-R) antagonist) were purchased from Sigma-Aldrich (St Louis, MO, USA). LY294002 (PI3 Kinase inhibitor) was purchased from Cell Signaling. All drugs were dissolved in PBS and were diluted to the desire concentrations with culture medium before administration. Control treatment represents the addition of the PBS to the culture medium as in the drug treatment.
Statistical Analysis
All statistical analyses were performed using the GraphPad Prism 5 (Version 5.01, GraphPad Software, Inc., USA). The data were presented as mean ± s.e.m. Results were analyzed using one-way ANOVA followed by Tukey post test, except Mann–Whitney test for the TTC quantification. For all statistically analysis,
RESULTS
GET-1 Mice Exhibited a Mild but Transient Sensorimotor Deficit after 30 minutes tMCAO
Previously, we showed that GET-1 mice exhibited a severe neurologic deficit as characterized by an impaired sensorimotor deficit after 2 hours of ischemia followed by 22 hours of reperfusion.
27
To preserve the motor function of the mice for subsequent water maze test, the duration of ischemia reduced to 30 minutes. In line with our previous study,
16
no significant differences in rectal temperature and rCBF were detected in the four experimental groups during the whole tMCAO procedure (data not shown). In addition, neurologic assessments revealed a mild sensorimotor deficit in the GET-1 mice only at 1 day after tMCAO (NTg versus GET-1: 0.12 ± 0.06 versus 0.58 ± 0.08,
Neurologic assessment of sensorimotor function after tMCAO
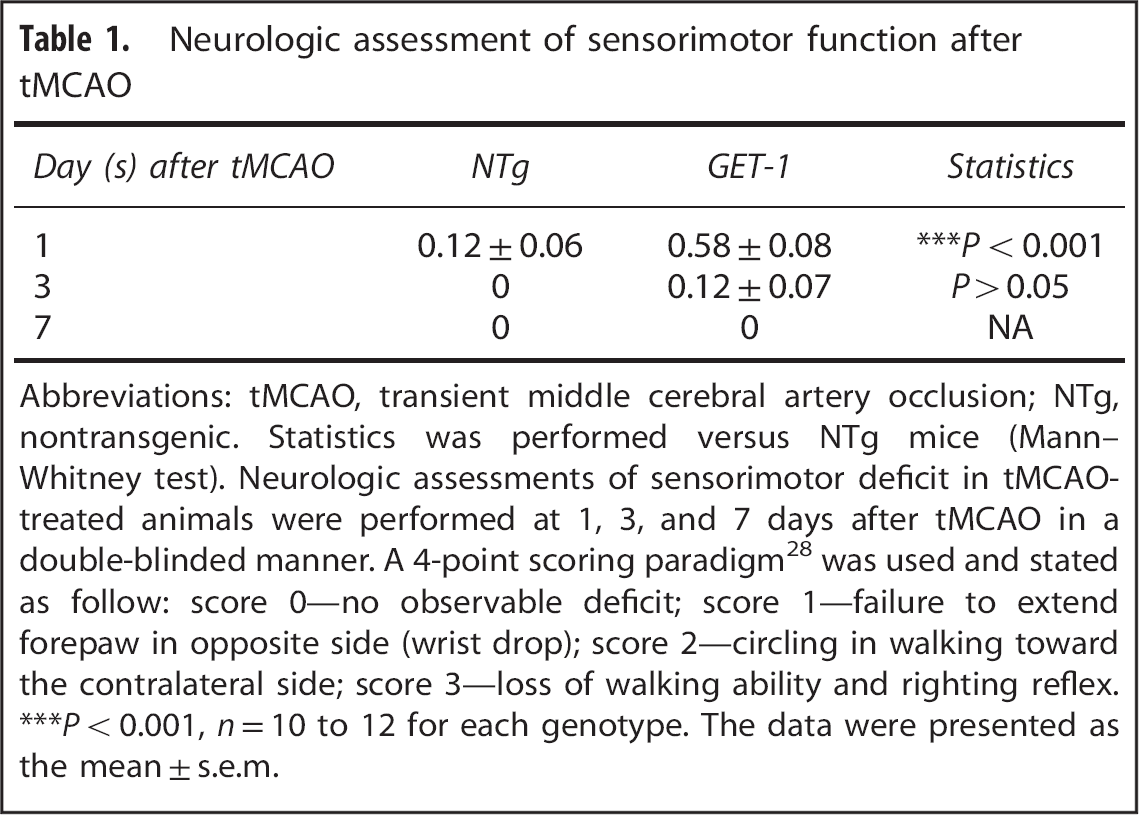
Abbreviations: tMCAO, transient middle cerebral artery occlusion; NTg, nontransgenic. Statistics was performed versus NTg mice (Mann–Whitney test). Neurologic assessments of sensorimotor deficit in tMCAO-treated animals were performed at 1, 3, and 7 days after tMCAO in a double-blinded manner. A 4-point scoring paradigm 28 was used and stated as follow: score 0—no observable deficit; score 1—failure to extend forepaw in opposite side (wrist drop); score 2—circling in walking toward the contralateral side; score 3—loss of walking ability and righting reflex.
Long-Term Astrocytic Endothelin-1 Overexpression Exacerbated Poststroke Cerebral Edema and Neurodegeneration
For
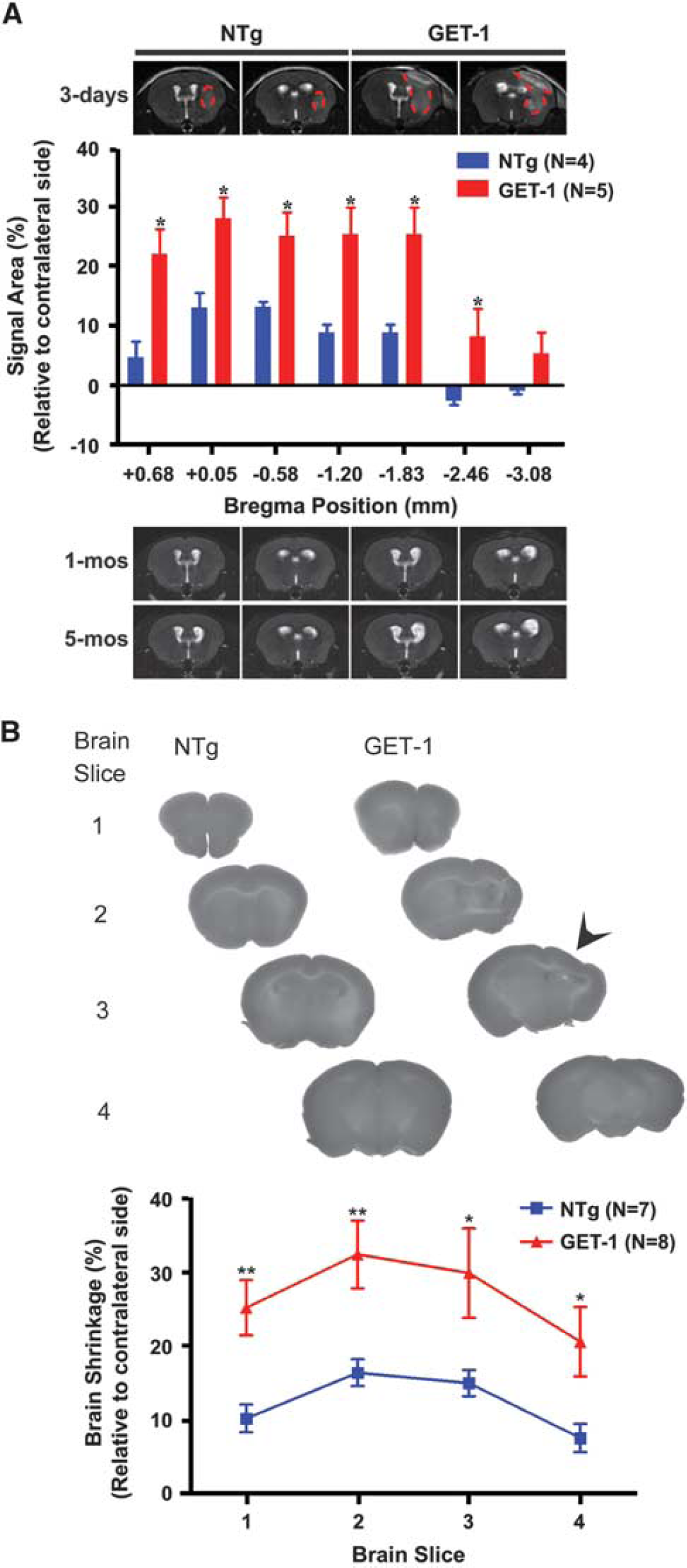
(
Long-Term Astrocytic Endothelin-1 Overexpression Exacerbated Poststroke Learning and Memory Deficit
Hippocampus-dependent spatial reference learning and memory was also evaluated using the spatial memory version of MWM assessment at 3 months after tMCAO. In the visible platform session, no sensorimotor deficit could be detected in all experimental mice as they performed equally well during the whole course of training (Figure 2A). In contrast, in the hidden platform session, tMCAO-operated GET-1 mice showed a significant retardation in the learning process when compared with the sham-operated GET-1 mice and the NTg counterparts as indicated by the
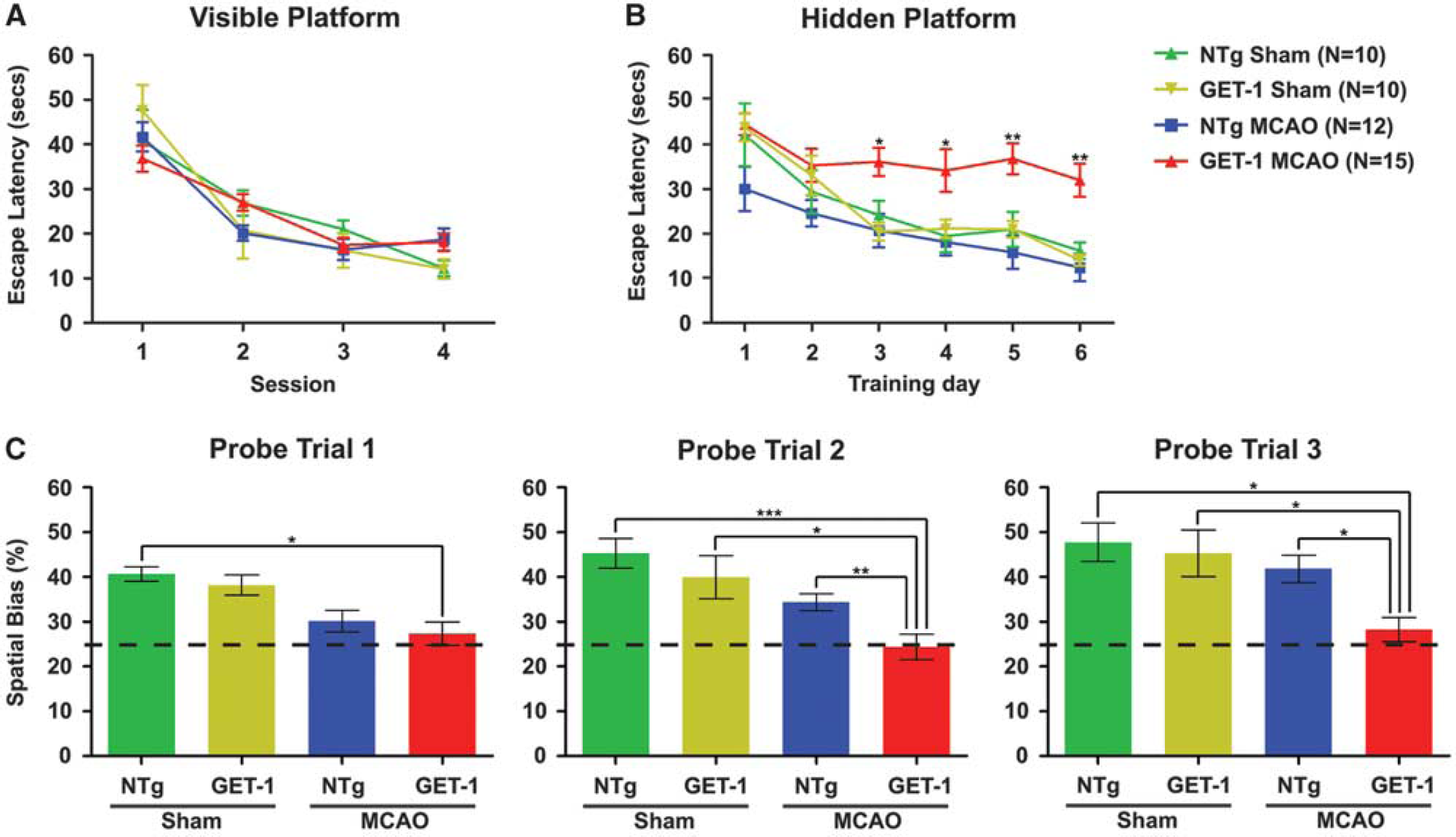
Mean escape latency (seconds) to locate the (
Astrocytic Endothelin-1 Overexpression Exacerbated Ischemic/Reperfusion-Induced Neuroinflammation and Oxidative Stress in the Hippocampus
To determine the neurologic impact of chronic astrocytic ET-1 exposure after mild I/R injury, sign of neuroinflammation and oxidative responses were determined by histologic and biochemical analyses. To correlate with the cognitive deficit, hippocampal region in the tMCAO-operated GET-1 and NTg mice was chosen for comparison. Histologic analysis at 3 days after tMCAO revealed that immunoreactivities of the antigen of TNF-α (Figure 3A) and cleaved caspase-3 (Figure 3B) were highly induced in the pyramidal neurons located at the CA2 region in the GET-1 brain, whereas it was significantly lower in the similar region of the NTg brain. Moreover, the level of oxidative stress as reflected by the expressions of GFAP (Figure 3C), HSP70 (Figure 3D), and Prx6 proteins (Figure 3E) were significantly elevated in the GET-1 hippocampus when compared with that of the NTg mice. Taken together, after a mild ischemic insult, ipsilateral hippocampus may subject to substantial neuroinflammatory and oxidative responses, which leading to progressive neuronal cell death.
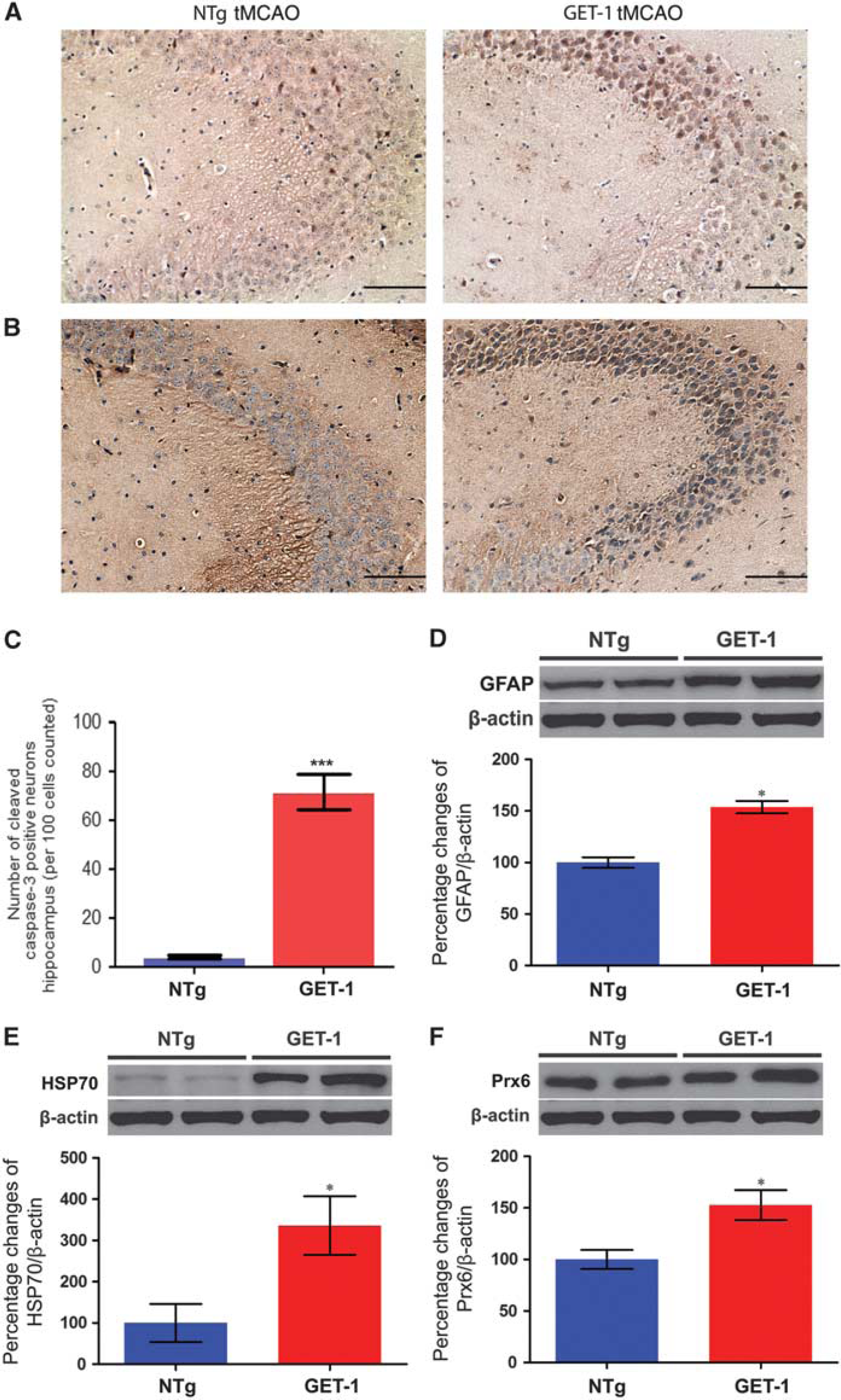
Photomicrographs show the immunoreactivity of (
Colocalization of Endothelin-1-Positive Astrocytes and Thioflavin-S-Positive Amyloid Deposition in the Peri-Vascular Region in the Ipsilateral GET-1 Brain after Mild Ischemic/Reperfusion Injury
To investigate whether there is a positive correlation between increased astrocytic ET-1 expression and amyloid deposition, immunohistologic analysis was performed by costaining ET-1 peptide with Thio-S in the brain sections of the tMCAO-operated GET-1 and NTg mice. In the penumbra region of the GET-1 brain, an intense ET-1-like immunoreactivity was detected in the perivascular region where reactive astrocytes localized, whereas it was generally absent in the similar region of the NTg brain. Similarly, with the same set of brain sections, most of the Thio-S-positive amyloid aggregates were colocalized with the GFAP-positive astrocytes in the perivascular region (Figure 4B). These findings indicate that astrocytic ET-1 exacerbrates cerebrovascular amyloid deposition possibly contributed by the action of reactive astrocytes after triggering by I/R injury.
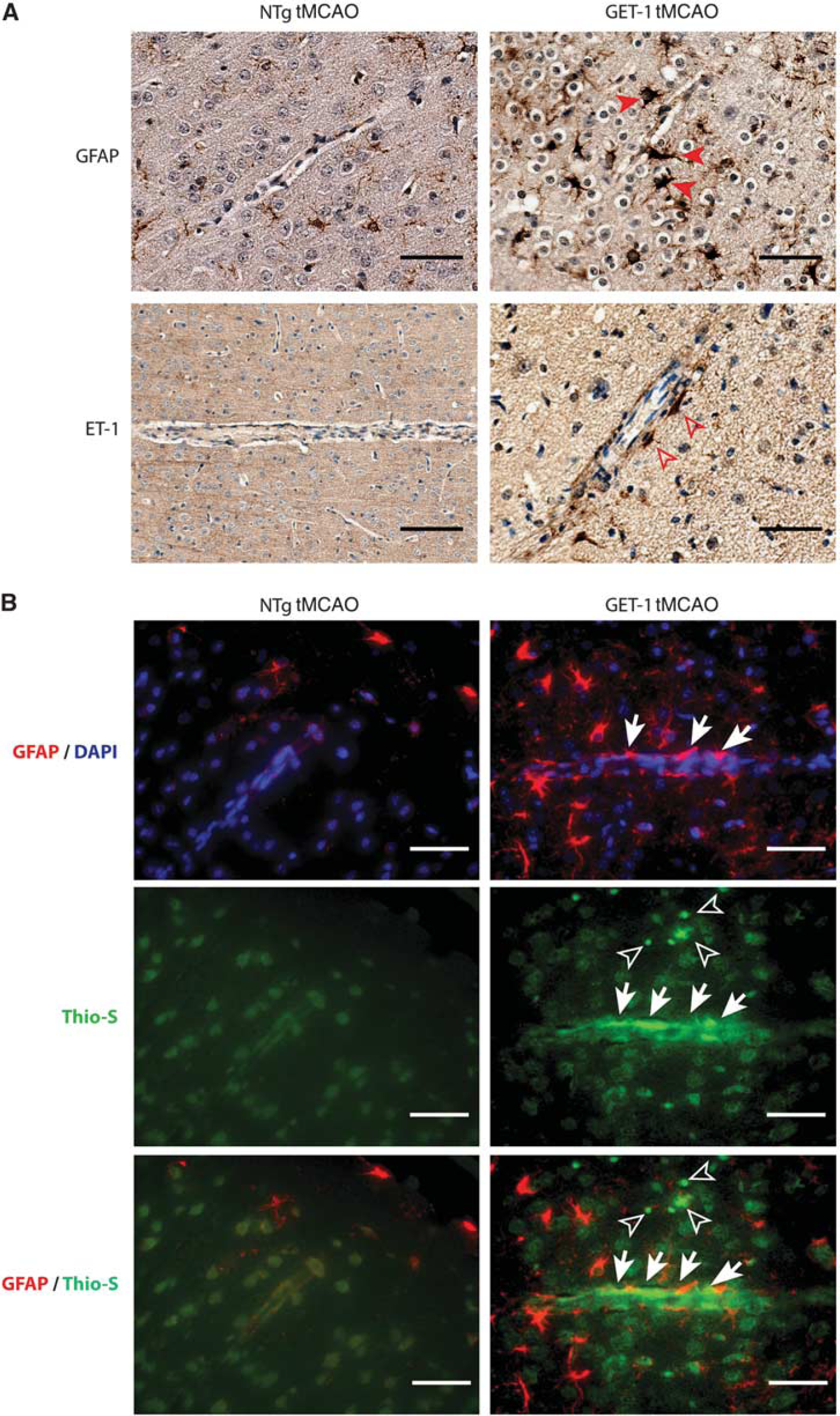
Photomicrographs show the results of immunostaining pattern of peri-vascular (
Both Endothelin A and Endothelin B Receptors Were Involved in PI3K/AKT-Dependent Amyloid Secretion in Endothelin-1 Overproducing Astrocytic Cells
To consolidate the direct effect of astrocytic ET-1 induction and astrocyte-derived amyloid deposition, an
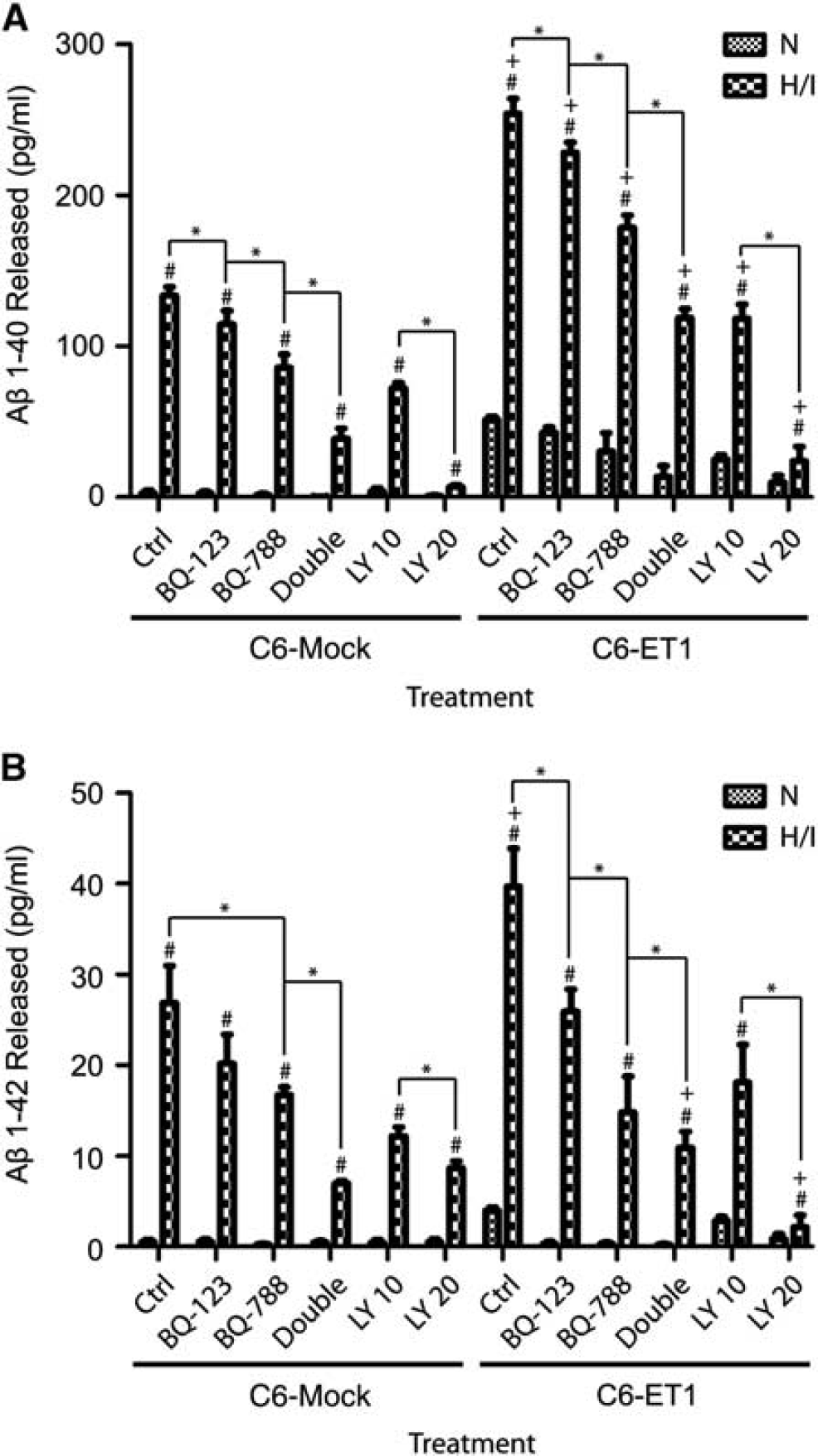
Secretion of (
DISCUSSION
Astrocytes, as a major structural and functional component of neurovascular unit in addition to neurons and endothelial cells, are involved in the regulation of cerebral microcirculation by integrating external stimuli, maintenance of BBB and intrinsic neuronal signals. Under stroke condition, ET-1 is released from both astrocytes and endothelial cells. In the endothelial cells, ET-1 level and ET-1 receptor expressions all increase after stroke. 31 In our previous study, transgenic mice overexpressing endothelin-1 in endothelial cells (TET-1) show more severe brain damage after 2 hours of occlusion. 15 Under 30 minutes of occlusion and 7 days of reperfusion, TET-1 mice show more severe BBB breakdown and increased oxidative stress that may result in neuronal apoptosis and glial reactivity, and contribute to cognitive deficits after short-term ischemia with long-term reperfusion. 16
In Alzheimer's disease, which is characterized to have dementia and memory loss, amyloid
In line with our previous study of a 30-minute tMCAO paradigm, 16 the sensorimotor deficit associated with mild ischemia could be fully recovered in the GET-1 mice as early as 7 days after tMCAO. The imaging results indicated that GET-1 mice showed a severe cerebral edema in the ipsilateral hemisphere at 3 days after tMCAO followed by a severe structural distortion and ventricular enlargement at 1 and 5 months after tMCAO. In addition, with 3-day reperfusion after 30 minutes tMCAO, GET-1 mice exhibited a severe neurodegeneration in the ipsilateral hemisphere where severe cerebral edema was previously observed. These results implicated that astrocytic ET-1 overexpression contributed to progressive neurodegeneration without affecting the normal sensorimotor function of the GET-1 mice.
Cognitive function experiment was performed after a long-term reperfusion (3 months after tMCAO) to examine the occurrence of the poststroke cognitive deficit. The MWM test was used for the cognitive deficit evaluation, as it is a classic method to assess hippocampus-dependent spatial reference learning and memory. 38 In the present study, there was no significant difference in the performance of the visible platform session between different genotypes with or without treatments, indicating that 30 minutes tMCAO followed by 3 months reperfusion did not affect the sensorimotor function of the experimental mice to complete the water maze task. However, in the hidden platform and probe trial sessions, tMCAO-operated GET-1 mice displayed an severe impairment in the spatial reference learning and memory as reflected by an obvious longer escape latency during the hidden platform training and almost without spatial bias (i.e., closed to 25%) during all probe trials. These observations suggested that long-term overexpression of astrocytic ET-1 induced poststroke spatial reference learning and memory deficit without affecting normal sensorimotor function.
Chronic elevation of ET-1 in astrocytes, in which the terminals of these cells surround the endothelium, may exaggerate the neurologic damage of mild ischemia by elevating poststroke vascular stress, since ET-1 is a strong vasoconstrictor. The upregulation of TNF-α and cleaved caspase-3 immunoreactivities in the pyramidal neurons in the ipsilateral hippocampus of the tMCAO-operated GET-1 brain suggested that both neuroinflammation and oxidative stress were induced. Western blot analysis also showed increased expressions of GFAP, HSP70, and Prx6 in the hippocampal lysates. Besides, in the perivascular region of the cerebrovessels in the penumbra region of these mice, cerebrovascular amyloid deposition was evidenced in the region where astrogliosis occurred. These results further supported the hypothesis that astrocytic ET-1 induction has a detrimental role in poststroke cognitive deficit by activating the deleterious response of the hypertrophic astrocytes such as amyloid production and secretion.
The direct effect of astrocytic ET-1 induction on amyloid peptide secretion was investigated. Previous studies have reported that amylogenic A
Impairment of the vascular function by the toxic accumulation of A
Taken together, the present study showed that the possible role of astrocytic ET-1 in contributing to the long-term poststroke cognitive dysfunction after mild ischemia and in the progression of cognitive deficits associated with neurodegenerative diseases such as stroke and AD with the use of targeted astrocytic ET-1 overexpression in transgenic mice and stably transfected ET-1 astrocytes.
Footnotes
The authors declare no conflict of interest.