
Abstract
Sex differences are well known in cerebral ischemia and may impact the effect of stroke treatments. In male rats, the MEK1/2 inhibitor U0126 reduces ischemia-induced endothelin type B (ETB) receptor upregulation, infarct size and improves acute neurologic function after experimental stroke. However, responses to this treatment in females and long-term effects on outcome are not known. Initial experiments used in vitro organ culture of cerebral arteries, confirming ERK1/2 activation and increased ETB receptor-mediated vasoconstriction in female cerebral arteries. Transient middle cerebral artery occlusion (tMCAO, 120 minutes) was induced in female Wistar rats, with U0126 (30 mg/kg intraperitoneally) or vehicle administered at 0 and 24 hours of reperfusion, or with no treatment. Infarct volumes were determined and neurologic function was assessed by 6-point and 28-point neuroscores. ETB receptor-mediated contraction was studied with myograph and protein expression with immunohistochemistry. In vitro organ culture and tMCAO resulted in vascular ETB receptor upregulation and activation of ERK1/2 that was prevented by U0126. Although no effect on infarct size, U0126 improved the long-term neurologic function after experimental stroke in female rats. In conclusion, early prevention of the ERK1/2 activation and ETB receptor-mediated vasoconstriction in the cerebral vasculature after ischemic stroke in female rats improves the long-term neurologic outcome.
Keywords
Introduction
Sex differences exist in occurrence, outcome, and treatment of cerebral ischemia.1, 2 Throughout much of the lifespan there is a higher incidence of stroke in men than in women. Sex differences also exist in the response to stroke therapy, showed by greater benefit and neurologic improvement after tissue plasminogen activator treatment in women compared with men.3, 4 Furthermore, sex-specific responses have been reported for aspirin therapy in stroke patients.5, 6
We have shown that vasoconstrictor endothelin type B (ETB) receptors are upregulated in smooth muscle cells of cerebral arteries after stroke in human 7 and rat.8, 9 Increased expression and contractile responses of ETB receptors are hypothesized to reduce cerebral blood flow further after an ischemic stroke and exacerbate tissue damage. 10 Upregulation of ETB receptors was observed in arteries placed in organ culture, and this has become an established in vitro method for studying underlying mechanisms of receptor upregulation. 11 Previous studies in male rats showed that upregulation of ETB and other vasoconstrictor receptors occurs via the MEK/ERK1/2 pathway in both stroke and organ culture models. 10 This finding led us to examine the MEK1/2 inhibitor U0126 which, not only attenuates vascular receptor upregulation, but also reduces infarct size and improves neurologic function when administered after experimental stroke.12, 13 The previous data suggest a promising therapeutic lead for stroke treatment, but it is not known whether similar pathways contribute to the effects of stroke in females. Therefore, we aim to investigate whether (1) vascular ETB receptor upregulation occurs in female cerebral arteries after transient middle cerebral artery occlusion (tMCAO) and organ culture, (2) the receptor upregulation is mediated via the MEK/ERK1/2 pathway, and (3) treatment with the MEK1/2 inhibitor U0126 suppresses ETB receptor upregulation and improves long-term neurologic outcome after ischemic stroke in female rats.
Materials and methods
All experiments and procedures were performed in full compliance with the guidelines set forth in the European Council's Convention for the Protection of Vertebrate Animals Used for Experimental and other Scientific Purposes, and were approved by the Malmö-Lund Institutional Ethics Committee under the Swedish National Department of Agriculture (M179-11, M188-12, M8-09). The study complies with the ARRIVE guidelines (Animal Research: Reporting In Vivo Experiments).
Organ Culture
Thirteen-week-old female Sprague Dawley rats (240 to 250 g) were anesthetized with CO2 and decapitated. Middle cerebral arteries (MCAs) were used directly as fresh, or placed in organ culture for 24 hours in Dulbecco's modified Eagle's medium (DMEM; Gibco, Invitrogen, Carlsbad, CA, USA) supplemented with penicillin (1,00,000 U/L), streptomycin (100 mg/L), and amphotericin B (0.25 mg/L) at 37°C in humidified 5% CO2 in air. Before incubation, the MEK1/2 inhibitor U0126 (LC Laboratories, Boston, MA, USA) 10 μmol/L, or equal volume of DMSO was added. For western blot, arteries from the Circle of Willis (tissue from two animals were pooled) were incubated as described above except for 6 hours of organ culture. In total n=39 animals were used for the in vitro studies (see figure legends for exact number of animals per group and experiment).
Middle Cerebral Artery Occlusion
Twelve-week-old female Wistar rats (250 to 265 g) were subjected to intraluminal MCA occlusion. 13 Before surgeries, the estrous cycle was monitored (daily collection of vaginal smears and characterization of the types of cells 14 present for three consecutive cycles, see Supplementary Information) and only female rats that were under low influence of estrogen, i.e. not in proestrus, at the day of surgery, were selected to minimize possible effects from hormonal fluctuation. Animals were anesthetized by 4.5% isoflurane in N2O:O2 (70:30) and kept anesthetized by 1% to 1.5% isoflurane in N2O/O2 through inhalation. Physiologic parameters were measured immediately before the occlusion (Supplementary Tables S1 and S2 in the Supplementary Information). Animals were occluded for 2 hours followed by reperfusion, and killed 2 or 14 days later. Average Laser-Doppler flow reductions of cerebral blood flow were as follows: 70±3% in untreated, 77±3% in vehicle, and 76±2% in U0126 animals (lower inclusion limit 58%). One of the U0126-treated rats died between 24 and 48 hours after tMCAO. N=32 animals were included in the in vivo experiments (see figure legends for exact number of animals per group and experiment).
U0126 Treatment
Study I (Figure 1) contains three experimental groups: untreated, U0126, and vehicle (DMSO), and rats were terminated at 48 hours of reperfusion. Animals treated with vehicle confirmed earlier studies showing that DMSO alone has no effect on the parameters studied. The MEK1/2 inhibitor U0126 (30 mg/kg intraperitoneally) was injected at 0 and 24 hours of reperfusion after tMCAO based on the previous evaluation of the drug in male rats. 13 Animals in study II (Figure 1) were administered U0126 or vehicle as described above and were killed 14 days after tMCAO. Experimental group assignments were randomized and blinded to the surgical experimenter.
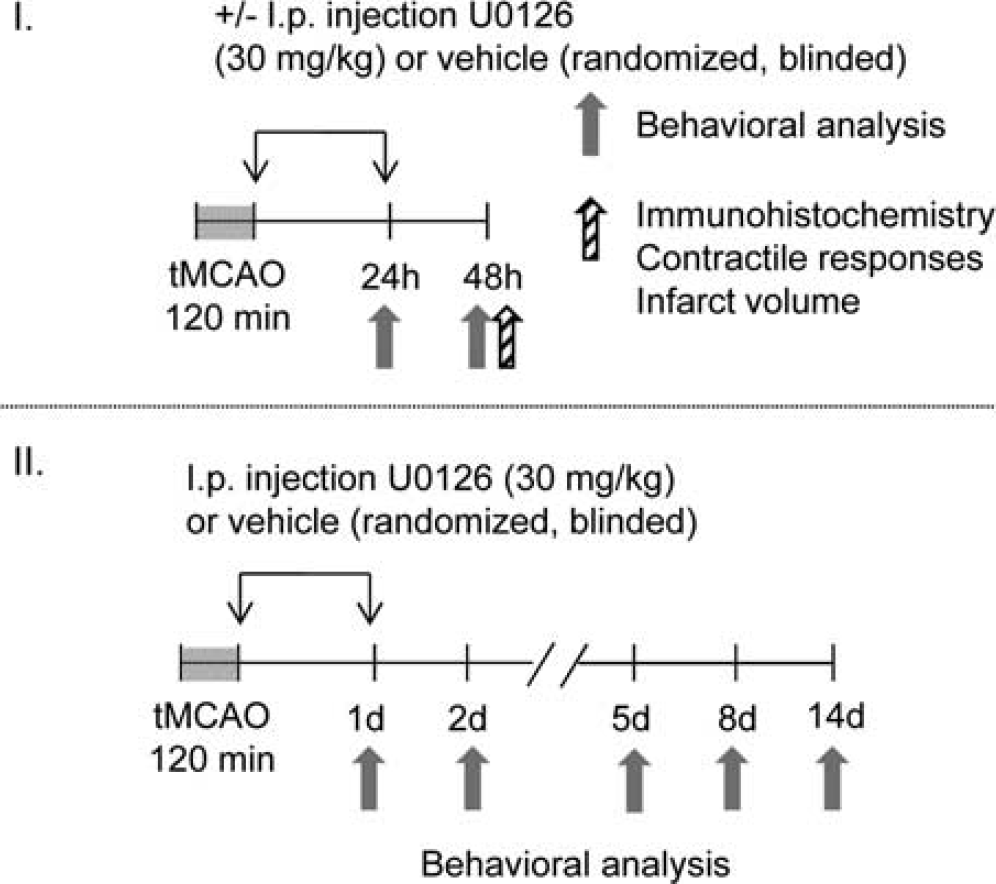
Experimental design. Animals were terminated 2 days (study I) and 14 days (study II) after transient middle cerebral artery occlusion (tMCAO).
Neurologic Examination
Neurologic examination was performed at 1, 2, 5, 8, and 14 days after tMCAO according to established 6-point15, 16 and 28-point 17 scoring systems by a blinded and experienced rater. Scoring was determined on a scale starting from 0 for severe impairment to the maximum score point for healthy function in the 28-point neuroscore. The 6-point neuroscore is graded in six levels from 0—no visible defects to 5—death.
Infarct Size Measurement
Brains were fixed in 4% paraformaldehyde and sectioned into 40 μm coronal sections using a microtome (Leica Biosystems, Nussloch, Germany), collected at 1 mm intervals and immunostained with neuron-specific nuclear protein NeuN (Millipore, Billerica, MA, USA, diluted to 1:1,000). Images were analyzed in a blinded manner using the ImageJ software (http://rsb.info.nih.gov/ij/) and infarct volumes were calculated by an indirect method of subtracting the nonlesioned volume of the ipsilateral hemisphere from the nonlesioned volume of the contralateral hemisphere. 18 The resulting values were analyzed with Grubb's test and one outlier was removed.
In vitro Pharmacology
Contractile responses were examined in a Mulvany-Halpern wire myograph (Danish Myo Technology A/S, Aarhus, Denmark) as described previously.19, 20 The contraction induced by K+ (63.5 mmol/L) was used as a reference for contractile capacity and set to 100%. Endothelial influences were blocked by L-NG-Nitroarginine methyl ester (L-NAME, 100 μmol/L) and indomethacin (10 μmol/L). Concentration—response curves were obtained by cumulative application of sarafotoxin 6c (S6c, ETB receptor agonist, 10−11 to 10−7 mol/L) and ET-1 (ETA and ETB receptor agonist, 10−11 to 10−7 mol/L). Before application of ET-1, ETB receptors were inhibited by BQ788 (1 μmol/L), or by desensitization with S6c. The maximum contraction elicited by an agonist is represented by EMAX, and pEC50 represents the negative logarithm of the agonist concentration that produced half the maximum response.
If not stated otherwise, the substances were purchased from Sigma-Aldrich (St Louis, MO, USA). ET-1 and S6c (Alexis Biochemicals, Farmingdale, NY, USA) were dissolved in 5% acetic acid and 0.1% bovine serum albumin in distilled water. L-NAME hydrochloride, indomethacin, and BQ788 were dissolved in bicarbonate buffer solution, ethanol, and DMSO, respectively.
Immunohistochemistry
Isolated MCAs were fixed in 4% paraformaldehyde followed by rinsing in sucrose containing phosphate buffers. Cryosections (10 μm) were permeabilized in PBST and then exposed overnight to primary antibody: sheep anti-rat ETB 1:100 (#ALX-210-506A, Enzo, Lausen, Switzerland) or mouse anti-rat dually phosphorylated ERK1/2 1:200 (#ab50011, Abcam, Cambridge, UK). Secondary antibodies Dylight 594 donkey anti-sheep (1:50, Jackson ImmunoResearch, West Grove, PA, USA) or FITC-conjugated donkey anti-mouse (1:100, Jackson ImmunoResearch) were applied for 1 hour. Sections were mounted in anti-fading medium containing 4′,6-diamidino-2-phenylindole (DAPI) that stains nuclei (Vectashield, Vector Laboratories Inc., Burlingame, CA, USA). The experiments were repeated to ensure reproducibility. Omission of the primary antibody served as a negative control. Immunoreactivity was visualized at the appropriate wavelengths with an epifluorescence microscope (Nikon 80i; Tokyo, Japan) and photographed with an attached Nikon DS-2Mv camera.
Qualitative assessment of immunohistochemical staining was evaluated by two different human readers, blinded to the study details and with extensive expertise in the field. Pairwise comparisons of smooth muscle cell layer immunoreactivity in the occluded MCA and non-occluded MCA were performed. In addition, the expression was evaluated by blinded measurement of the fluorescence intensity in the smooth muscle layer using the ImageJ software. The mean intensity in the occluded MCA was normalized to the non-occluded MCA within the same animal. One outlier was removed after analysis with Grubb's test. For comparative studies, immunoreactivity was visualized with the same microscope settings during the same day. Quantification of the immunofluorescence intensity was not performed in the vehicle group due to low animal numbers and only qualitative assessment of the immunohistochemical staining was performed for this group, see description above.
Western Blot
Cerebral arteries (basilar, middle, anterior, and posterior) cultured for 6 hours in the presence of DMSO or U0126 (10 μmol/L) were snap-frozen in liquid nitrogen followed by protein extraction. The samples were boiled for 5 minutes in Laemmli sample buffer and 2-mercaptoethanol (Bio-Rad, Hercules, CA, USA), loaded on a 4% to 15% linear gradient Tris—HCl gel and separated by SDS-PAGE (5 μg/lane). Proteins were transferred onto a nitrocellulose membrane (Bio-Rad) and incubated overnight in primary antibodies against active dually phosphorylated ERK1/2 (1:1,000; #9101, Cell Signaling Technology, Beverly, MA, USA) and smooth muscle specific protein SM22-α (1:1,000; #ab10135, Abcam) followed by secondary horseradish peroxidase-linked anti-rabbit (1:2,000; Cell Signaling) or anti-goat (1:10,000; Pierce Biotechnology, Rockford, IL, USA) antibodies for 1 hour. Protein bands were visualized by exposure to a charged coupled device camera (LAS1000; Fujifilm, Stamford, CT, USA) using a chemiluminescence kit (Pierce Biotechnology). Membranes were stripped and reprobed for total levels of ERK1/2 (1:1,000; #9107, Cell Signaling). Active dually phosphorylated ERK1/2 expressions levels were calculated as the ratio of total levels of ERK1/2 normalized to SM22-α expression.
Statistics
Data are expressed as mean±standard error of the mean (s.e.m.), and n refers to the number of rats. Statistical analyses were performed with nonparametric Mann—Whitney's test or Kruskal—Wallis test followed by Dunn's multiple comparison test. P<0.05 was considered as significant.
Results
Endothelin Type B Receptor-Mediated Vasoconstriction and ERK1/2 Activation in Female Cerebral Arteries after In vitro Organ Culture
In initial experiments, in vitro organ culture of isolated cerebral arteries was used to investigate whether increased ETB receptor-mediated vasoconstriction and ERK1/2 activation, similar to the observations in males, occur in females. Smooth muscle cell-specific contractile responses were examined by application of the selective ETB receptor agonist S6c in the presence of L-NAME and indomethacin to block endothelial-mediated responses by nitric oxide and prostaglandins. S6c elicited no contractile responses in freshly obtained MCAs while a strong vasoconstriction was seen after 24 hours of organ culture (Figure 2A). There were no differences in the responses of MCAs taken from animals killed at the time of estrus or diestrus (data not shown); these data were pooled. ET-1-induced vasoconstriction (measured after desensitization by S6c) was enhanced after organ culture as compared with fresh arteries (Supplementary Table S3). However, the contractile capacity (K+-induced responses) was higher in cultured arteries than in nonincubated fresh arteries (6.4±0.5 vs 3.4±0.5 mN).
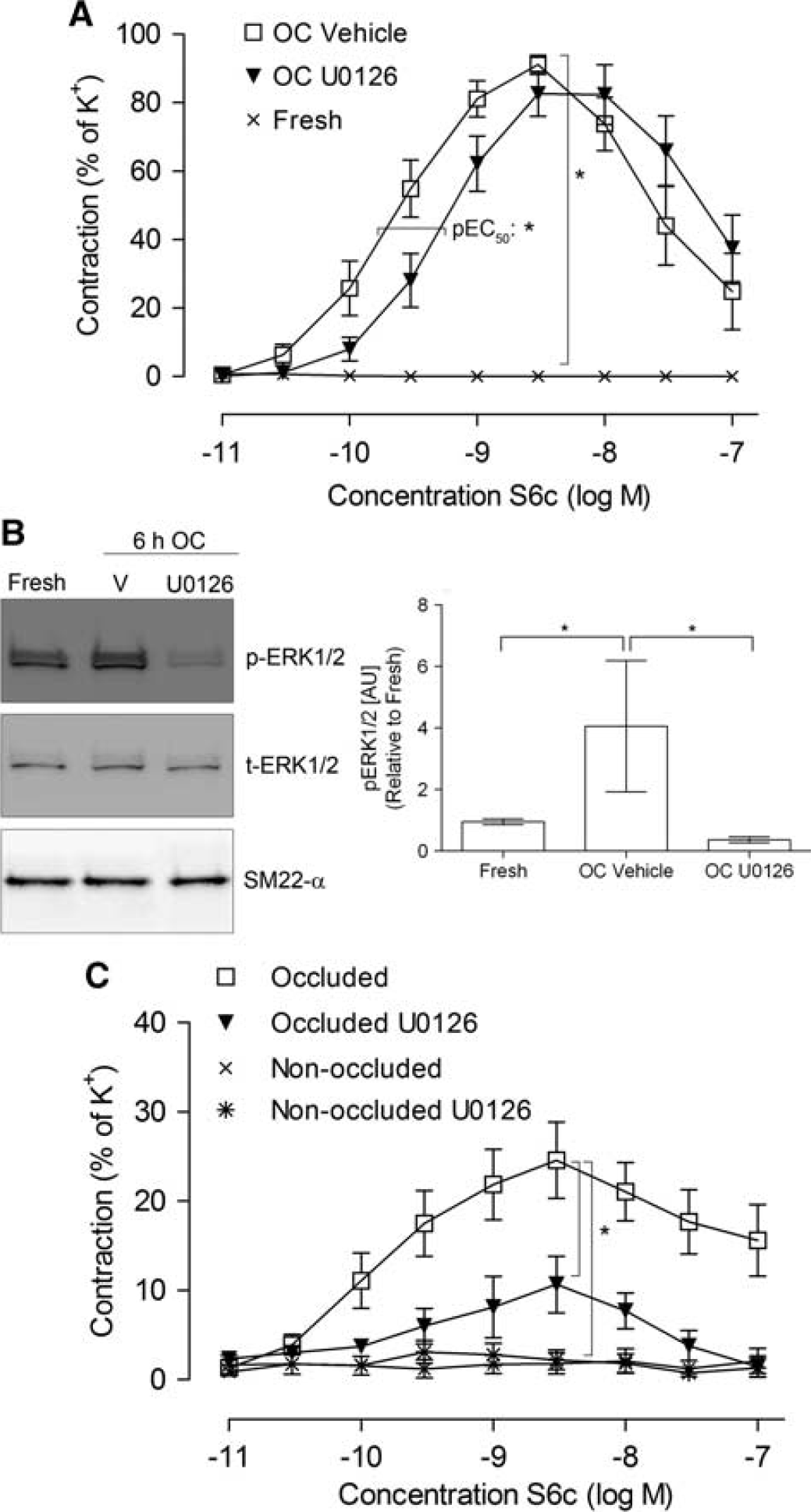
Increased endothelin type B (ETB) receptor-mediated vasoconstriction and p-ERK1/2 expression in female rat MCAs. (
Arteries incubated in the presence of U0126 (10 μmol/L) resulted in a right-ward shift of the concentration—response curve (change in pEC50), suggesting involvement of the MEK/ERK1/2 pathway in ETB receptor upregulation after organ culture (Figure 2A). Activation of ERK1/2 was confirmed at 6 hours of organ culture by western blot (Figure 2B), and U0126 prevented this process. Next, we investigated the role of ERK1/2 in ETB receptor upregulation and outcome after experimental stroke.
ERK1/2-Mediated Increase in Endothelin Type B Receptor Vasoconstriction In Female Rats after Transient Middle Cerebral Artery Occlusion
Rats were subjected to 120 minutes tMCAO and thereafter treated with the MEK1/2 inhibitor U0126 (30 mg/kg intraperitoneally) at 0 and 24 hours of reperfusion. In concert to the in vitro results from nonincubated arteries, left-side non-occluded MCAs elicited weak contractile responses to S6c (Figure 2C). Occlusion of the right MCA significantly enhanced the ETB receptor-mediated contraction (Figure 2C). After treatment with U0126, the vasoconstriction to S6c was markedly reduced. ET-1-induced vasoconstriction was not significantly different in non-occluded and occluded MCAs, and not affected by U0126 (Supplementary Table S4).
Increased Endothelin Type B Receptor and p-ERK1/2 Expression after Transient Middle Cerebral Artery Occlusion Is Prevented by U0126
Expression of ETB receptor was examined in the smooth muscle cell layer of occluded and non-occluded MCAs with immunohistochemistry. We observed a marked increase in ETB receptor expression in occluded MCAs compared with non-occluded MCAs of untreated and vehicle-treated animals, while animals treated with U0126 did not show any increase in arterial ETB receptor expression (Figure 3A). This observation was substantiated by blinded measurements of fluorescence intensity in the smooth muscle cell layer demonstrating a significant increase in ETB receptor immunoreactivity in the occluded MCA (Figure 3C). This increase in ETB receptor expression was prevented by treatment with U0126. As shown in Figures 3B and 3D occlusion of the MCA resulted in increased p-ERK1/2 immunoreactivity in the smooth muscle cell layer compared with non-occluded MCAs in untreated and vehicle-treated animals that was abolished after treatment with the MEK1/2 inhibitor.
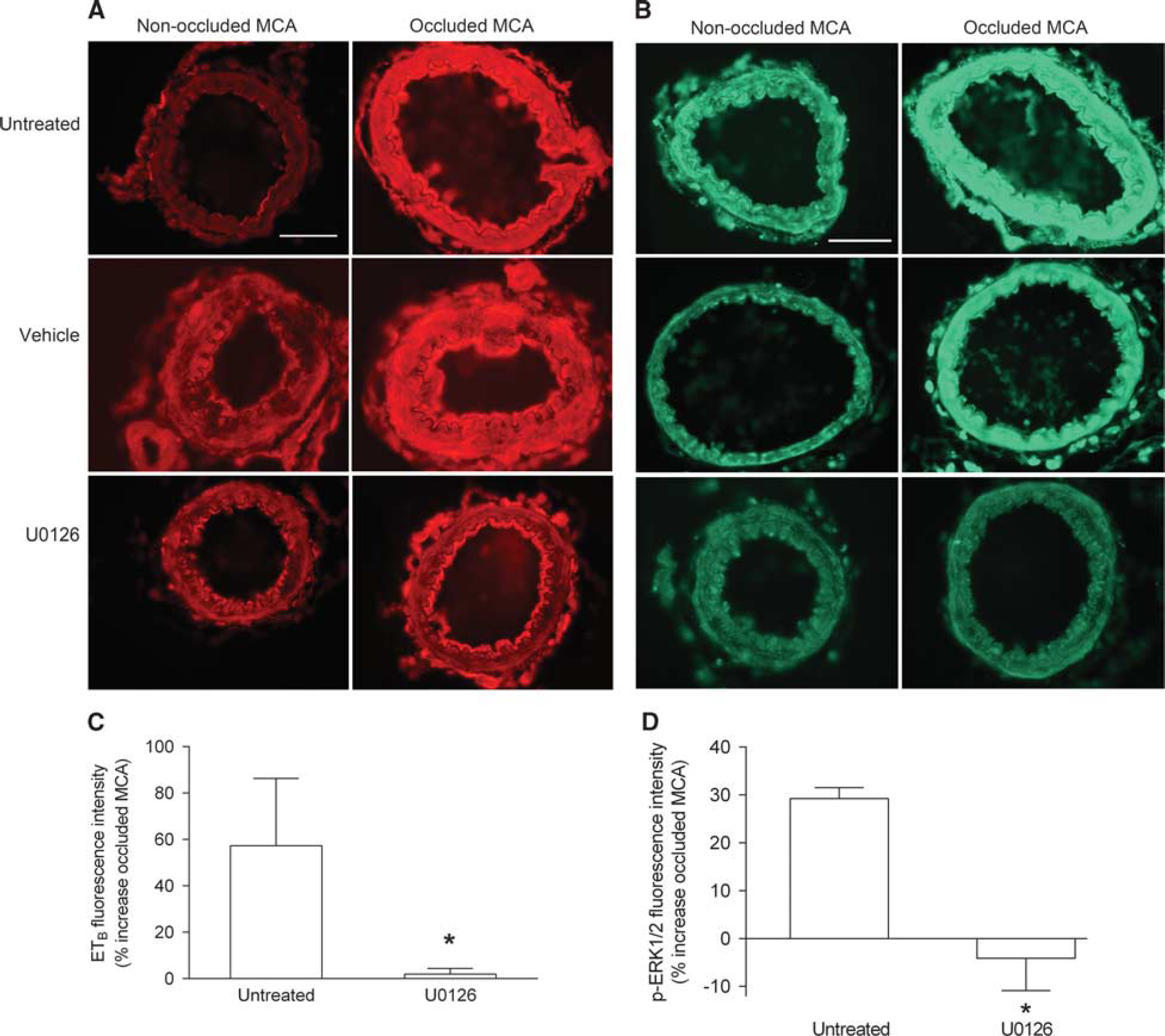
Ischemia-induced increase in smooth muscle endothelin type B (ETB) receptor and p-ERK1/2 expression after transient middle cerebral artery occlusion (tMCAO) is prevented by U0126 treatment. (
U0126 Improves Neurologic Function 14 Days after Transient Middle Cerebral Artery Occlusion Without Affecting the Infarct Volume
Brain infarct volume 2 days after tMCAO was assessed using NeuN staining to identify areas of healthy tissue. The infarct volumes were not significantly different in the experimental groups (Figures 4A and 4B).
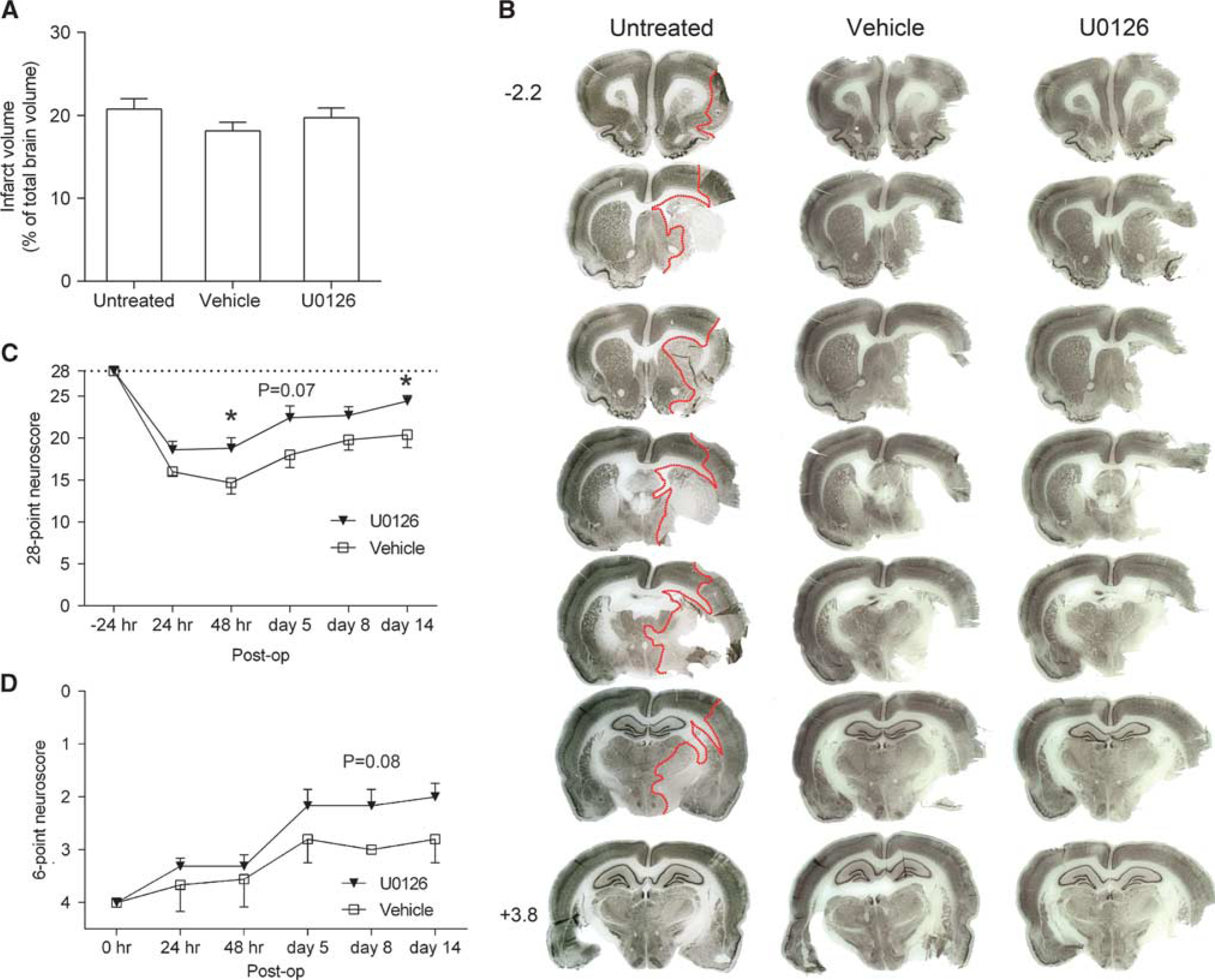
MEK1/2 inhibition results in improved neurologic function after transient middle cerebral artery occlusion (tMCAO). (
A long-term study was performed to investigate effects of U0126 treatment on the neurologic function up to 14 days after tMCAO. Neurologic function was assessed at 1, 2, 5, 8, and 14 days after tMCAO by 28-point and 6-point neuroscores (Figures 4C and 4D). Despite similar infarct volumes, there were significant improvements in the neurologic function at 2 and 14 days with the 28-point neuroscore in animals treated with U0126 compared with animals treated with vehicle (Figure 4C). Gross evaluation of the neurologic function with 6-point neuroscore did not reach statistical significance (Figure 4D).
Discussion
The main findings of this work are (1) vasoconstrictor ETB receptors located in the smooth muscle cells are upregulated in female rat MCAs subjected to in vivo tMCAO and in vitro organ culture; (2) ETB receptor upregulation in females is mediated via the MEK/ERK1/2 pathway (elevated p-ERK1/2), and (3) the MEK1/2 inhibitor U0126 given after tMCAO attenuates ischemia-induced ETB receptor upregulation and improves the long-term sensorimotor function. These findings indicate that vasoconstrictor ETB receptor upregulation, suggested to reduce cerebral blood flow and aggravate tissue damage after stroke, occurs in female rats after focal cerebral ischemia, similar to the previous findings in males.9, 13 Our results suggest that the MEK1/2 inhibitor U0126 could be a potential treatment for stroke in both sexes.
Endothelin Type B Receptor-Mediated Vasoconstriction after Transient Middle Cerebral Artery Occlusion and Organ Culture
There are two subtypes of ET receptors; ETA receptor activation results in vascular smooth muscle cell contraction and cell proliferation, 21 while ETB receptors normally are expressed on endothelial cells where they induce vasodilatation and mediate clearance of ET-1. However, ETB receptors have been shown to induce vasoconstriction in rat mesenteric arteries 22 and pulmonary resistance arteries from human and rat. 23 We have previously shown increased vasoconstriction mediated by ETB receptors in the cerebral circulation after experimental stroke.8, 9 Increased ETA and ETB receptor mRNA and protein were found in the smooth muscle cell layer of MCAs subjected to tMCAO.9, 13 To this date, studies on ET receptor plasticity in stroke, reviewed by, 10 have been conducted only on male rats.
A more complete understanding of the mechanisms underlying sex differences in stroke could result in more appropriate treatment strategies for patients of both sexes. To unravel ET receptor-mediated vasoactive responses in females, MCAs were subjected to tMCAO and organ culture followed by contractile function studies with myograph. Significantly increased ETB receptor-mediated vasoconstriction was observed in right MCAs subjected to tMCAO compared with the non-occluded left MCAs where only weak contractile responses via ETB receptors were detected (EMAX: 25% and 3%, respectively). Similar magnitude in EMAX for ETB vasoconstriction has been reported in male MCAs subjected to tMCAO followed by 24 hours of reperfusion [30%; 11 40%; 12 21% 24 ) and to a larger extent after 48 hours of reperfusion (EMAX: 68% 9 ). Sex differences in ETB receptor-mediated contraction have been observed in other disease models; male aortas from hypertensive rats was reported to show a higher increase in ETB receptor contraction than females. 25 Additionally, we recently discovered that organ cultured human cerebral arteries from women are less responsive to ET-1 and angiotensin II than males. 19 There may be a sex difference in the ETB receptor-mediated vasoconstriction after tMCAO; however, additional studies with direct comparison of males and females would be needed to address this point. The present study explores ET receptor-mediated responses in female MCAs subjected to experimental stroke and organ culture. We found increased ETB receptor-mediated vasoconstriction after in vivo experimental stroke and in vitro organ culture, demonstrating that ETB receptor upregulation previously reported for males, also occurs in females.
Endothelin Type B Receptor Upregulation Is Mediated by MEK/ERK1/2 Signaling Pathways
Next, we focused on underlying signaling pathways in ETB receptor upregulation after tMCAO. ETB receptor upregulation has been associated with increased activation of ERK1/2, 13 which was also seen in the present study as increased p-ERK1/2 expression in arteries after tMCAO and 6 hours of organ culture assessed with immunohistochemistry and western blot, respectively. Increased p-ERK1/2 has been shown in numerous studies on focal and global ischemia; however, whether its actions are protective or detrimental are under debate. 26 In the present study, MEK1/2 inhibition by treatment with U0126 at 0 hour and 24 hours after tMCAO attenuated the ischemia-induced increase in ETB receptor-mediated vasoconstriction (EMAX: 25% vs 12% in U0126). Additionally, U0126 prevented the increase in vascular smooth muscle cell ETB receptor expression.
Improved Long-Term Neurologic Function by U0126
The infarct volume at 48 hours after tMCAO in the current study was not affected by treatment with U0126. In contrast, our previous results in males show significantly reduced infarct volume by U0126.12, 13 Female rats experience less ischemic damage than males when subjected to similar type of stroke, 27 which the present data suggest. Estrogen has been suggested to account for these differences by neuroprotective mechanisms, and although the females in the present study were on low estrogen at the day of surgery, previous hormonal exposure likely has caused irreversible changes.
Importantly, treatment with U0126 improved the long-term sensorimotor function up to 14 days after tMCAO. Similarly to the present findings, improved motor control without affecting the infarct size was recently reported for memantine treatment, an N-methyl-D-aspartate glutamate receptor antagonist given 2 hours after the stroke. 28 In the study by Lopez-Valdez et al, 28 the authors suggest that neuroprotection did not have a significant role in the improved neurologic outcome. The mechanisms by which U0126 improves long-term neurologic function without affecting the infarct volume in the present study are not known, but we speculate that U0126 exerts beneficial effects on the cerebrovasculature that are not large enough to determine infarct volume or to be detected by NeuN staining, but significant enough to contribute to improved long-term recovery after stroke. Perhaps in females, where cerebral arteries are under larger influence of endothelial vasodilatory mechanisms,29, 30, 31 increased ETB receptor expression and cerebral vasoconstriction after stroke might not have the same detrimental effect as in males, and thereby determine infarct volume to a lesser extent. We suggest that early activation of ERK1/2 is detrimental and by preventing its action to reduce ETB receptor upregulation and vasoconstriction in the acute phase, in return improved long-term poststroke neurologic function is achieved.
Conclusion
The present study shows for the first time in females that ETB receptor-mediated vasoconstriction in cerebral arteries increases after focal cerebral ischemia and organ culture. Inhibition of the MEK/ERK1/2 pathway attenuates this response and significantly improves the long-term sensorimotor function. MEK1/2 inhibition is therefore a promising therapeutic strategy for stroke in both males and females.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.