
Abstract
Glutamate transporters (excitatory amino-acid transporters (EAATs)) are essential for brain homeostasis. While previous studies indicate that the vascular endothelium contributes to glutamate efflux in the adult brain, little information is available regarding glutamate uptake in the immature brain. The present study shows a differential expression pattern of EAATs between cortical microvessels in adults and newborns. In addition, adult cortical endothelial cells take up glutamate more efficiently than neonatal cells. Our findings indicate age-specific changes in extracellular glutamate regulation by brain endothelial cells, suggesting differences in the efficiency of glutamate efflux during an excitotoxic process that, in turn, may contribute to age-specific brain vulnerability.
INTRODUCTION
The amino acid
In the mature cortex, extracellular glutamate levels are regulated by three major high-affinity sodium-dependent excitatory amino-acid transporters, EAAT1–3. EAAT1 and 2 are mainly expressed by glial cells, while EAAT3 is strongly detected in neurons. 1 Accumulating evidence indicates that mature microvascular endothelial cells also express EAAT1–3.5,6 In particular, brain endothelial cells appear to control extracellular glutamate concentrations by inducing the uptake and, subsequently, the efflux of glutamate across the blood-brain barrier.7,8 To our knowledge, EAAT1–3 expression in neonatal endothelial cells has not been studied.
We hypothesized that age-specific differences in CMECs would be associated with differences in their regulation of extracellular glutamate levels. We verified this hypothesis by comparing the expression patterns of EAAT1–3 and glutamate uptake in neonatal and adult cortical endothelial cells and microvessels.
MATERIALS AND METHODS
Animal care and manipulations complied with the recommendations issued by the French and European guidelines for the care and use of laboratory animals (Council Directive 86/609/EEC, license no. 21CAE035).
Animals
Ten-day-old (P10; neonate) or P50 (adult) NMRI mice (Janvier; Le Genest-St-Isle, France) were used. Animals were given food and water
Chemicals and Antibodies
The culture medium Dulbecco's modified Eagle's medium, normal donkey serum, and Alexa Fluor-linked secondary antibodies (A21206; A11055; A21209) were obtained from Invitrogen (Cergy-Pontoise, France). The anti-PECAM antibody (5550274) was from BD Biosciences (Meylan, France). Anti-EAAT1 (sc15316), 2 (sc7760), and 3 (sc52658), and peroxidase-conjugated secondary antibodies (sc2030; sc2031; sc2033) were from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Cell lysis buffer was from Cell Signaling Technology (Boston, MA, USA). The EAAT inhibitor TFB-TBAO was from TOCRIS (Bristol, UK). All other materials used, including the anti-β-actin antibody (A5441) and glutamate, were from Sigma-Aldrich (Saint-Quentin Fallavier, France).
Preparation of Brain Sections
Neonatal and adult mice were anesthetized with isoflurane (inhaled) and pentobarbital (intramuscular), respectively, and perfused transcardially with paraformaldehyde (4% in PBS; pH 7.4; 4°C). Brains were immersed overnight in the same fixative (4°C) and stored in 25% sucrose/PBS solution for 48 hours before freezing. Coronal sections (40 μm) were cut on a cryostat (Leica, Rueil-Malmaison, France).
Cortical Microvessel Isolation
Cortical microvessels from neonate and adult mice were prepared as previously described. 9 Sample were frozen (−;80 °C) until western blot analysis or resuspended in cold paraformaldehyde (4% in PBS; pH 7.4), spread on slides overnight at 4°C, rinsed twice with PBS, and frozen at −;20°C for immunocytochemistry.
Primary Cultures of Cortical Microvessel Endothelial Cells
As previously described, 4 freshly isolated microvessels were digested with collagenase/dispase and DNAse, erythrocytes were lysed, and then microvessels were seeded onto 24-well plates. Presence of astrocytes in both microvessels and CMECs was controlled by western blot and immunolabeling against GFAP (Supplementary Figure 1). When CMECs reached confluence (3–5 DIV), cells were used for western blot analysis or exposed up to 60 minutes to 500 μL of Dulbecco's modified Eagle's medium containing glutamate (50 μmol/L) in the presence or absence of the EAAT inhibitor, TFB-TBAO (600 nmol/L). Media were then collected from 5 to 60 minutes and frozen at −80 °C for glutamate concentration assays.
Immunolabeling
Samples were incubated overnight at 4°C with antibodies to PECAM, EAAT1, EAAT2 or EAAT3 in PBS containing 0.1% normal donkey serum and 3% Triton X-100. Tissues were incubated for 2 hours with the corresponding Alexa Fluor-linked secondary antibody. Cell nuclei were visualized with 1 μg/mL Hoechst 33258. Fluorescent labeling of isolated microvessels and brain sections were observed under a Leica DMI-600B fluorescence and a TCS-SP2 confocal microscope, respectively. The specificity of immunolabeling was assessed by omitting the primary antibody.
Western Blot Analysis
Proteins from cortical microvessels or CMECs extracts were loaded onto 10% SDS-polyacrylamide gels and electroblotted onto nitrocellulose membranes. Membranes were incubated overnight (4°C) with primary antibodies to β-actin, EAAT1, EAAT2, or EAAT3. After incubation with the corresponding peroxidase-coupled secondary antibodies, proteins were visualized using the ECL Plus immunoblotting detection kit (Amersham, UK). The intensity of immunoreactive bands was quantified using a blot analysis system (BioRad Laboratories, Marnes-la-coquette, France) and data expressed as the ratio of protein to β-actin.
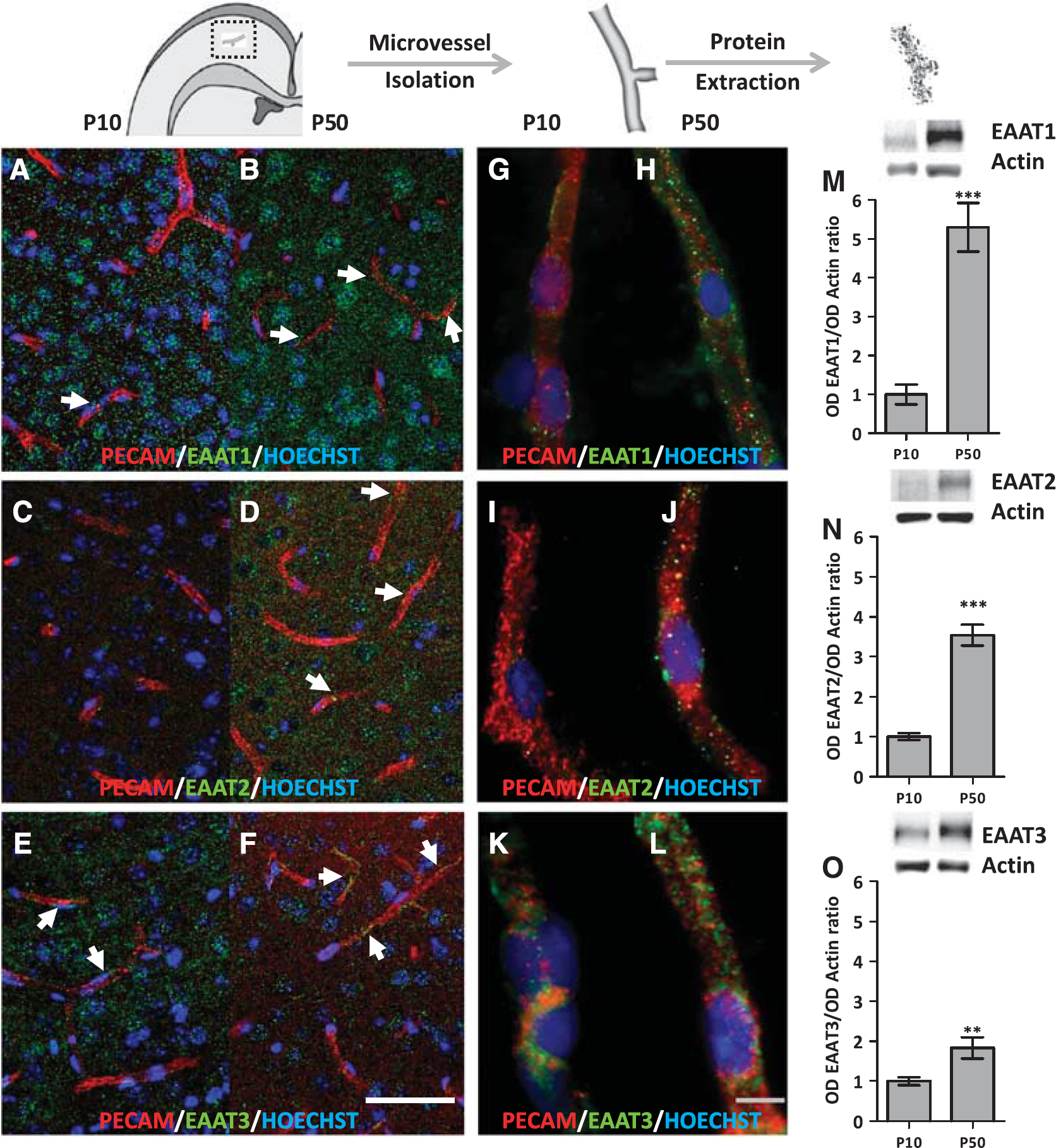
Expression of excitatory amino-acid transporter (EAAT) 1–3 in cortical microvessels derived from neonatal (P10) and adult (P50) mice. (
Glutamate Concentration Assay
The glutamate concentration of media from CMECs was quantified following the instructions provided with the Glutamate Assay Kit (Biovision, Milpitas, CA, USA). Absorbance was measured at 450 nm using a Chameleon plate reader (Mustionkatu, Turku, Finland).
Statistical Analysis
Data are expressed as means ± s.e.m. Relative optical densities from immunoblots were compared using an unpaired
RESULTS
Expression Pattern of EAATs in Neonate and Adult Cortical Microvessels
Using an antibody against the endothelial cell marker PECAM, a double-immunolabeling approach was used to visualize the expression of EAATs in cortical vessels from P10 and P50 brain slices (Figures 1A–F). At P10, numerous EAAT1-positive cells were seen in the neocortex (Figure 1A). Few puncta were colocalized with PECAM-positive microvessels (Figure 1A, arrow), indicating that most of the EAAT1 labeling was associated with non-endothelial, probably neural cells (Figure 1A). At P50, the density of EAAT1 puncta was clearly higher than at P10. This increase concerned both microvessels and non-endothelial cells (Figure 1B, arrows). Although EAAT2 immunolabeling was very low at P10, a similar increase in puncta density was observed in microvessels and non-endothelial cells at P50 (Figures 1C and 1D). In contrast, the density of EAAT3-labeled puncta decreased between P10 and P50 in the parenchyma, whereas there did not appear to be a marked difference in microvessels (Figures 1E and 1F, arrows).
Because EAAT immunoreactivity was observed in both endothelial and non-endothelial cells, we performed immunocytochemistry (Figures 1G–L) and western blotting experiments (Figures 1M–O) with isolated cortical microvessels. At P10, immunoreactivity for EAAT1 and 2 was quite low (Figures 1G and II) whereas EAAT3 was still detectable (Figure 1K). At P50, immunolabeling for all three EAATs was observed (Figures 1H, 1J, and 1L). EAAT labeling was thus more pronounced in adult than in neonatal microvessels (Figures 1G–L). The expression levels of EAATs in isolated microvessels at P10 and P50 were quantified by western blotting (Figures 1M–O). When compared with neonates, adult microvessels exhibited a 5.3-, 3.5-, and 1.8-fold increase in EAAT1, EAAT2, and EAAT3 protein levels, respectively (
Quantification of Glutamate Uptake by Neonate and Adult CMECs
Because immunocytochemistry and western blotting revealed age-specific expression patterns of the EAATs in cortex-enriched microvessels, we hypothesized that nCMECs and aCMECs could differ in terms of glutamate uptake. First, we controlled that age-dependent specificities persisted in cultured CMECs (Figures 2A–D). When compared with nCMEC, aCMEC did not exhibit significant differences of EAAT1 protein levels. In contrast, aCMEC presented a 1.7- and 1.9-fold increase in EAAT2 and EAAT3 protein levels, respectively (
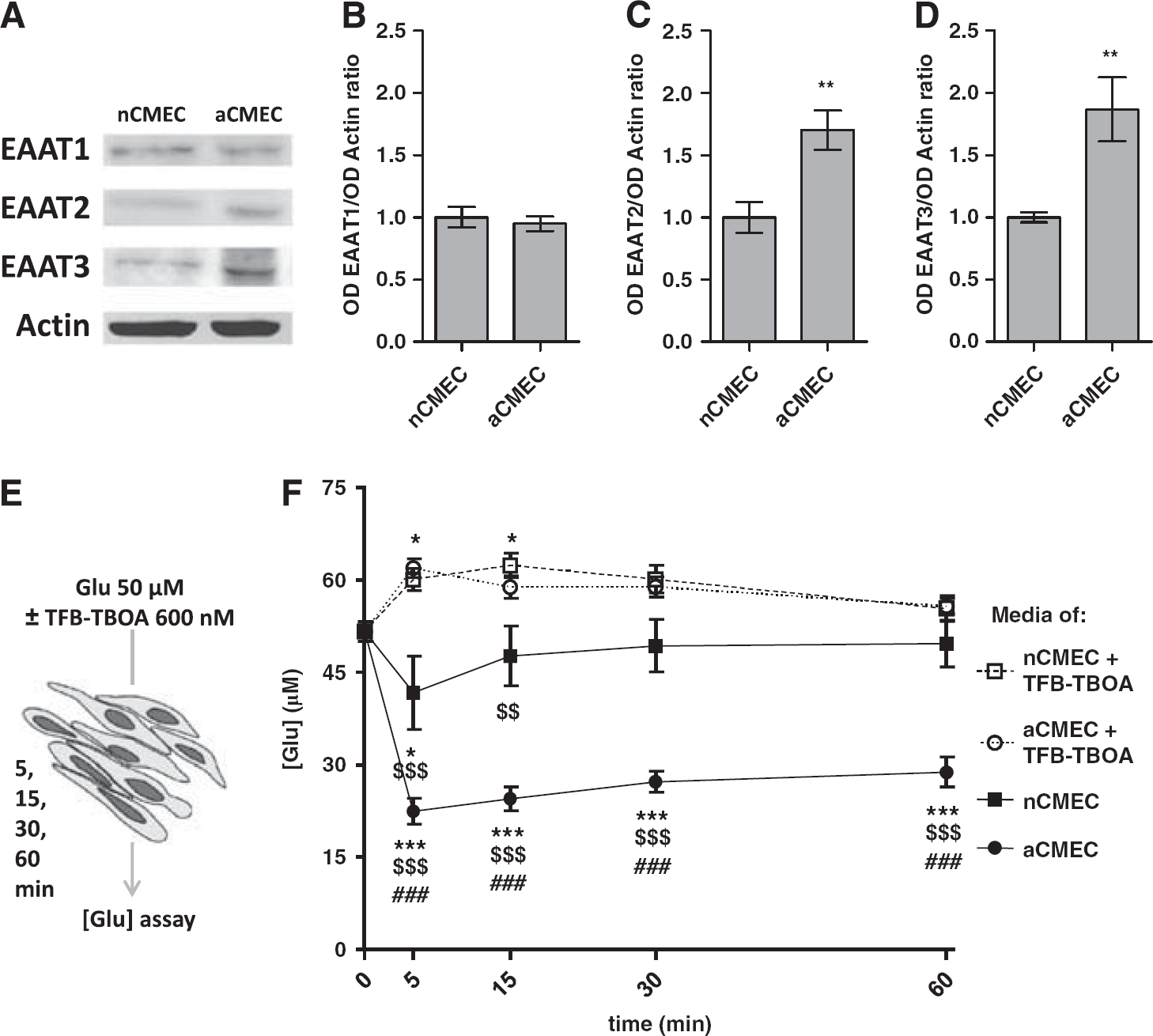
Quantification of glutamate uptake by neonatal and adult CMECs. (
DISCUSSION
The present study revealed (i) different expression patterns of EAATs between neonate and adult cortical microvessels and cultured CMECs, and (ii) improved EAAT-mediated uptake of glutamate in aCMECs. Taken together, these data suggest that the microvasculature of the immature cortex presents different functional characteristics from those in the mature brain, in particular regarding glutamate uptake that may impact the brain-to-blood efflux of glutamate.5–8 These results are in line with the higher sensitivity of nCMECs to glutamate. 4 Indeed, nCMECs express more NMDAR and are more prone to release vascular protease under glutamate stimulation than aCMECs. 4 Together with the present data, this suggests that the low ability of nCMECs to take up glutamate may facilitate these functional differences.4,9
Previous ontogenic studies have shown an increase in the expression of the glial transporters EAAT1 and EAAT2 during cortical development,10,11 while, contrasting with microvessels, expression levels of the predominantly neuronal transporter EAAT3 are higher in the immature cortex. 10 Moreover, the lack of EAAT1 and EAAT2 results in morbidity associated with developmental abnormalities such as impaired neuronal migration, indicating the essential role of EAATs in brain development. 12 Glutamate also has age-specific trophic effects in the developing cortex. 3 Although EAATs are not the only glutamate transporters expressed by CMECs, 6 the differential expression patterns of the EAATs in microvessels suggest that glutamate effects in the immature cortex are not subject to the same regulatory processes as in adulthood regarding brain-to-blood efflux. In particular, the molecular and functional specificities of mature versus immature endothelial cells with respect to EAAT expression and glutamate uptake support specific glutamate homeostasis pathways that are essential for cortical development.
Besides the trophic effects of glutamate during cortical development, excitotoxicity has been pinpointed as a major process in the complex setting of neonate brain lesions.3,13 Excess glutamate contributes to blood-brain barrier breakdown and associated neurodegeneration at the neurovascular unit level. 14 Thus, low brain-to-blood glutamate efflux in the immature brain could contribute to the high vulnerability of blood-brain barrier and neurons of the developing cortex to excitotoxic insults. 9 In addition, the present data suggest that promising therapeutic strategies characterized in adult models of stroke and head injuries, based on increasing glutamate efflux by enhancing the brain-to-blood gradient, may not be as effective in neonates.5,15
In conclusion, our data provide new evidence of the molecular and functional specificities of immature endothelial cells with respect to EAAT expression and glutamate uptake. They reinforce the hypothesis of vascular involvement in both the developmental and pathologic effects of glutamate in the immature brain. Finally, they justify the investigation of vessel-oriented neuroprotective strategies in neonates.
Footnotes
The authors declare no conflict of interest.
ACKNOWLEDGMENTS
We thank Y Ramdani for her technical assistance with confocal microscopy, E Mitard for schematic illustrations, and S Rasika for English editing.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.