
Abstract
Cortical spreading depression (CSD) involves mass depolarization of neurons and glial cells accompanied with changes in regional cerebral blood flow (rCBF) and energy metabolism. To further understand the mechanisms of CBF response, we examined the temporal diametric changes in pial arteries, pial veins, and cortical capillaries. In urethane-anesthetized mice, the diameters of these vessels were measured while simultaneously recording rCBF with a laser Doppler flowmeter. We observed a considerable increase in rCBF during depolarization in CSD induced by application of KCl, accompanied by a transient dip of rCBF with marked vasoconstriction of pial arteries, which resembled the response to pin-prick-induced CSD. Arterial constriction diminished or disappeared during the second and third passages of CSD, whereas the rCBF increase was maintained without a transient dip. Long-lasting oligemia with a decrease in the reciprocal of mean transit time of injected dye and mild constriction of pial arteries was observed after several passages of the CSD wave. These results indicate that CSD-induced rCBF changes consist of initial hyperemia with a transient dip and followed by a long-lasting oligemia, partially corresponding to the diametric changes of pial arteries, and further suggest that vessels other than pial arteries, such as intracortical vessels, are involved.
Keywords
Introduction
Cortical spreading depression (CSD) is a mass depolarization of neurons and glial cells, followed by sustained suppression of spontaneous neuronal activity. It slowly propagates in gray matter regardless of functional divisions or the vascular network, 1 and is thought to be involved in triggering headache because of the similarity between the electrophysiologic characteristics of CSD and the symptoms of migraine aura. 2 Cortical spreading depression-like depolarization waves, known as peri-infarct depolarizations, have been detected with high frequency in aneurismal subarachnoid hemorrhage, delayed ischemic stroke after subarachnoid hemorrhage, malignant hemispheric stroke, spontaneous intracerebral hemorrhage, and traumatic brain injury. These depolarizations often induce profound metabolic and hemodynamic effects, resulting in exacerbation of the ischemic tissue outcome. 3
Cortical spreading depression is accompanied with various changes in blood flow and energy metabolism.4, 5 It induces remarkable increases in glucose utilization and metabolism in the cerebral cortex, 6 and the cerebral metabolic rate of oxygen shows a prolonged increase. 7 This elevation of oxygen consumption often leads to tissue hypoxia, 8 and changes in partial tissue pressure of oxygen (PO2) elicited by CSD are closely linked to cerebral blood flow (CBF) changes. 9
Cortical spreading depression has been experimentally elicited with a variety of chemical, electrical, and mechanical stimuli in various animals, and research in animal models has provided much information about its pathologic role and also clues to potential clinical therapies. The response of CBF to CSD involves the production and release of multiple vasodilating and vasoconstricting factors by diverse cells within the neurovascular unit. 10 In mice, CSD causes severe vasoconstriction, resulting in CBF and oxygenation decreases to ischemic level.11, 12, 13 The CSD-induced hyperemia is followed by a long-lasting reduction to below the prestimulus level, so-called post-CSD oligemia.14, 15, 16 At least three vasomotor elements, vasoconstrictive tone that temporally overlaps with the depolarization, profound hyperemia that starts at or soon after the onset of depolarization and long-lasting hypoemia, were proposed by Ayata. 4 He suggested that the magnitude and timing of each vasomotor element seemed to be dependent on species, systemic and physiologic state, pharmacological state, and other factors.
We have reported that potassium-induced CSD elicited a biphasic response of regional CBF (rCBF) in anesthetized animals, namely initial oligemia with capillary flow cessation followed by hyperemia,17, 18 and initial constriction followed by propagated dilation of pial arteries. 19 Furthermore, we showed heterogeneous changes of red blood cell (RBC) velocity in intraparenchymal capillaries, namely a sustained decrease and a remarkable increase, while rCBF was elevated. 20 The suppression of RBC velocity was due to mass depolarization of neuronal and glial cells. 21 In this work, to elucidate the mechanisms controlling the changes of microcirculation associated with CSD, we examined the behavior of various types of surface vessels, including pial arteries (arterioles) and veins (venules) and cortical capillaries, by means of laser-scanning confocal fluorescence microscopy.
Materials and methods
General Procedures
Animals were used with the approval (No. 09058) of the Animal Ethics Committee of Keio University (Tokyo, Japan), and all experimental procedures were in accordance with the university's guidelines and the ARRIVE (Animal Research: Reporting In Vivo Experiments) reporting guidelines for the care and use of laboratory animals. Both sexes of Tie2-green fluorescent protein (GFP) transgenic mice (STOCK Tg [TIE2GFP] 287Sato/J; Jackson Lab, Bar Harbor, ME, USA) (12 to 24 weeks old, weighing 30.5±4.1 g, N=30) were used. Tie 2, a receptor of angiopoietins 1 and 2, is expressed specifically in endothelial cells, and fluorescent endothelial cells in Tie2-GFP transgenic mice can be specifically identified by means of fluorescence microscopy or confocal laser microscopy. 22
Under urethane anesthesia (1.1 g/kg of body weight, intraperitoneally), each mouse was fixed to a head-holder (SG-4N modified to be flexible around the horizontal axis; Narishige Scientific Instrument Laboratory, Tokyo, Japan) and was intratracheally intubated to provide artificial ventilation with a small animal ventilator (SAR-830/AP; CWE, Inc., Ardmore, PA, USA) and to monitor endtidal CO2 with a CO2 analyzer (microCapStar; CWE, Inc.). Endtidal CO2 level was maintained constant by adjusting the ventilation rate. Arterial blood pressure (ABP) was continuously recorded through a femoral arterial catheter via a surgical strain-gage (MLT0670 and ML117, ADInstruments Pty. Ltd., Bella Vista, NSW, Australia) and heart rate (HR) was determined from the ABP wave. Mean arterial blood pressure (MABP) was maintained within ±20 mm Hg in each mouse throughout the experiment. The data of a mouse in which MABP decreased below 50 mm Hg was excluded (N=1). Body temperature was maintained at around 37°C with a heating-pad and thermo-controller (BWT-100, Bioresearch Center Co., Ltd., Nagoya, Japan). Arterial blood gas analysis was performed with a blood gas analyzer (RapidLab 348, Siemens AG, Munich, Germany) before and after induction of CSD.
A cranial window of approximately 3 mm diameter was made at the parieto-temporal region of the left, right, or both sides of the cerebral cortex, and sealed with a circular 140-μm-thick quartz glass fixed with dental cement over the intact dura to prevent dehydration. 23 A posterior hole, approximately 1 mm in diameter with a small incision of the dura, centered at the coordinates of 5 mm posterior and 2 mm lateral to the bregma, was made. After confirmation that no CSD episode had occurred for at least 30 minutes and all parameters had remained stable for at least 10 minutes, CSD was induced by (1) chemical stimulation with KCl solution (1.0 mol/L, 5 μL) application (N=16) or (2) mechanical stimulation (pin prick) by inserting a 30G needle (outer diameter 0.3 mm) approximately 1 mm (N=14). The CSD induction was performed only once in one hemisphere, because hemodynamic and metabolic changes associated with second CSD may be different from those of first CSD. 24 It is well established that CSD does not propagate from one hemisphere to the other. Therefore, CSD was induced at both hemispheres in some mice.
Analysis of Diameter of Various Vessels
Vessels were observed through the cranial window with a confocal microscope equipped with an argon laser (λ=488 nm) (532-BS-A04; CVI Melles Griot, Carlsbad, CA, USA), a multibeam confocal scanning unit (CSD-22, Yokogawa Electric Co., Tokyo, Japan), a CCD camera (ImagEM, Hamamatsu Photonics, Hamamatsu, Japan) and a × 20 water immersion objective lens (CFI Fluor × 20 NA=0.50; Nikon Co., Tokyo, Japan). Confocal images consisting of 512 × 512 pixels with the in-plane pixel size of 0.75 μm were captured every 5 seconds from just after KCl application for 5 minutes, followed by every 10 seconds. The gain of confocal images and the exposure time (1.5 to 2.0 seconds) were individually adjusted and fixed in each mouse. The sequential images were collected and saved as an AVI file with LuminaVision software (Mitani Co., Tokyo, Japan). The brain surface (depth zero) was set at the level at which the focused pial arteries appeared largest, and images were captured to the depth of 20 to 30 μm. Therefore, we could recognize pial arteries and pial veins in all animals, and also cortical capillaries in some animals within the region of interest (ROI). The diameter of microvessels (pial arteries, pial veins, and cortical capillaries) was evaluated with the image analysis software, ImagePro (Media Cybernetics Inc., Rockville, MD, USA). The diameter of the vessels was evaluated as the width of identical GFP-indicating endothelium. Plural vessels of different branches were evaluated in each mouse.
Measurement of Direct Current Potential and Regional Cerebral Blood Flow
A direct current (DC) potential electrode (EEG-5002Ag, Bioresearch Center Co., Ltd.) and the probe (BF52, Advance Co., Ltd., Tokyo, Japan) of a laser Doppler flowmeter (LDF) (ALF 21R, Advance Co., Ltd.) were positioned on the intact dura and on the skull, respectively, to avoid possible mechanical induction of CSD by incision of the dura. The wavelength of LDF was 780 nm, which did not interfere with the laser excitation or detection of the fluorescence signal. The DC potential was applied at 1 to 100 Hz and digitized at 1 kHz with a differential headstage and differential extracellular amplifier (Model 4002 and EX1; Dagan Co., Minneapolis, MN, USA). Continuous recordings of ABP as well as HR, rCBF, and DC potential were stored on a multichannel recorder (PowerLab 8/30; ADInstruments Pty. Ltd.) and LabChart software (ADInstruments Pty. Ltd.) was used for off-line analysis as described previously. 20 A schematic of the cranial window and the positions of the DC potential electrode, the LDF probe and additional small window for KCl application are shown in Figure 1A.
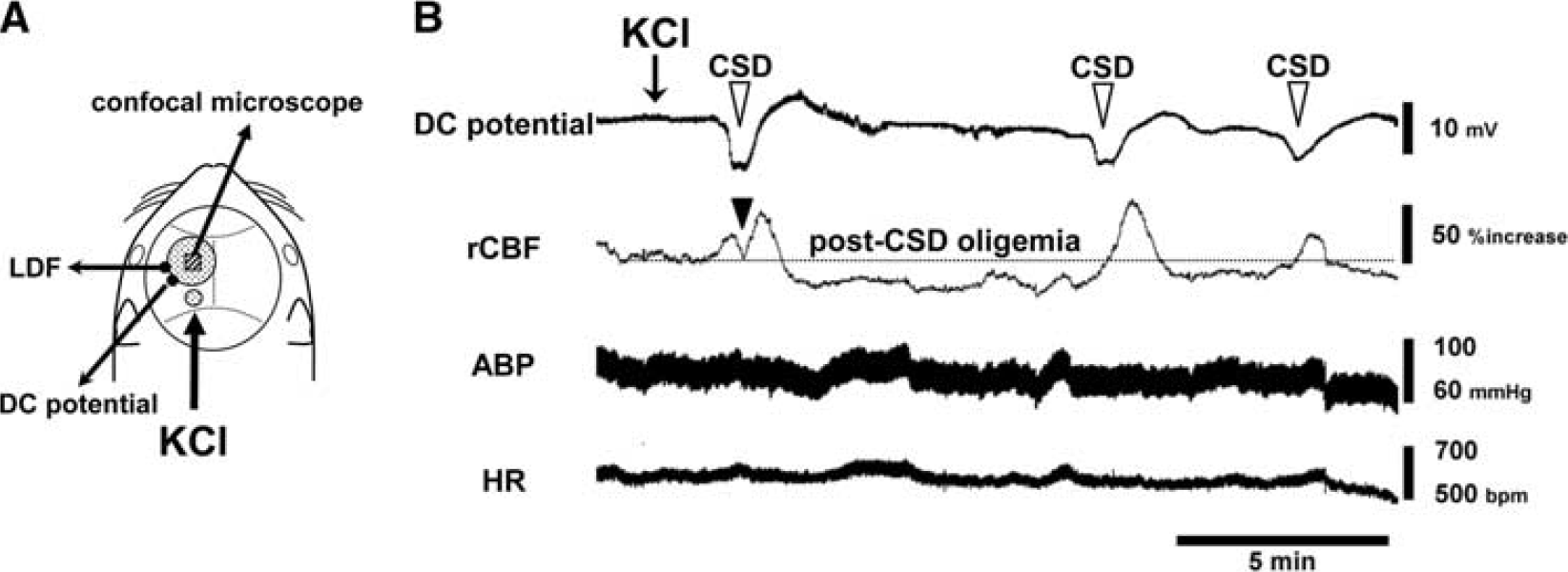
Experimental setup and typical recordings. (
Cortical microflow was also evaluated using a modified version 25 of our original software KEIO-IS1 26 based on a dye-dilution method before and 15 minutes after the last CSD passage. A 50-μL aliquot of rhodamine B isothiocyanate-dextran 70 (RITC-dextran; 8 mg/mL; Sigma-Aldrich Co., St Louis, MO, USA) was injected into the tail vein, and 100 frames of fluorescence images excited with an Hg lamp (LH-M100CB-1; Nikon Co.) equipped with a band-pass filter (G2A, Nikon Co.) were captured at a rate of 10 frames/s and saved as an AVI file with LuminaVision. Using our original software KEIO-IS1 with the aid of Matlab software (The Mathworks, Inc., Natick, MA, USA), ROI was divided into 50 × 50 matrices, creating 2,500 dilution curves, and the mean transit time (MTT) of the dye was calculated in each matrix. After reconstruction of the 2D-flow map, the reciprocal of MTT (1/MTT) was calculated as an index of cortical microflow in parenchyma without any large vessels.
Data Analysis
Changes of rCBF and vessel diameter were indicated as a percentage (%) and change of DC potential was calculated by subtracting the averaged level for 30 to 60 seconds before the onset of the DC potential deflection of each CSD. Direct current potential, rCBF, and diametric response of various vessels were temporally averaged every 5 seconds or 10 seconds. A criterion time was set at the transient dip in rCBF for the evaluation of rCBF and DC potential, and at the transient constriction in arterial diameter for the evaluation of vessel diameter during the first CSD, because these transient changes were very rapid and were observed in all animals. During the second and third CSD, the criterion time was set at the onset of deflection of DC potential. Maximum changes of rCBF were obtained by averaging for 10 seconds around the peak of the response curve. Maximum changes of pial arteries (other than transient constriction) were obtained by averaging 3 to 6 frames around the trough or peak of the response curve to eliminate random fluctuations, because these changes were much smaller and broader. Maximum changes of pial veins and capillaries were obtained as the averages of 3 to 6 frames at the same timing as for pial arteries.
All data are reported as means±s.d. Changes of rCBF and vessel diameter were analyzed using a paired t-test versus the pre-CSD level. The correlation between basal diameter and % change was analyzed and Pearson's product moment correlation coefficient was obtained from Pearson's correlation and least-squares regression analysis. A P-value of <0.05 was considered to be statistically significant.
Results
General Results
Physiologic parameters before and after CSD induction by KCl application or pin prick are shown in Table 1. These parameters did not significantly change during the experiment.
Physiologic parameters before and after CSD induction by KCl application or pin prick
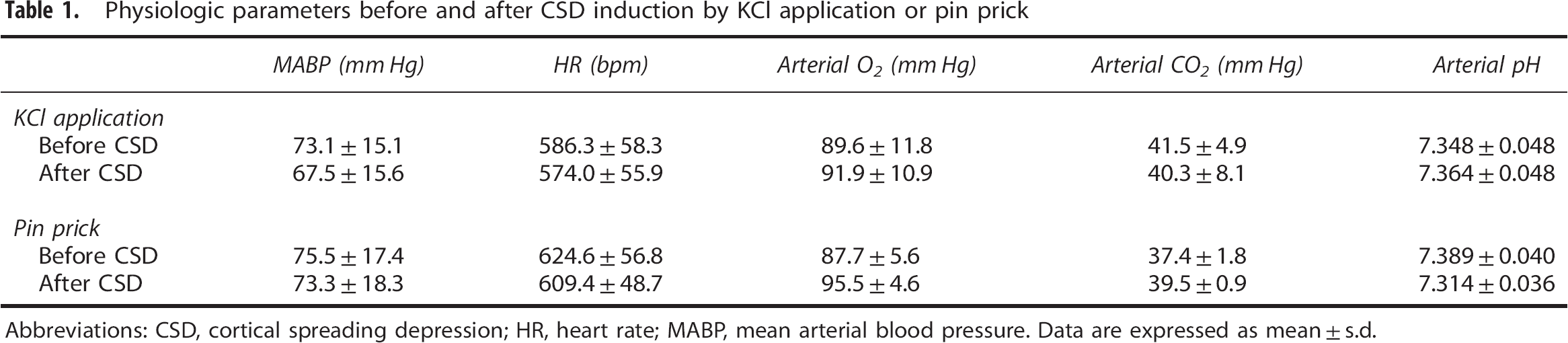
Abbreviations: CSD, cortical spreading depression; HR, heart rate; MABP, mean arterial blood pressure.
Data are expressed as mean±s.d.
As shown in Figure 1B, one application of KCl induced repetitive negative deflections of DC potential (i.e., CSD) accompanied with rCBF increase with an interval of 5 to 10 minutes. Regional CBF response was accompanied with a transient dip during the first CSD, and rCBF declined below the preapplication level after CSD induction (post-CSD oligemia), whereas response of ABP and HR specifically associated with CSD occurrence was not seen.
Changes in Diameter of Various Types of Vessel
Pial arteries and veins could be discriminated because the arteries showed dense and thick GFP-expressing endothelial cells, whereas the venous endothelium showed sparse and spot-like distributions. Capillaries were distinguished on the basis of a diameter of <10 μm and a uniform distribution of GFP. Approximately 1 minute after KCl application, pial arteries extremely constricted, concomitantly with a transient decrease of rCBF, followed by dilation and long-lasting slight constriction during oligemia. However, pial veins and cortical capillaries showed smaller changes (Figure 2; Supplementary Video 1).
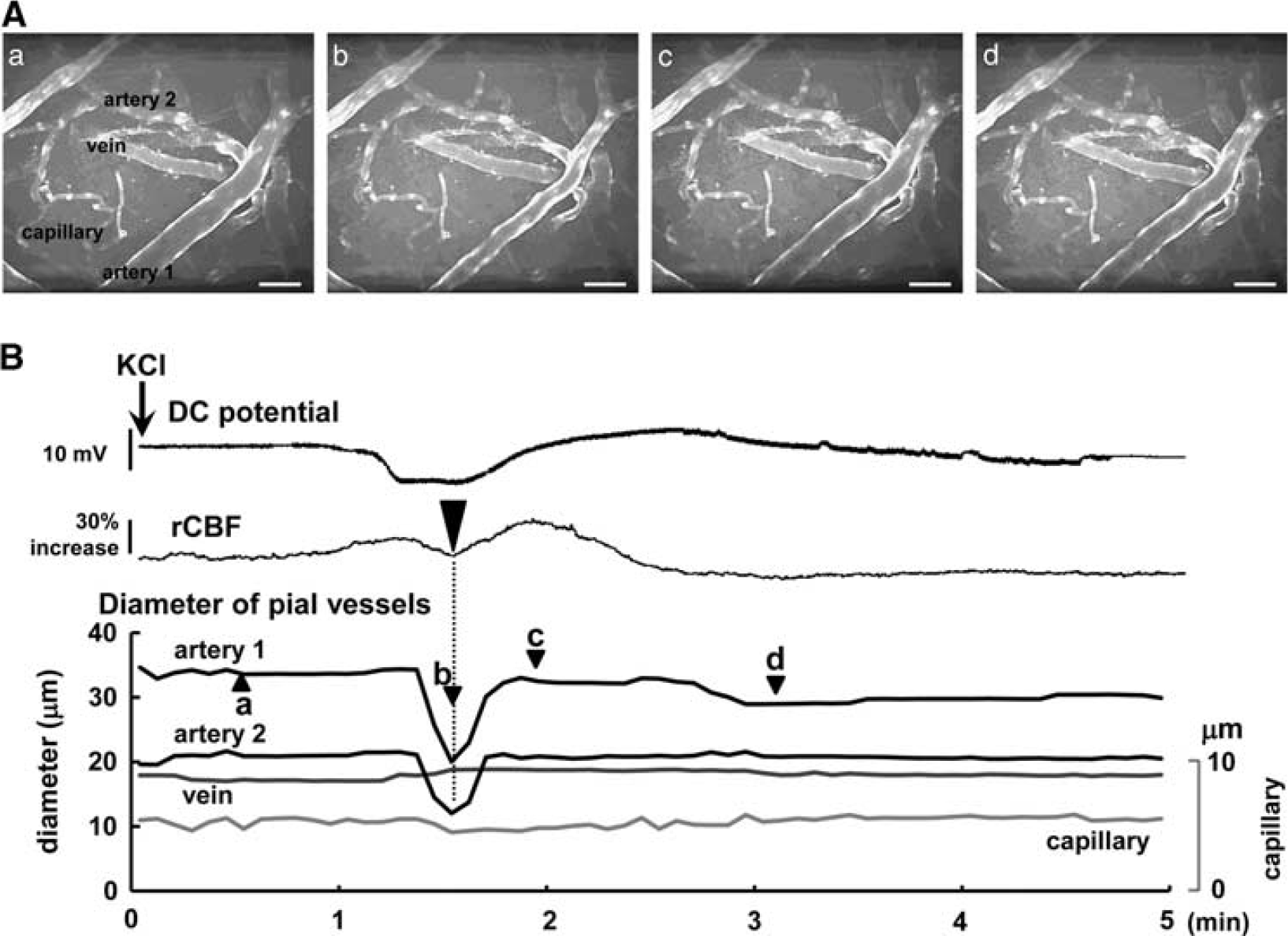
Typical images of the brain surface under the cranial window, and evaluation of vessel diameter. Confocal laser-scanning images (
Temporal changes of diameter of various vessels are shown along with the changes of DC potential and rCBF in Figure 3. KCl application elicited an rCBF increase together with a DC potential deflection. During the phase of hyperperfusion associated with the first CSD, a transient dip of rCBF was observed in all mice (N=15). All pial arteries in the ROI markedly constricted at approximately the same time as the transient dip of rCBF, with a time difference of 9.6±13.6 seconds, followed by slight dilation and long-lasting constriction (Figure 3A (b) and (c)). Decreased arterial diameter returned to baseline or reached a peak under the baseline after 32.1±10.7 seconds from the beginning of constriction. Regional CBF and diameter of the arteries generally began to increase just before the onset of DC potential deflection. However, the second CSD elicited a quite small and broad constriction, followed by a mild and relatively long-lasting dilation (Figure 3B (c)). Further smaller constriction with no apparent dilation was observed during the third CSD (Figure 3C (c)). As shown in Figures 3B and 3C, whether constriction was observed or not, and despite the mild dilation, a prominent increase of rCBF was elicited by the second and third CSD. No transient dip of rCBF was observed during the second or third CSD. A smaller change of arterial diameter with a prominent increase of rCBF was observed during the fourth or later CSD passage in some mice (data not shown).
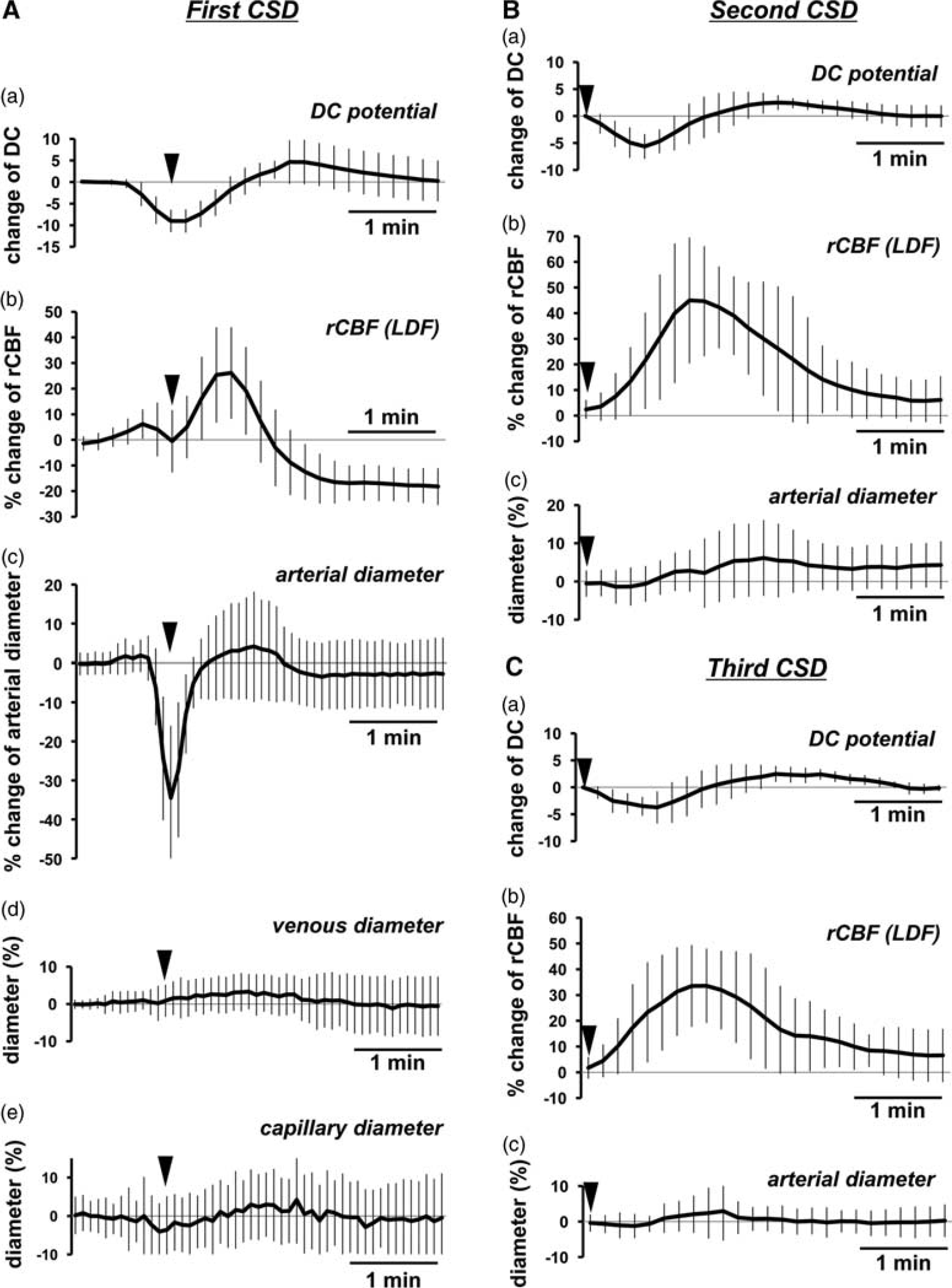
Time course of changes of DC potential, rCBF, and vessel diameter associated with repeated CSD induced by KCl application. Average response of DC potential (a), rCBF measured with LDF (b) and diameters of arteries (c), veins (d), and capillaries (e) associated with first CSD (
Temporal changes of diameter of veins and capillaries in response to the first CSD passage are shown in Figure 3A (d) and (e). Diameter of measured veins (29.6±16.7 μm) before KCl application was identical to that of arteries (26.6±9.0 μm). Transient constriction of veins was not observed, but there was a mild and long-lasting dilation during the first CSD. Slight constriction in capillaries was observed coincident with the arterial vasoconstriction phase during the first CSD, but the changes in diameter were small and heterogeneous.
Peak changes of vessels and rCBF increases are summarized in Figure 4. Initial transient constriction and subsequent dilation were seen in pial arteries, and the responses were diminished with repeated CSD passage. Mild dilation and mild constriction were seen in veins and capillaries, respectively. These responses were also diminished with repeated CSD passage. However, marked increase of rCBF was repeatedly observed as shown in Figure 4 (b).
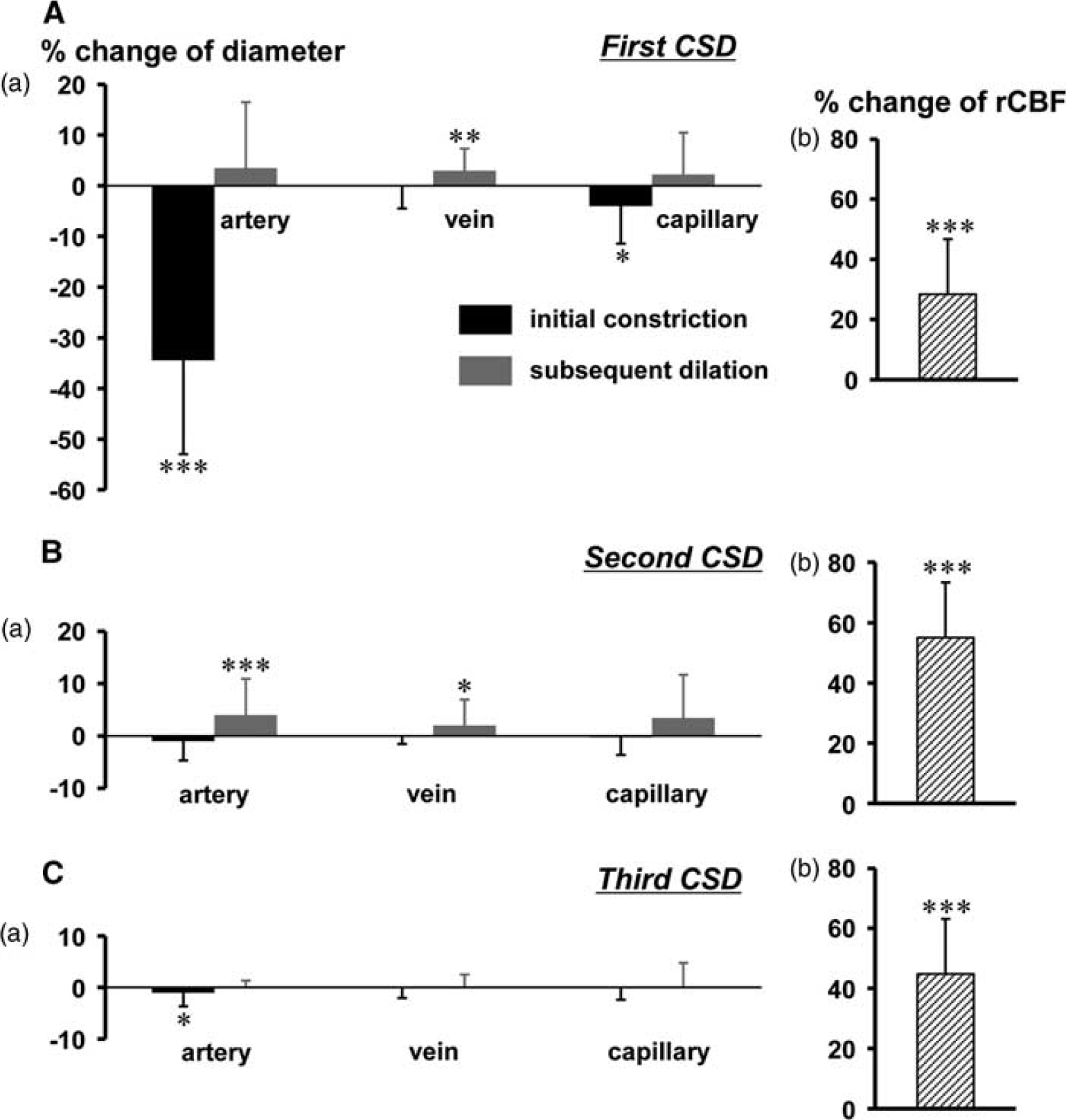
Summary of the diametric changes of various vessels (a) and rCBF increase during the subsequent dilation phase (b) associated with CSD. Peak changes of diameter (a) of pial arteries, pial veins, and cortical capillaries elicited by first (
Arterial Vasoreactivity to Cortical Spreading Depression
Large arteries of more than 40 μm in diameter showed rather small constriction in response to the first CSD. There were low but significant correlations between basal diameter and % change in diameter in the initial transient constriction (R=0.342, P<0.05) and between basal diameter and subsequent dilation (R=−0.339, P<0.05) (n=42 in 14 mice) (Figure 5).
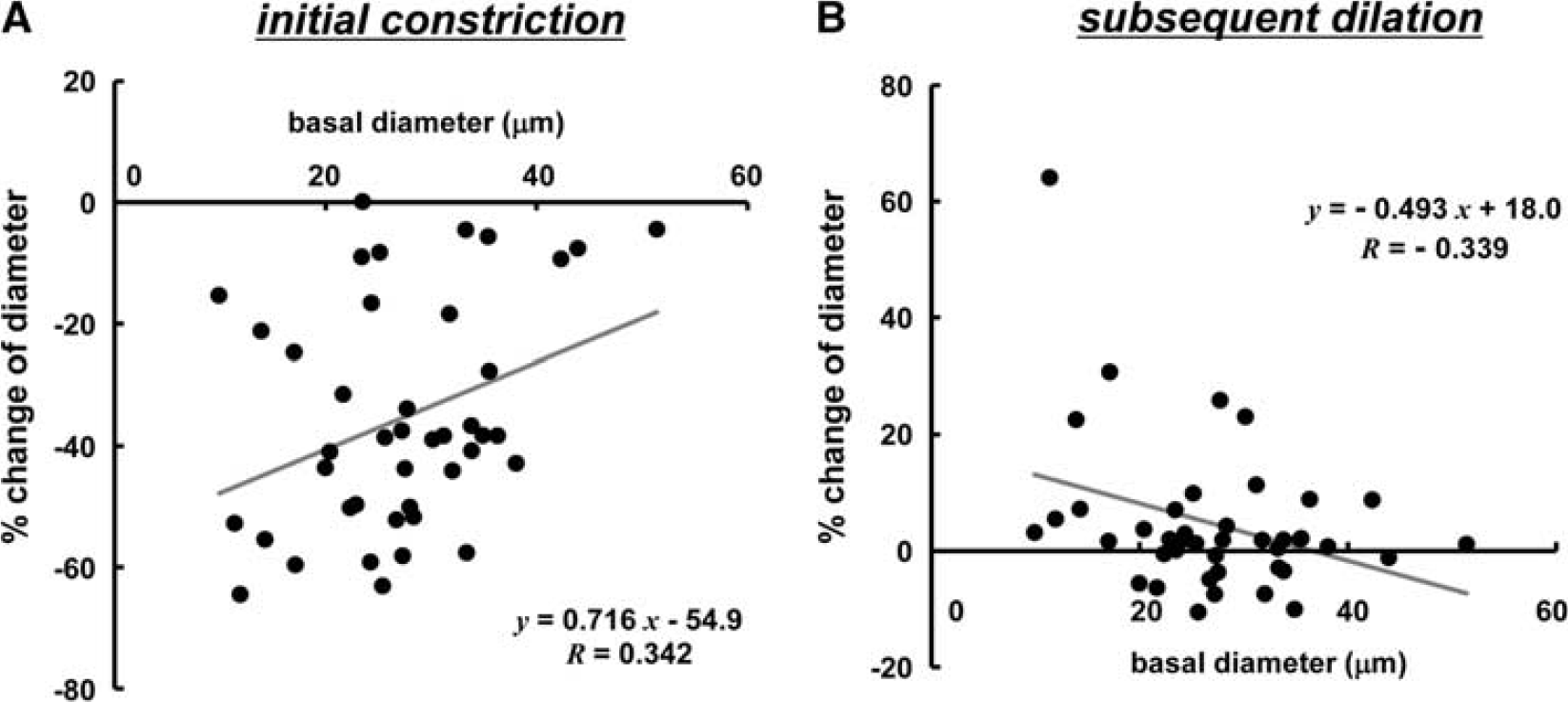
Correlation of changes of pial arteries with basal diameter. Scatter diagram and regression line of initial vasoconstriction (
To rule out the possibility that the arterial reaction might be due to potassium diffusion to the recording area, arterial reactivity in response to pin prick-induced CSD was examined along with DC potential and rCBF measurements. We found that CSD was induced by pin prick in 11 cases in 26 trials (42.3%) in 14 mice. Parameters were temporally averaged in 10 cases in which confocal images were successfully captured, as shown in Figure 6. When CSD was elicited, more pronounced constriction was evoked than in the case of KCl application (P<0.01). Subsequent dilation was milder and four arteries did not return to the baseline level (Figure 6C). A profound transient dip and mild subsequent increase of rCBF were seen (Figure 6B).
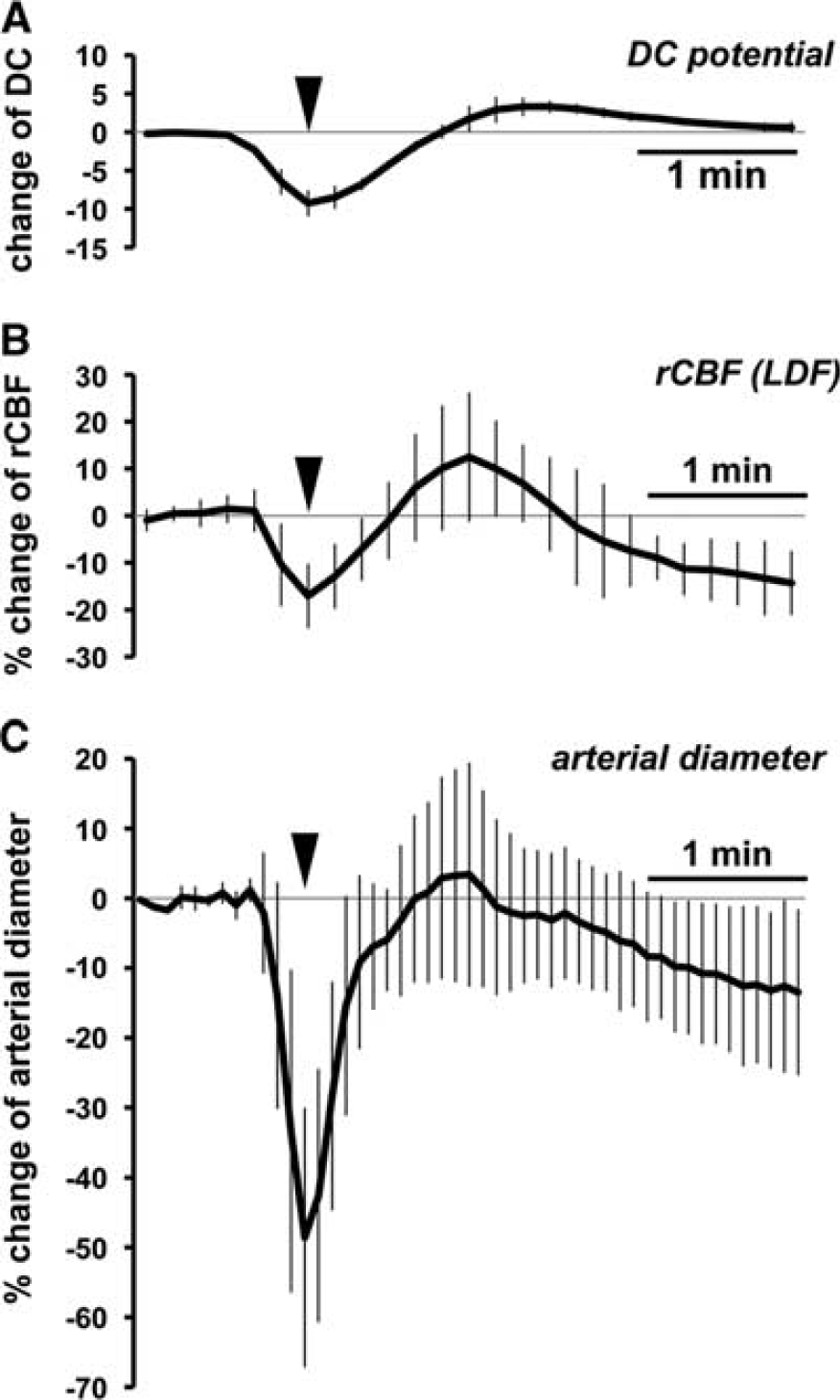
Time course of changes of DC potential (
Cortical Microflow and Vessel Diameter
After several CSD episodes, rCBF declined persistently below the preKCl level (post-CSD oligemia). We evaluated 1/MTT as an index of cortical microflow using a dye-dilution method, KEIO-IS1 (Figure 7A; Supplementary Video 2) and compared the results with LDF-evaluated rCBF and vessel diameter. As shown in Figure 7B, 1/MTT was approximately 63% of the LDF-evaluated rCBF. Changes of 1/MTT were significantly correlated with the LDF-evaluated changes of rCBF (R=0.721, P<0.05) (Figure 7C). Pial arteries showed slight constriction, whereas the diameter of veins and capillaries did not show any significant change (Figure 7D). Changes of arterial diameter were not significantly correlated with the changes of rCBF (Figure 7E) (R=0.027, P>0.1).
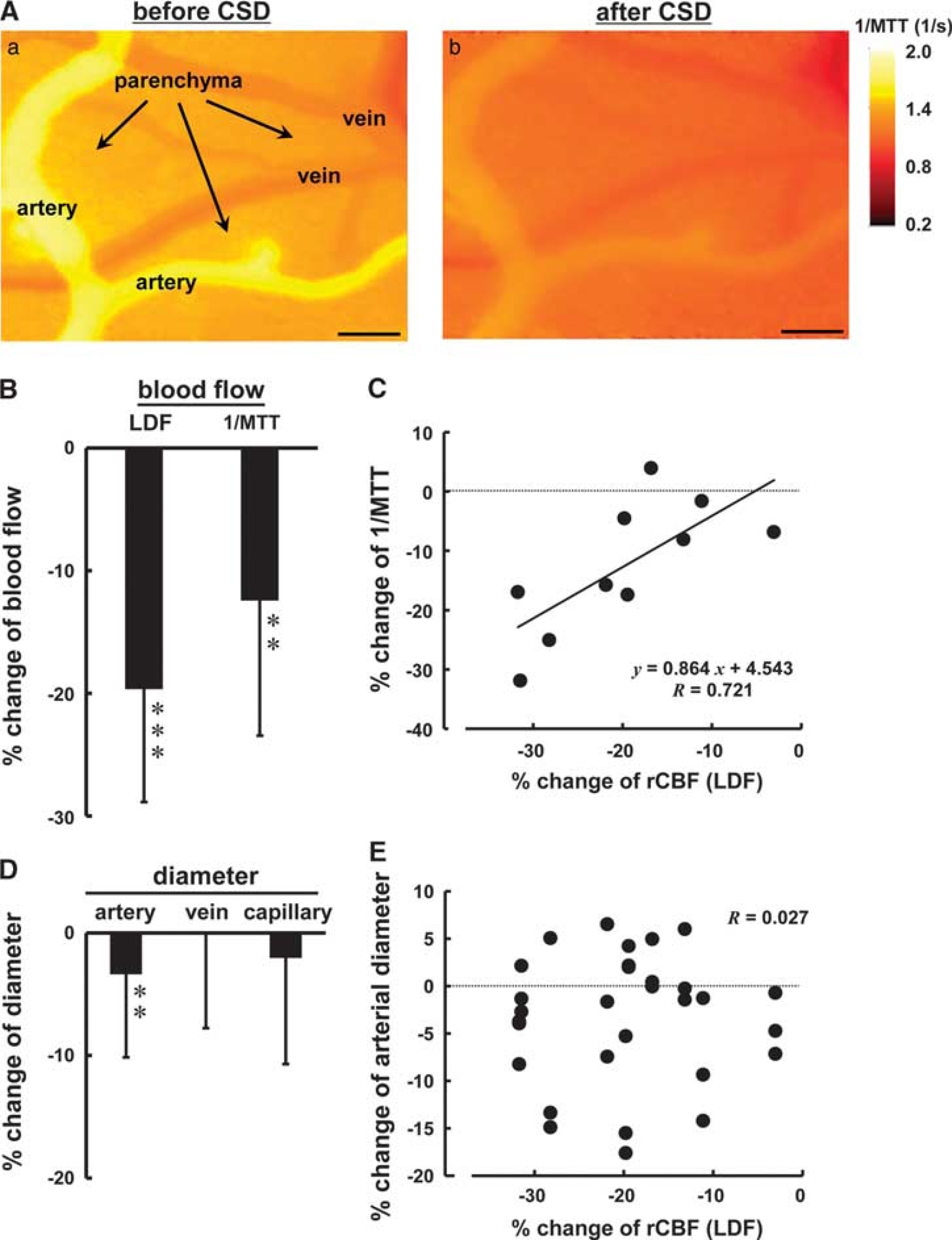
Reconstructed 2-D microflow map evaluated using the dye-dilution method, KEIO-IS1 (
Discussion
We examined diametric changes in various types of vessels during and after passage of CSD with a confocal laser-scanning fluorescence microscope and identified three vasomotor elements, initial constriction, subsequent dilation, and long-lasting vasoconstriction, which temporally corresponded to initial transient hypoperfusion, prominent hyperemia, and post-CSD oligemia, respectively. The response was diminished with each consecutive passage of CSD, even though the LDF-evaluated rCBF increase response was maintained. The diametric changes of pial veins and capillaries were much smaller than those of pial arteries. In the period of post-CSD oligemia, the decrease in cortical microflow measured using a dye-dilution method, KEIO-IS1, amounted to half of the response in LDF-evaluated rCBF, concomitantly with mild constriction of pial arteries.
The diametric changes in response to first CSD were very different from those in response to subsequent CSDs. It is well known that CSD may be induced by mechanical stimulation, and we positioned the DC potential electrode on the intact dura and the LDF probe on the skull to avoid mechanical induction of CSD. However, we cannot rule out the possibility of CSD occurrence in response to incision of the small window in the dura for KCl application. Indeed, a single pin prick induced CSD in 42.3% of trials. However, incision of the dura may be much milder than the 1-mm-deep pin prick, so we think the likelihood of accidental triggering of CSD is low. Furthermore, we confirmed that there was no occurrence of CSD for more than 30 minutes. Even if accidental CSD had been triggered, we have observed that rCBF recovered from oligemia within 30 to 60 minutes and the apparent transient dip of rCBF measured with LDF was elicited by subsequent CSD in a similar protocol (data not shown). Therefore, we considered that the CSD induced by KCl application was the first CSD in this experiment.
It is known that hypoxia and hypotension can alter the kinetics of the CSD-related CBF response. 27 Anesthesia affects MABP and CSD-induced CBF changes. 8 In this experiment, MABP and arterial O2 were rather low, and arterial CO2 was rather high, but we think the reason for this was that the mice were ventilated with room air under urethane anesthesia. We confirmed that autoregulation of CBF was maintained in these animals by the observation of stable CBF independent of ABP fluctuation before application of KCl. Therefore, we considered that the condition of the animals remained within the physiologic range.
We observed transient but pronounced vasoconstriction and hypoperfusion especially during the first passage of CSD. The period between the peak of arterial constriction and the transient dip of rCBF was approximately 9 seconds on average. The analyzed ROI was a 500-μm square in the clearly visible part of the cranial window of approximately 2 mm in diameter. Differences in the distance from the analyzed ROI and the LDF probe to the focus of CSD generation might account at least in part for the time lag between the rCBF dip and initial transient vasoconstriction. Therefore, it is reasonable to consider that marked constriction of pial arteries might trigger the transient dip in rCBF response.
The initial vasoconstriction might be a direct effect of extracellular potassium on vascular smooth muscle, at least in part, because elevated K+ is known to induce constriction of isolated basilar arteries. 11 Intraparenchymal microinjection of KCl also induced constriction of adjacent cortical arteries; however, CSD-induced vasoconstriction contains a thapsigargin-sensitive component, while KCl-induced vasoconstriction does not. 28 A contribution of an astrocytic Ca2+ wave and calcium-dependent phospholipase A2 to the initial vasoconstriction was suggested, so that the initial vasoconstriction in their case might not mediated by neuronal vesicular neurotransmitter release or activities of neurons innervating the vasculature. 28 Perivascular astrocytes may have played a key role in such a rapid constriction, probably via a cation transport mechanism, rather than release of vasoactive substances or metabolites.
On the onset of DC potential deflection, increase of rCBF and slight dilation of pial arteries often preceded the deep trough of rCBF and marked vasoconstriction. Cortical spreading depression caused an immediate rise of extracellular K+ level to more than 50 mmol/L.29, 30 Moderate elevation (<20 mmol/L) of extracellular K+ causes vasodilation via K+ channels, 31 whereas higher concentrations lead to vasoconstriction. 32 We observed a rather similar diametric response to pin prick, indicating that the response was not due to diffusion of potassium applied on the brain surface. Therefore, a moderate increase of extracellular K+ level might initially induce vasodilation, while further increase could lead to vasoconstriction.
In addition to our findings, initial transient hypoperfusion and marked constriction of pial arteries have been widely reported,11, 17, 19, 33 although there is also a report that initial hypoperfusion was much smaller or absent. 8 In mice, the CSD-related initial hypoperfusion is pronounced, and the hyperemic response is markedly diminished; indeed, the hyperemic peak barely reaches baseline.11, 24 Pronounced hypoperfusion induced by CSD in mice as compared with rats may be due to greater sensitivity of mouse cerebral vessels to elevated extracellular K+ concentration. 11 Subsequent dilation was attenuated in the presence of nitric oxide synthase inhibitor. 11 Production and/or degradation of nitric oxide might be different between species. In contrast, the rCBF response with a small dip appears similar to the CBF response in rats,7, 11, 20 in spite of the marked vasoconstriction in this experiment. We found profound hypoperfusion and severe oligemia with a hyperperfusion phase under baseline, as measured with LDF, in urethane-anesthetized C57BL/6J mice treated with a similar protocol (Supplementary Figure). Thus, strain differences may well exist.
Furthermore, vasoconstriction may be induced by arrival of a CSD wave traveling at 3 to 5 mm/min, so vasoconstriction might switch to dilation at the proximal end of the detection area when the CSD wave arrives at the distal end (Supplementary Video 3). Rapid or mild constriction could be detected as a transient dip of CBF in a smaller detection area by means of laser speckle flowmetry, though it was not detected with LDF in rats. 11 Calculations with our software KEIO-IS1 revealed initial oligemia with capillary flow stop followed by hyperemia and then oligemia, based on microscopical hemodilution curves in rats. 17 Thus, LDF might be less able to detect rapid changes in vessel diameter.
Increased activity of neurons and glial cells leads to production of metabolites and increased local blood flow in response to the metabolic demand (reviewed by Busija et al 10 ). Cortical spreading depression elevates cerebral metabolic rate and oxygen consumption,15, 34 resulting in a marked increase of CBF. 9 Furthermore, CSD induces severe hypoxia owing to the transient increase in O2 demand exceeding vascular O2 supply, which cannot be reversed with additional O2 inhalation. 8 We reported that CSD induced a decrease in PO2 in parenchyma approximately concurrently with DC potential deflection, followed by a rCBF increase in rats exposed to a similar protocol. 20 The rCBF increase appeared rather later than the DC potential deflection in this experiment. The CSD-induced hyperperfusion might be caused at least in part by compensation for local hypoxia. Thus, vasoconstriction seems to counteract vasodilation, resulting in the transient dip of rCBF. Furthermore, the marked constriction might reduce the hyperperfusion response to first CSD (Figure 4A (b)), compared with second or third CSD.
We found a weak but significant correlation of the CSD-induced diametric change of pial arteries with the basal diameter (Figure 5A). Approximately one-third of the arteries apparently did not dilate, as shown in Figure 5B, where the diameter did not return to the baseline level after marked constriction. Most pial arteries, however, showed an apparent vasodilation phase followed by a mild constriction phase corresponding to post-CSD oligemia, as shown in Figure 3A (c). Thus, vasoreactivity including constriction and dilation in response to CSD might be more potent in small arteries than in large arteries. This is consistent with findings in neonatal rats, 28 and also with a report that marked vasoconstriction was observed only in small arterioles (less than 40 μm) but not in large epicortical arteries in response to KCl microinjection in cats. 19 The diameter of pial arteries on the cortical surface of mouse is less than 50 μm. Therefore, it is reasonable that all arteries constricted in our experiment.
As compared with arteries, diametric changes of veins and capillaries were much smaller and prolonged, and diminished with repeated CSD passage. No obvious constriction was induced in veins, probably due to sparse vascular smooth muscle. We observed slight constriction and subsequent dilation of capillaries, but the changes were so heterogeneous. Because capillaries are located in parenchyma that lie approximately 50 μm below the brain surface, it was difficult to observe the entire length of capillaries in the same ROI. When the diameter at multiple points of a single capillary was measured, the change of diameter appeared to fluctuate randomly in 7 of 15 capillaries (data not shown). Thus, diametric changes of veins and capillaries do not seem to be necessary for functional hyperemia.
However, capillary flow is important to maintain neurovascular coupling. We observed slight constriction of capillaries in this experiment and a decrease of RBC velocity in intraparenchymal capillaries in adult rats, 20 in accordance with findings in neonatal rats. 28 Capillary bed constriction may also occur via swelling or morphologic change of glial cells35, 36 or neurons. 8 It has been suggested that contractility of pericytes surrounding capillaries has a role in determining capillary flow. 37 However, the contribution of pericytes to capillary flow control seems to be negligible during CSD-induced hyperemia, because passive dilation of capillaries, which occurred irrespective of the presence of pericytes, has been reported. 38 Thus, vasoreactivity of capillaries may not contribute to functional hyperemia, but may influence RBC flow in capillary and thus the function of neurovascular coupling.
No transient dip of rCBF in response to the second or later CSD could be observed in our experiment, whereas the peaks of the hyperperfusion phase were comparable. Reduced or absent arteriolar diametric responses to frequent repetitive CSD evoked by continuous stimulation have been reported in rats. 12 Yuzawa et al 24 indicated that the hemodynamic and metabolic responses to a second CSD could be superimposed on the oligemic phase, reduced CBF and suppression of metabolism after the first CSD in cortex of naive mice. They suggested that the first CSD might induce long-lasting severe hypoxia in the parenchyma and suppress the vasoconstrictive response to subsequent CSD. Indeed, we have reported suppression of RBC velocities in intraparenchymal capillaries during and after passage of CSD in rats. 20 Prolonged suppression of vasomotor activity and intraparenchymal metabolism might be due at least in part to the first CSD.
We also found a decrease in 1/MTT in the parenchyma concomitantly with slight constriction of pial arteries during post-CSD oligemia (Figure 7). It should be noted that rCBF evaluated with LDF and cortical microflow evaluated with KEIO-IS1 may not always correspond, due to practical and theoretical dissimilarities between the methods, namely the former reflects RBC velocity and the latter reflects plasma flow. However, a good correspondence of the changes elicited by hypercapnia-induced global vasodilation, as evaluated by the two methods, has been shown. 26 The high correlation obtained in this study supports the validity of comparing results obtained with the two methods, and the small decrease in 1/MTT suggests a contribution of intracortical blood flow rather than pial arteries. This is consistent with a previous report indicating that diametric changes of pial arteries may not always reflect rCBF responses evaluated with LDF. 39 It has been reported that CSD induced long-lasting perturbation of the cortical vasoreactivity to hypercapnia and basal forebrain electrical stimulation during oligemia in rats. 9 Thus, some derangement of diametric control of pial arteries might occur during post-CSD oligemia.
We found a rapid constriction of pial arteries counteracting the vasodilation response, which is seen as a small dip of rCBF, by simultaneous recording of rCBF and diameter. However, the reason why the hyperperfusion response is larger than the diametric changes in pial arteries is unclear. We consider that intraparenchymal vessels are important for the hyperperfusion response to CSD, since a constant increase in rCBF was elicited in spite of diminished vasomotor changes in the pial arteries.
Layer-specific vasodilation was showed, where electrical stimulation of the nucleus basalis of Meynert in mice induced vasodilation throughout the cortical layers, except for the cortical surface and the upper part of layer V of penetrating arteries. 40 Since such stimulation-induced hyperperfusion was measured with LDF, 41 vasodilation of intraparenchymal penetrating arteries rather than pial arteries lying at the cortical surface might contribute to the regional cortical blood flow response. Although diameter changes in pial arteries were diminished with each passage of CSD, the increase in LDF-evaluated rCBF was constant in our experiments. Thus, other vessels that we could not see in this study, such as penetrating arteries or intraparenchymal arteries, might dilate and contribute to the increase in rCBF, especially in the second and later CSD passages.
Peri-infarct depolarization, a similar phenomenon to CSD, occurs at a high frequency in the cerebral cortex in various pathophysiologic situations, such as stroke and subarachnoid hemorrhage. 3 Vasoconstriction accompanied with blood flow reduction might exacerbate the impairment of cortical blood vessel control, resulting in impaired brain function. Mild and long-lasting vasoconstriction with prolonged reduction of rCBF (oligemia) is associated with mitochondrial depolarization and activation of calcineurin, accompanied with impaired neurovascular coupling, 42 and is mediated by constrictor prostanoids and a metabolite of arachidonic acid, 20-hydroxyeicosatetraenoic acid. 43 Initial vasoconstriction with transient reduction of rCBF might result in a slow-down or stop of RBC flow in capillaries, vascular dysfunction, aggravation of parenchymal hypoxia, or greater damage to brain function. Further work to uncover the mechanism of acute vasoconstriction might lead to new therapeutic possibilities for protection against cerebral injury.
Conclusion
We observed a rapid and remarkable constriction, subsequent dilation and then long-lasting constriction of pial arteries in response to CSD. Each of these responses was diminished with successive passages of CSD, even though the rCBF increase response was well maintained. The CSD-induced rCBF increase was not always accompanied with vasodilation of pial arteries. These results indicate that intracortical vessels such as penetrating arteries are also involved.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.